Энолаза
| фосфопируватгидратаза | |||
|---|---|---|---|
 Димер дрожжевой енолазы. [1] | |||
| Идентификаторы | |||
| Номер ЕС. | 4.2.1.11 | ||
| Номер CAS. | 9014-08-8 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| Энолаза, N-концевой домен | |||
|---|---|---|---|
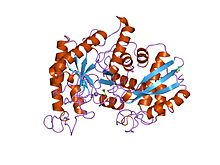 Рентгеновская структура и каталитический механизм енолазы омаров | |||
| Идентификаторы | |||
| Символ | Энолаза_N | ||
| Пфам | PF03952 | ||
| Пфам Клан | CL0227 | ||
| ИнтерПро | ИПР020811 | ||
| PROSITE | PDOC00148 | ||
| СКОП2 | 1элс / СКОПе / СУПФАМ | ||
| |||
| Энолаза | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | Энолаза | ||
| Пфам | PF00113 | ||
| ИнтерПро | IPR000941 | ||
| PROSITE | PDOC00148 | ||
| |||
Фосфопируватгидратаза , обычно известная как енолаза , представляет собой металлофермент ( EC 4.2.1.11 ), который катализирует превращение 2-фосфоглицерата (2-PG) в фосфоенолпируват (PEP), девятую и предпоследнюю стадию гликолиза . реакция Химическая :
- 2-фосфо- D -глицерат фосфоенолпируват + H 2 O

Фосфопируватгидратаза принадлежит к семейству лиаз , в частности гидролиаз, которые расщепляют углерод-кислородные связи. Систематическое название этого фермента — 2-фосфо- D -глицератгидролиаза (фосфоенолпируватобразующая) .
Реакция обратима и зависит от концентрации субстратов в окружающей среде. [3] Оптимальный pH для человеческого фермента составляет 6,5. [4] Энолаза присутствует во всех тканях и организмах, способных к гликолизу или ферментации . Фермент был открыт Ломанном и Мейергофом в 1934 году. [5] и с тех пор был выделен из различных источников, включая человеческие мышцы и эритроциты . [4] У людей дефицит ENO1 связан с наследственной гемолитической анемией , а дефицит ENO3 связан с болезнью накопления гликогена XIII типа .
Изоферменты [ править ]
У человека есть три субъединицы енолазы: α , β и γ , каждая из которых кодируется отдельным геном, который может объединяться с образованием пяти различных изоферментов : αα, αβ, αγ, ββ и γγ. [3] [6] Три из этих изоферментов (все гомодимеры) чаще встречаются в клетках взрослого человека, чем остальные:
- αα или ненейрональная енолаза (NNE). Также известен как енолаза 1 . Обнаружен в различных тканях, включая печень, мозг, почки, селезенку, жировую ткань. Он присутствует на определенном уровне во всех нормальных клетках человека.
- ββ или мышечная енолаза (MSE). Также известен как енолаза 3 . Этот фермент в основном ограничен мышцами, где он присутствует в мышцах в очень высоких количествах.
- γγ или нейрон-специфическая енолаза (NSE). Также известен как енолаза 2 . Экспрессируется на очень высоких уровнях в нейронах и нервных тканях, где может составлять до 3% от общего количества растворимого белка. В большинстве клеток млекопитающих он экспрессируется на гораздо более низких уровнях.
Разные изоферменты, присутствуя в одной и той же клетке, легко образуют гетеродимеры . [ нужна ссылка ]
Структура [ править ]
Энолаза является членом большого суперсемейства енолаз . Его молекулярная масса составляет 82 000–100 000 дальтон в зависимости от изоформы. [3] [4] В альфа-енолазе человека две субъединицы ориентированы антипараллельно , так что Glu 20 одна субъединица образует ионную связь с Arg 414 другой субъединицы. [3] Каждая субъединица имеет два отдельных домена. Меньший N-концевой домен состоит из трех α-спиралей и четырех β-листов . [3] [6] Более крупный С-концевой домен начинается с двух β-листов, за которыми следуют две α-спирали, и заканчивается бочонком, состоящим из чередующихся β-листов и α-спиралей, расположенных так, что β-бета-листы окружены α-спиралями. [3] [6] Компактная глобулярная структура фермента является результатом значительных гидрофобных взаимодействий между этими двумя доменами.
Энолаза — высококонсервативный фермент, активность которого особенно важна для пяти остатков в активном центре. По сравнению с енолазой дикого типа, мутантная енолаза, отличающаяся либо по Glu 168 , Глу 211 , Свет 345 , или Лис 396 остаток имеет уровень активности, сниженный в 105 раз. [3] Также изменения, затрагивающие Его 159 оставить мутанту только 0,01% его каталитической активности. [3] Неотъемлемой частью енолазы являются два Mg 2+ кофакторы в активном центре, которые служат для стабилизации отрицательных зарядов в субстрате. [3] [6]
В последнее время подработка функций нескольких енолаз, таких как взаимодействие с плазминогеном , привлекла интерес к каталитическим петлям ферментов и их структурному разнообразию. [7] [8]
 Трехмерное изображение димера енолазы в антипараллельной ориентации. N-концевая Glu одного димера 20 образует ионную связь с C-концевым Arg другого 414 для стабилизации четвертичной структуры фермента.
Трехмерное изображение димера енолазы в антипараллельной ориентации. N-концевая Glu одного димера 20 образует ионную связь с C-концевым Arg другого 414 для стабилизации четвертичной структуры фермента. Активный сайт енолазы в середине ствола С-концевого домена. На фото два Mg 2+ кофакторы и пять высококонсервативных остатков, необходимых для правильной каталитической функции: Его 159 , Глу 168 , Глу 211 , Свет 345 , Свет 396 .
Активный сайт енолазы в середине ствола С-концевого домена. На фото два Mg 2+ кофакторы и пять высококонсервативных остатков, необходимых для правильной каталитической функции: Его 159 , Глу 168 , Глу 211 , Свет 345 , Свет 396 .


Механизм [ править ]

Предполагается, что с использованием изотопных зондов общий механизм превращения 2-PG в PEP представляет собой реакцию элиминирования E1cB с участием промежуточного карбаниона. [9] Следующий подробный механизм основан на исследованиях кристаллической структуры и кинетики . [3] [10] [11] [12] [13] [14] [15] Когда субстрат, 2-фосфоглицерат, связывается с α-енолазой, его карбоксильная группа координируется с двумя кофакторами ионов магния в активном центре. Это стабилизирует отрицательный заряд депротонированного кислорода, одновременно увеличивая кислотность альфа-водорода. Лиз Энолазы 345 депротонирует альфа-водород, и полученный отрицательный заряд стабилизируется за счет резонанса с карбоксилатным кислородом и кофакторами ионов магния. После образования промежуточного карбаниона гидроксид C3 удаляется в виде воды с помощью Glu. 211 , и формируется PEP.
Кроме того, внутри фермента происходят конформационные изменения, которые способствуют катализу. В α-енолазе человека субстрат поворачивается в нужное положение при связывании с ферментом за счет взаимодействия с двумя каталитическими ионами магния, Gln. 167 и Лис 396 . Движения петель Сер 36 Его 43 , Быть 158 в Гли 162 и Асп 255 в Асн 256 разрешить Серу 39 согласовать с Мг 2+ и закройте активный сайт. Помимо координации с каталитическими ионами магния, pKa альфа-водорода субстрата также снижается из-за протонирования фосфорильной группы His. 159 и его близость к Аргу 374 . Арг 374 также вызывает Лис 345 в активном центре депротонируется, что запускает Lys 345 за его роль в механизме.
Диагностическое использование [ править ]
В недавних медицинских экспериментах концентрации енолазы были отобраны в попытке диагностировать определенные состояния и их тяжесть. Например, более высокие концентрации енолазы в спинномозговой жидкости низкой степени злокачественности, более сильно коррелировали с астроцитомой чем другие протестированные ферменты ( альдолаза , пируваткиназа , креатинкиназа и лактатдегидрогеназа ). [16] То же исследование показало, что самая быстрая скорость роста опухоли наблюдалась у пациентов с самым высоким уровнем энолазы спинномозговой жидкости.Повышенные уровни енолазы также были выявлены у пациентов, недавно перенесших инфаркт миокарда или нарушение мозгового кровообращения . Был сделан вывод, что уровни нейрон-специфической енолазы спинномозговой жидкости, сывороточного NSE и креатинкиназы (тип BB) являются показательными для прогностической оценки пострадавших от остановки сердца. [17] Другие исследования были сосредоточены на прогностической ценности значений NSE у жертв мозгового кровообращения. [18]
Аутоантитела к альфа-енолазе связаны с ревматоидным артритом. [19] и редкий синдром, называемый энцефалопатией Хашимото . [20]
Ингибиторы [ править ]
Низкомолекулярные ингибиторы енолазы были синтезированы как химические зонды (субстраты-аналоги) каталитического механизма фермента, а в последнее время исследовались как потенциальные средства лечения рака и инфекционных заболеваний. [21] [22] Большинство ингибиторов обладают металлохелатирующими свойствами и связываются с ферментом путем взаимодействия со структурным атомом магния Mg(A). [23] [24] Наиболее активным из них является фосфоноацетогидроксамат. [24] который в своей непротонированной форме имеет сродство к ферменту pM. Он имеет структурное сходство с предполагаемым каталитическим промежуточным продуктом между PEP и 2-PG. Были предприняты попытки использовать этот ингибитор в качестве препарата против трипаносом. [25] и в последнее время в качестве противоракового средства, в частности, при глиобластоме с дефицитом енолазы из-за гомозиготной делеции гена ENO1 как части локуса-супрессора опухоли 1p36 ( синтетическая летальность ). [26] Натуральный фосфонатный антибиотик SF2312 ( CAS 107729-45-3), активный против грамположительных и отрицательных бактерий, особенно в анаэробных условиях. [27] является высокоэффективным ингибитором енолазы 4zcw , который связывается аналогично фосфоноацетогидроксамату 4za0 . [28] SF2312 ингибирует активность енолазы как эукариотического , так и прокариотического происхождения. [29] что отражает сильную эволюционную консервативность енолазы и древнее происхождение пути гликолиза. SF2312 представляет собой хиральную молекулу, содержащую только 3S-энантиомер, проявляющую ингибирующую енолазу активность и биологическую активность в отношении бактерий. [30] Совсем недавно было показано, что производное SF2312, названное HEX, и его пролекарство POMHEX проявляют противоопухолевую активность против глиомы с делецией ENO1 на доклинической внутричерепной ортотопической мышиной модели. [31] Аллостерическое связующее, ENOblock. [22] Первоначально был описан как ингибитор енолазы, но впоследствии было показано, что он на самом деле не ингибирует фермент, а, скорее, мешает ферментативному анализу енолазы in vitro. [32] Было обнаружено, что ENOblock изменяет клеточную локализацию енолазы, влияя на ее вторичные, негликолитические функции, такие как регуляция транскрипции. [33] Последующий анализ с использованием коммерческого анализа также показал, что ENOblock может ингибировать активность енолазы в биологических средах, таких как клетки и ткани животных. [33] Метилглиоксаль также описан как ингибитор енолазы человека. [34]
Ингибиторы энолазы с переходным состоянием активного сайта были исследованы доклинически для лечения различных микробных патогенов, а также в прецизионной онкологии для опухолей с гомозиготными делециями 1p36, в которых отсутствует ENO1. [31] [35] [36] [37] [38] [39] [40]
Фторид является известным конкурентом субстрата енолазы 2-PG. Фторид может образовывать комплекс с магнием и фосфатом, который связывается в активном центре вместо 2-PG. [4] Одно исследование показало, что фторид может ингибировать бактериальную енолазу in vitro . [41] Ингибирующая енолазу активность аниона фтора может способствовать противокариесному эффекту фторидной зубной пасты за счет ограничения выработки молочной кислоты (продукта гликолиза, для которого требуется энолаза). [ нужна медицинская ссылка ]
Ссылки [ править ]
- ^ PDB : 2ONE ; Чжан Э., Брюэр Дж. М., Минор В., Каррейра Л. А., Лебиода Л. (октябрь 1997 г.). «Механизм енолазы: кристаллическая структура асимметричной димерной енолазы-2-фосфо-D-глицерата/енолазы-фосфоенолпирувата при разрешении 2,0 А». Биохимия . 36 (41): 12526–12534. дои : 10.1021/bi9712450 . ПМИД 9376357 .
- ^ ВВП : 2XSX ; Воллмар М., Кристофинска Э., Чайкуад А., Кройер Т., Кокинг Р., Вондельфт Ф., Бунтра С., Эроусмит CH, Вайгельт Дж., Эдвардс А., Юэ В.В., Опперманн Ю. (2010). «Кристаллическая структура бета-енолазы человека ENOB». Банк данных по белкам .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж Панчоли V (июнь 2001 г.). «Многофункциональная альфа-енолаза: ее роль в заболеваниях». Клеточные и молекулярные науки о жизни . 58 (7): 902–920. дои : 10.1007/pl00000910 . ПМИД 11497239 . S2CID 9191423 .
- ^ Jump up to: Перейти обратно: а б с д Хоорн Р.К., Фликверт Дж.П., Стаал Г.Е. (ноябрь 1974 г.). «Очистка и свойства енолазы эритроцитов человека». Международный журнал биохимии . 5 (11–12): 845–852. дои : 10.1016/0020-711X(74)90119-0 . hdl : 1874/18158 . S2CID 86699159 .
- ^ Ломан, К; Мейерхоф, О (1934). «О ферментативном превращении фосфоглицериновой кислоты в пировиноградную и фосфорную кислоту» [Ферментативное превращение фосфоглицериновой кислоты в пировиноградную и фосфорную кислоту]. Биохимический журнал (на немецком языке). 273 :60–72.
- ^ Jump up to: Перейти обратно: а б с д Пешавария М, День ИН (апрель 1991 г.). «Молекулярная структура гена енолазы, специфического для мышц человека (ENO3)» . Биохимический журнал . 275 (Часть 2) (Часть 2): 427–433. дои : 10.1042/bj2750427 . ПМЦ 1150071 . ПМИД 1840492 .
- ^ Эхингер С., Шуберт В.Д., Бергманн С., Хаммершмидт С., Хайнц Д.В. (октябрь 2004 г.). «Плазмин(оген)-связывающая альфа-енолаза из Streptococcus pneumoniae: кристаллическая структура и оценка сайтов связывания плазмин(оген)». Журнал молекулярной биологии . 343 (4): 997–1005. дои : 10.1016/j.jmb.2004.08.088 . ПМИД 15476816 .
- ^ Рагунатан К., Харрис П.Т., Спурбек Р.Р., Арвидсон К.Г., Арвидсон Д.Н. (июнь 2014 г.). «Кристаллическая структура эффективного ингибитора адгезии гонококков: енолаза из Lactobacillus gasseri». Письма ФЭБС . 588 (14): 2212–2216. doi : 10.1016/j.febslet.2014.05.020 . ПМИД 24859038 . S2CID 9976031 .
- ^ Диново ЕС, Бойер П.Д. (1971). «Изотопные зонды механизма енолазной реакции» . J Биол Хим . 240 (14): 4586–93. дои : 10.1016/S0021-9258(18)62051-4 .
- ^ Пойнер Р.Р., Лафлин Л.Т., Сова Г.А., Рид Г.Х. (февраль 1996 г.). «На пути к идентификации кислотно-основных катализаторов в активном центре енолазы: сравнение свойств вариантов K345A, E168Q и E211Q». Биохимия . 35 (5): 1692–1699. дои : 10.1021/bi952186y . ПМИД 8634301 .
- ^ Рид Г.Х., Пойнер Р.Р., Ларсен Т.М., Ведекинд Дж.Э., Рэймент I (декабрь 1996 г.). «Структурные и механистические исследования енолазы». Современное мнение в области структурной биологии . 6 (6): 736–743. дои : 10.1016/S0959-440X(96)80002-9 . ПМИД 8994873 .
- ^ Ведекинд Дж. Э., Рид Г. Х., Рэймент I (апрель 1995 г.). «Октаэдрическая координация на участке металла с высоким сродством в енолазе: кристаллографический анализ комплекса MgII-фермент из дрожжей при разрешении 1,9 А». Биохимия . 34 (13): 4325–4330. дои : 10.1021/bi00013a022 . ПМИД 7703246 .
- ^ Ведекинд Дж. Э., Пойнер Р. Р., Рид Г. Х., Рэймент I (август 1994 г.). «Хелирование серина 39 с Mg2+ фиксирует ворота в активном центре енолазы: структура бис(Mg2+) комплекса дрожжевой енолазы и промежуточного аналога фосфоноацетогидроксамата с разрешением 2,1-А». Биохимия . 33 (31): 9333–9342. дои : 10.1021/bi00197a038 . ПМИД 8049235 .
- ^ Ларсен Т.М., Ведекинд Дж.Э., Рэймент И., Рид Г.Х. (апрель 1996 г.). «Карбоксилатный кислород субстрата соединяет ионы магния в активном центре енолазы: структура дрожжевого фермента в комплексе с равновесной смесью 2-фосфоглицерата и фосфоенолпирувата при разрешении 1,8 А». Биохимия . 35 (14): 4349–4358. дои : 10.1021/bi952859c . ПМИД 8605183 .
- ^ Дюкеррой С., Камю С., Жанен Дж. (октябрь 1995 г.). «Рентгеновская структура и каталитический механизм енолазы омаров». Биохимия . 34 (39): 12513–12523. дои : 10.1021/bi00039a005 . ПМИД 7547999 .
- ^ Ройдс Дж. А., Тимперли В. Р., Тейлор CB (декабрь 1981 г.). «Уровни енолазы и других ферментов в спинномозговой жидкости как показатели патологических изменений» . Журнал неврологии, нейрохирургии и психиатрии . 44 (12): 1129–1135. дои : 10.1136/jnnp.44.12.1129 . ПМЦ 491233 . ПМИД 7334408 .
- ^ Ройне Р.О., Сомер Х., Касте М., Вийникка Л., Каронен С.Л. (июль 1989 г.). «Неврологический исход после внебольничной остановки сердца. Прогнозирование с помощью ферментного анализа спинномозговой жидкости». Архив неврологии . 46 (7): 753–756. дои : 10.1001/archneur.1989.00520430047015 . ПМИД 2742544 .
- ^ Хэй Э., Ройдс Дж.А., Дэвис-Джонс Г.А., Льютас Н.А., Тимперли В.Р., Тейлор CB (июль 1984 г.). «Энолаза спинномозговой жидкости при инсульте» . Журнал неврологии, нейрохирургии и психиатрии . 47 (7): 724–729. дои : 10.1136/jnnp.47.7.724 . ПМЦ 1027902 . ПМИД 6747647 .
- ^ Лундберг К., Кинлох А., Фишер Б.А., Вегнер Н., Уэйт Р., Чарльз П. и др. (октябрь 2008 г.). «Антитела к пептиду 1 цитруллинированной альфа-енолазы специфичны для ревматоидного артрита и перекрестно реагируют с бактериальной енолазой». Артрит и ревматизм . 58 (10): 3009–3019. дои : 10.1002/арт.23936 . ПМИД 18821669 .
- ^ Фуджи А., Йонеда М., Ито Т., Ямамура О., Сатоми С., Хига Х. и др. (май 2005 г.). «Аутоантитела против аминоконца альфа-енолазы являются полезным диагностическим маркером энцефалопатии Хашимото». Журнал нейроиммунологии . 162 (1–2): 130–136. дои : 10.1016/j.jneuroim.2005.02.004 . ПМИД 15833368 . S2CID 43249019 .
- ^ Андерсон В.Е., Вайс П.М., Клеланд В.В. (июнь 1984 г.). «Промежуточные аналоги реакции енолазы». Биохимия . 23 (12): 2779–2786. дои : 10.1021/bi00307a038 . ПМИД 6380574 .
- ^ Jump up to: Перейти обратно: а б Юнг Д.В., Ким В.Х., Пак Ш.Х., Ли Дж., Ким Дж., Су Д. и др. (2 апреля 2013 г.). «Уникальный низкомолекулярный ингибитор енолазы проясняет свою роль в фундаментальных биологических процессах». АКС Химическая биология . 8 (6): 1271–1282. дои : 10.1021/cb300687k . ПМИД 23547795 .
- ^ Пойнер Р.Р., Рид Г.Х. (август 1992 г.). «Структура бивалентного катионного комплекса с фосфоноацетогидроксаматом в активном центре енолазы». Биохимия . 31 (31): 7166–7173. дои : 10.1021/bi00146a020 . ПМИД 1322695 .
- ^ Jump up to: Перейти обратно: а б Чжан Э., Хатада М., Брюэр Дж. М., Лебиода Л. (май 1994 г.). «Каталитическое связывание ионов металлов в енолазе: кристаллическая структура комплекса енолаза-Mn2+-фосфоноацетогидроксамат при разрешении 2,4 А». Биохимия . 33 (20): 6295–6300. дои : 10.1021/bi00186a032 . ПМИД 8193144 .
- ^ Мюллер Ф.Л., Колла С., Аквиланти Е., Манзо В.Е., Дженовезе Г., Ли Дж. и др. (август 2012 г.). «Удаления пассажиров создают терапевтические уязвимости при раке» . Природа . 488 (7411): 337–342. Бибкод : 2012Natur.488..337M . дои : 10.1038/nature11331 . ПМЦ 3712624 . ПМИД 22895339 .
- ^ Ватанабэ Х., Ёсида Дж., Танака Э., Ито М., Миядо С., Шомура Т. (1986). «Исследование нового антибиотика на основе фосфоновой кислоты SF-2312». Научный представитель Мэйдзи Сейка Кайша . 25 : 12–17.
- ^ Леонард П.Г., Сатани Н., Максвелл Д., Лин Ю.Х., Хаммуди Н., Пэн З. и др. (декабрь 2016 г.). «SF2312 является природным фосфонатным ингибитором енолазы» . Химическая биология природы . 12 (12): 1053–1058. дои : 10.1038/nchembio.2195 . ПМК 5110371 . ПМИД 27723749 .
- ^ Кручинска Дж., Ломбардо М.Н., Эрландсен Х., Хазин А., Дуай С.С., Паттис Дж.Г. и др. (ноябрь 2019 г.). «Функциональная и структурная основа ингибирования енолазы E. coli с помощью SF2312: имитатор промежуточного карбаниона» . Научные отчеты . 9 (1): 17106. Бибкод : 2019НатСР...917106К . дои : 10.1038/s41598-019-53301-3 . ПМК 6863902 . ПМИД 31745118 .
- ^ Писанески Ф., Лин Ю.Х., Леонард П.Г., Сатани Н., Ян В.К., Хаммуди Н. и др. (июль 2019 г.). «Энантиомер 3 S стимулирует ингибирующую енолазу активность в SF2312 и его аналогах» . Молекулы . 24 (13): 2510. doi : 10,3390/molecules24132510 . ПМК 6651268 . ПМИД 31324042 .
- ^ Jump up to: Перейти обратно: а б Лин Ю.Х., Сатани Н., Хаммуди Н., Ян В.К., Барекатаин Ю., Хадка С. и др. (декабрь 2020 г.). «Ингибитор енолазы для таргетного лечения рака с удаленным ENO1» . Природный метаболизм . 2 (12): 1413–1426. дои : 10.1038/s42255-020-00313-3 . ПМЦ 7744354 . ПМИД 33230295 .
- ^ Сатани Н., Лин Й.Х., Хаммуди Н., Рагхаван С., Георгиу Д.К., Мюллер Флорида (28 декабря 2016 г.). «ЭНОблок не ингибирует активность гликолитического фермента енолазы» . ПЛОС ОДИН . 11 (12): e0168739. Бибкод : 2016PLoSO..1168739S . дои : 10.1371/journal.pone.0168739 . ПМК 5193436 . ПМИД 28030597 .
- ^ Jump up to: Перейти обратно: а б Чо Х., Ум Дж., Ли Дж.Х., Ким В.Х., Кан В.С., Ким Ш.Х. и др. (март 2017 г.). «ЭНОблок, уникальный низкомолекулярный ингибитор негликолитических функций енолазы, облегчает симптомы диабета 2 типа» . Научные отчеты . 7 : 44186. Бибкод : 2017NatSR...744186C . дои : 10.1038/srep44186 . ПМЦ 5341156 . ПМИД 28272459 .
- ^ Петкевич Дж., Гамиан А., Станишевска М., Даниэлевич Р. (апрель 2009 г.). «Ингибирование енолазы, специфичной для мышц человека, метилглиоксалем и необратимое образование конечных продуктов гликирования». Журнал ингибирования ферментов и медицинской химии . 24 (2): 356–364. дои : 10.1080/14756360802187679 . ПМИД 18830874 . S2CID 85416928 .
- ^ Ежевски А.Дж., Лин Ю.Х., Рейш Дж.А., Калп-Хилл Р., Барекатайн Ю., Ян В.К. и др. (16 сентября 2021 г.). «Нацеливание на гликолиз хозяина как стратегия противомалярийного развития» . Границы клеточной и инфекционной микробиологии . 11 : 730413. doi : 10.3389/fcimb.2021.730413 . ПМЦ 8482815 . ПМИД 34604112 .
- ^ Миллер Дж.Дж., Шах И.Т., Хаттен Дж., Барекатейн Ю., Мюллер Э.А., Мустафа А.М. и др. (июль 2021 г.). «Структурно-ориентированное микробное нацеливание антистафилококковых пролекарств» . электронная жизнь . 10 : е66657. дои : 10.7554/eLife.66657 . ПМЦ 8318587 . ПМИД 34279224 .
- ^ [Бесплатная статья PMC] [PubMed] Микати М.О., Миллер Дж.Дж., Осборн Д.М., Барекатайн Ю., Гебремайкл Н., Шах И.Т. и др. (ноябрь 2020 г.). «Активация противомикробного пролекарства стафилококковой глиоксалазой GloB» . ОКС Инфекционные болезни . 6 (11): 3064–3075. doi : 10.1021/acsinfecdis.0c00582 . ПМЦ 8543975 . ПМИД 33118347 . S2CID 226052354 .
- ^ Майтитуохети М., Кеунг Э.З., Тан М., Ян Л., Алам Х., Хан Г. и др. (октябрь 2020 г.). «Перепрограммирование энхансеров приводит к зависимости от гликолиза и передачи сигналов IGF при мутантной меланоме KMT2D» . Отчеты по ячейкам . 33 (3): 108293. doi : 10.1016/j.celrep.2020.108293 . ПМЦ 7649750 . ПМИД 33086062 .
- ^ Кручинска Дж., Ломбардо М.Н., Эрландсен Х., Хазин А., Дуай С.С., Паттис Дж.Г. и др. (ноябрь 2019 г.). «Функциональная и структурная основа ингибирования енолазы E. coli с помощью SF2312: имитатор промежуточного карбаниона» . Научные отчеты . 9 (1): 17106. Бибкод : 2019НатСР...917106К . дои : 10.1038/s41598-019-53301-3 . ПМК 6863902 . ПМИД 31745118 .
- ^ Писанески Ф., Лин Ю.Х., Леонард П.Г., Сатани Н., Ян В.К., Хаммуди Н. и др. (июль 2019 г.). «Энантиомер 3 S стимулирует ингибирующую енолазу активность в SF2312 и его аналогах» . Молекулы . 24 (13): 2510. doi : 10,3390/molecules24132510 . ПМК 6651268 . ПМИД 31324042 .
- ^ Хютер Ф.Дж., Псаррос Н., Душнер Х. (апрель 1990 г.). «Выделение, характеристика и кинетика ингибирования енолазы из Streptococcus rattus FA-1» . Инфекция и иммунитет . 58 (4): 1043–1047. дои : 10.1128/IAI.58.4.1043-1047.1990 . ПМК 258580 . ПМИД 2318530 .
Дальнейшее чтение [ править ]
- Холт А., Уолд Ф. (декабрь 1961 г.). «Выделение и характеристика енолазы мышц кролика» . Журнал биологической химии . 236 (12): 3227–3231. дои : 10.1016/S0021-9258(18)94000-7 . ПМИД 13908561 .
- Бойер П.Д., Ларди Х. и Мирбак К. (ред.), Ферменты, 2-е изд., том. 5, Academic Press, Нью-Йорк, 1961, с. 471-494.
- Вестхед EW, Маклейн Дж. (август 1964 г.). «Очистка енолазы пивных и пекарских дрожжей с получением единственного активного компонента» . Журнал биологической химии . 239 (8): 2464–2468. дои : 10.1016/S0021-9258(18)93875-5 . ПМИД 14235523 .
Внешние ссылки [ править ]
- Энолаза в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)









