Пируваткиназа
| Пируваткиназа | |||
|---|---|---|---|
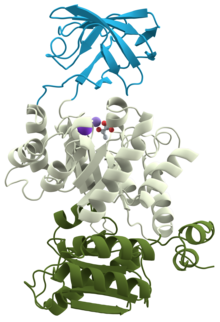 3D-структура пируваткиназы ( 1PKN ) | |||
| Идентификаторы | |||
| Номер ЕС. | 2.7.1.40 | ||
| Номер CAS. | 9001-59-6 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
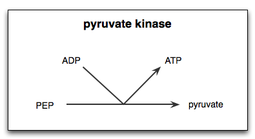
Пируваткиназа — фермент, участвующий в последней стадии гликолиза . Он катализирует перенос фосфатной группы от фосфоенолпирувата (ФЕП) к аденозиндифосфату (АДФ), образуя одну молекулу пирувата и одну молекулу АТФ . [1] Пируваткиназа получила неправильное название (несовместимое с обычной киназой ) до того, как было признано, что она не катализирует напрямую фосфорилирование пирувата , которое не происходит в физиологических условиях. [2] Пируваткиназа присутствует у животных в виде четырех различных тканеспецифичных изоферментов, каждый из которых обладает определенными кинетическими свойствами, необходимыми для адаптации к изменениям метаболических потребностей различных тканей.
у Изоферменты позвоночных
У позвоночных экспрессируются четыре изозима пируваткиназы: L (печень), R (эритроциты), M1 (мышцы и мозг) и M2 (ткани раннего плода и большинство тканей взрослого человека). Изоферменты L и R экспрессируются геном PKLR , тогда как изоферменты M1 и M2 экспрессируются геном PKM2 . Изоферменты R и L отличаются от M1 и M2 тем, что они регулируются аллостерически. Кинетически изоферменты R и L пируваткиназы имеют два различных конформационных состояния; один с высоким сродством к субстрату и один с низким сродством к субстрату. R-состояние, характеризующееся высоким сродством к субстрату, служит активированной формой пируваткиназы и стабилизируется PEP и фруктозо-1,6-бисфосфатом (FBP), способствуя гликолитическому пути. Т-состояние, характеризующееся низким сродством к субстрату, служит инактивированной формой пируваткиназы, связанной и стабилизированной АТФ и аланином , вызывая фосфорилирование пируваткиназы и ингибирование гликолиза. [3] Изофермент М2 пируваткиназы может образовывать тетрамеры или димеры. Тетрамеры обладают высоким сродством к PEP, тогда как димеры имеют низкое сродство к PEP. Ферментативную активность можно регулировать путем фосфорилирования высокоактивных тетрамеров ПКМ2 в неактивные димеры. [4]
Ген ПКМ состоит из 12 экзонов и 11 интронов . PKM1 и PKM2 представляют собой разные продукты сплайсинга M-гена (PKM1 содержит экзон 9, а PKM2 содержит экзон 10) и различаются только по 23 аминокислотам в пределах участка из 56 аминокислот (378-434 а.к.) на их карбокси-конце . [5] [6] Ген PKM регулируется посредством гетерогенных рибонуклеотидных белков, таких как hnRNPA1 и hnRNPA2. [7] Мономер PKM2 человека содержит 531 аминокислоту и представляет собой одну цепь, разделенную на домены A, B и C. Разница в аминокислотной последовательности между PKM1 и PKM2 позволяет PKM2 аллостерически регулироваться с помощью FBP и образовывать димеры и тетрамеры, тогда как PKM1 может образовывать только тетрамеры. [8]
Изоферменты бактерий [ править ]
Многие Enterobacteriaceae, включая E. coli , имеют две изоформы пируваткиназы, PykA и PykF, которые на 37% идентичны у E. coli (Uniprot: PykA , PykF ). Они катализируют ту же реакцию, что и у эукариот, а именно образование АТФ из АДФ и ПЭП, последнюю стадию гликолиза , стадию, необратимую в физиологических условиях. PykF аллостерически регулируется FBP, что отражает центральное положение PykF в клеточном метаболизме. [9] Транскрипция PykF в E. coli регулируется глобальным регулятором транскрипции Cra (FruR). [10] [11] [12] Было показано, что PfkB ингибируется MgATP при низких концентрациях Fru-6P, и эта регуляция важна для глюконеогенеза . [13]
Реакция [ править ]
Гликолиз [ править ]
Реакция пируваткиназы при гликолизе состоит из двух стадий. Во-первых, PEP переносит фосфатную группу на ADP, образуя АТФ и енолят пирувата. Во-вторых, к еноляту пирувата необходимо добавить протон, чтобы получить функциональную форму пирувата, необходимую клетке. [14] Поскольку субстратом пируваткиназы является простой фосфосахар, а продуктом является АТФ, пируваткиназа является возможным основным ферментом для эволюции цикла гликолиза и может быть одним из самых древних ферментов во всей земной жизни. . Фосфоенолпируват мог присутствовать абиотически, и было показано, что он образуется с высоким выходом по примитивному пути триозогликолиза. [15]
Было обнаружено , что в дрожжевых клетках взаимодействие дрожжевой пируваткиназы (YPK) с PEP и его аллостерическим эффектором фруктозо-1,6-бисфосфатом (FBP) усиливается присутствием Mg. 2+ . Следовательно, Мг 2+ Был сделан вывод, что он является важным кофактором в катализе PEP в пируват пируваткиназой. Кроме того, ион металла Mn 2+ Было показано, что он оказывает аналогичный, но более сильный эффект на YPK, чем Mg. 2+ . Связывание ионов металлов с металлосвязывающими сайтами пируваткиназы увеличивает скорость этой реакции. [16]
Реакция, катализируемая пируваткиназой, является заключительной стадией гликолиза. Это один из трех этапов этого пути, ограничивающих скорость. Ограничивающие скорость этапы — это более медленные, регулируемые этапы пути, которые, таким образом, определяют общую скорость пути. В гликолизе стадии, ограничивающие скорость, связаны либо с гидролизом АТФ, либо с фосфорилированием АДФ, в результате чего этот путь становится энергетически выгодным и по существу необратимым в клетках. Этот последний этап строго регулируется и намеренно необратим, поскольку пируват является важнейшим промежуточным строительным блоком для дальнейших метаболических путей. [17] После образования пирувата он либо поступает в цикл ТСА для дальнейшего производства АТФ в аэробных условиях, либо превращается в молочную кислоту или этанол в анаэробных условиях.
реакция обратная Глюконеогенез :
Пируваткиназа также служит регуляторным ферментом глюконеогенеза — биохимического пути, в котором печень генерирует глюкозу из пирувата и других субстратов. Глюконеогенез использует неуглеводные источники для снабжения глюкозой мозга и эритроцитов во время голодания, когда прямые резервы глюкозы истощены. [17] Во время голодания пируваткиназа ингибируется, что предотвращает «утечку» фосфоенолпирувата от превращения в пируват; [17] вместо этого фосфоенолпируват превращается в глюкозу посредством каскада реакций глюконеогенеза . Хотя глюконеогенез использует аналогичные ферменты, он не является обратным гликолизу. Напротив, это путь, который обходит необратимые этапы гликолиза. Более того, глюконеогенез и гликолиз не происходят в клетке одновременно в любой момент, поскольку они взаимно регулируются передачей сигналов в клетках. [17] После завершения пути глюконеогенеза вырабатываемая глюкоза выводится из печени, обеспечивая энергию для жизненно важных тканей в состоянии голодания.
Регламент [ править ]
Гликолиз строго регулируется на трех его каталитических стадиях: фосфорилировании глюкозы гексокиназой , фосфорилировании фруктозо-6-фосфата и фосфофруктокиназой переносе фосфата от PEP к ADP пируваткиназой. В условиях дикого типа все три эти реакции необратимы, имеют большую отрицательную свободную энергию и отвечают за регуляцию этого пути. [17] Активность пируваткиназы наиболее широко регулируется аллостерическими эффекторами, ковалентными модификаторами и гормональным контролем. Однако наиболее значимым регулятором пируваткиназы является фруктозо-1,6-бисфосфат (FBP), который служит аллостерическим эффектором фермента.
Аллостерические эффекторы [ править ]
Аллостерическая регуляция — это связывание эффектора с участком белка, отличным от активного сайта, вызывающее конформационные изменения и изменяющее активность данного белка или фермента. Было обнаружено, что пируваткиназа аллостерически активируется FBP и аллостерически инактивируется АТФ и аланином. [18] Тетрамеризации пируваткиназы способствуют FBP и серин, тогда как диссоциации тетрамера способствует L-цистеин. [19] [20] [21]
Фруктозо-1,6-бисфосфат [ править ]
FBP является наиболее важным источником регуляции, поскольку он происходит на пути гликолиза. FBP представляет собой гликолитический промежуточный продукт, образующийся в результате фосфорилирования фруктозо-6-фосфата . FBP связывается с аллостерическим сайтом связывания домена C пируваткиназы и изменяет конформацию фермента, вызывая активацию активности пируваткиназы. [22] В качестве промежуточного продукта гликолитического пути FBP обеспечивает прямую стимуляцию , поскольку чем выше концентрация FBP, тем выше аллостерическая активация и величина активности пируваткиназы. Пируваткиназа наиболее чувствительна к воздействию FBP. В результате остальные регуляторные механизмы служат вторичной модификацией. [9] [23]
Ковалентные модификаторы [ править ]
Ковалентные модификаторы служат косвенными регуляторами, контролируя фосфорилирование, дефосфорилирование, ацетилирование, сукцинилирование и окисление ферментов, что приводит к активации и ингибированию ферментативной активности. [24] В печени глюкагон и адреналин активируют протеинкиназу А , которая служит ковалентным модификатором путем фосфорилирования и деактивации пируваткиназы. Напротив, секреция инсулина в ответ на повышение уровня сахара в крови активирует фосфопротеинфосфатазу I, вызывая дефосфорилирование и активацию пируваткиназы для усиления гликолиза. Та же ковалентная модификация оказывает противоположное действие на ферменты глюконеогенеза. Эта система регуляции отвечает за предотвращение бесполезного цикла посредством предотвращения одновременной активации пируваткиназы и ферментов, катализирующих глюконеогенез. [25]
Гормональный контроль [ править ]
Чтобы предотвратить бесполезный цикл , гликолиз и глюконеогенез строго регулируются, чтобы гарантировать, что они никогда не действуют в клетке одновременно. В результате ингибирование пируваткиназы глюкагоном, циклическим АМФ и адреналином не только останавливает гликолиз, но и стимулирует глюконеогенез. Альтернативно, инсулин препятствует действию глюкагона, циклического АМФ и адреналина, вызывая нормальное функционирование пируваткиназы и остановку глюконеогенеза. Кроме того, было обнаружено, что глюкоза ингибирует и нарушает глюконеогенез, не затрагивая активность пируваткиназы и гликолиз. В целом взаимодействие между гормонами играет ключевую роль в функционировании и регуляции гликолиза и глюконеогенеза в клетке. [26]
метформина эффект Тормозящий
Метформин, или диметилбигуанид , является основным средством лечения диабета 2 типа. Было показано, что метформин косвенно влияет на пируваткиназу посредством ингибирования глюконеогенеза. В частности, добавление метформина связано с заметным снижением потока глюкозы и увеличением потока лактата/пирувата по различным метаболическим путям. Хотя метформин не влияет непосредственно на активность пируваткиназы, он вызывает снижение концентрации АТФ. Из-за аллостерического ингибирующего действия АТФ на пируваткиназу снижение уровня АТФ приводит к уменьшению ингибирования и последующей стимуляции пируваткиназы. Следовательно, увеличение активности пируваткиназы направляет метаболический поток через гликолиз, а не глюконеогенез. [27]
генов Регуляция
Гетерогенные рибонуклеотидные белки (hnRNP) могут действовать на ген PKM, регулируя экспрессию изоформ M1 и M2. Изоформы PKM1 и PKM2 представляют собой сплайсинговые варианты гена PKM, отличающиеся одним экзоном. Различные типы hnRNP, такие как hnRNPA1 и hnRNPA2, проникают в ядро во время условий гипоксии и модулируют экспрессию, так что PKM2 активируется. [28] Гормоны, такие как инсулин, повышают экспрессию PKM2, в то время как гормоны, такие как трийодтиронин (Т3) и глюкагон, способствуют снижению экспрессии PKM2. [29]
связывающий элементы ответа на углеводы ( ChREBP Белок , )
ChREBP представляет собой фактор транскрипции , который регулирует экспрессию L-изозима пируваткиназы. [30] Модуль, чувствительный к глюкозе, содержит домены, которые являются мишенями для регуляторного фосфорилирования в зависимости от концентрации глюкозы и цАМФ, которые затем контролируют ее импорт в ядро. [31] Его также можно дополнительно активировать путем прямого связывания глюкозо-6-фосфата. [30] [32] Попав в ядро, его ДНК-связывающие домены активируют транскрипцию пируваткиназы. [31] Следовательно, высокий уровень глюкозы и низкий уровень цАМФ вызывают дефосфорилирование ChREBP , что затем усиливает экспрессию пируваткиназы в печени. [30]
применения Клинические
Дефицит [ править ]
Генетические дефекты этого фермента вызывают заболевание, известное как дефицит пируваткиназы . В этом состоянии недостаток пируваткиназы замедляет процесс гликолиза. Этот эффект особенно разрушительен для клеток, в которых отсутствуют митохондрии , поскольку эти клетки должны использовать анаэробный гликолиз в качестве единственного источника энергии, поскольку цикл ТЦА недоступен. Например, эритроциты , которые в состоянии дефицита пируваткиназы, быстро испытывают дефицит АТФ и могут подвергаться гемолизу . Следовательно, дефицит пируваткиназы может вызвать хроническую несфероцитарную гемолитическую анемию (CNSHA). [33]
- Мутация LR гена PK
Дефицит пируваткиназы обусловлен аутосомно-рецессивным признаком. У млекопитающих есть два гена пируваткиназы: PK-LR (который кодирует изоферменты пируваткиназы L и R) и PK-M (который кодирует изофермент пируваткиназы M1), но только PKLR кодирует изофермент красной крови, который вызывает дефицит пируваткиназы. Было идентифицировано более 250 мутаций гена PK-LR, связанных с дефицитом пируваткиназы. Тестирование ДНК способствовало открытию местоположения PKLR на хромосоме 1 и разработке тестов прямого секвенирования генов для молекулярной диагностики дефицита пируваткиназы. [34]
пируваткиназы ингибирования Применение
форм кислорода ( Ингибирование активных ) АФК
Активные формы кислорода (АФК) представляют собой химически активные формы кислорода. Было показано, что в клетках легких человека АФК ингибируют изофермент М2 пируваткиназы (PKM2). АФК достигают этого ингибирования путем окисления Cys358 и инактивации PKM2. В результате инактивации PKM2 поток глюкозы больше не превращается в пируват, а вместо этого используется в пентозофосфатном пути, что приводит к снижению и детоксикации АФК. Таким образом, вредное воздействие АФК усиливается и вызывает больший окислительный стресс на клетках легких, что приводит к потенциальному образованию опухолей. Этот ингибирующий механизм важен, поскольку он может указывать на то, что регуляторные механизмы PKM2 ответственны за содействие устойчивости раковых клеток к окислительному стрессу и усилению онкогенеза. [35] [36]
Ингибирование фенилаланина
Обнаружено, что фенилаланин действует как конкурентный ингибитор пируваткиназы в мозге. Хотя степень ингибирующей активности фенилаланина одинакова как в клетках плода, так и в клетках взрослого человека, ферменты в клетках мозга плода значительно более уязвимы к ингибированию, чем ферменты в клетках мозга взрослого человека. Исследование ПКМ2 у детей с генетическим заболеванием головного мозга фенилкетонурией (ФКУ) показало повышенный уровень фенилаланина и снижение эффективности ПКМ2. Этот ингибирующий механизм дает представление о роли пируваткиназы в повреждении клеток головного мозга. [37] [38]
Пируваткиназа раке при
Раковые клетки характеризуются ускорением метаболизма, и считается, что пируваткиназа играет роль в развитии рака. По сравнению со здоровыми клетками, раковые клетки имеют повышенные уровни изоформы PKM2, особенно димера с низкой активностью. Поэтому уровни PKM2 в сыворотке используются в качестве маркеров рака. Димер с низкой активностью позволяет накапливать фосфоенолпируват (ФЕП), оставляя большие концентрации гликолитических промежуточных продуктов для синтеза биомолекул, которые в конечном итоге будут использоваться раковыми клетками. [8] Фосфорилирование PKM2 митоген-активируемой протеинкиназой 1 (ERK2) вызывает конформационные изменения, которые позволяют PKM2 проникать в ядро и регулировать экспрессию гликолитических генов, необходимую для развития опухоли. [39] Некоторые исследования утверждают, что во время канцерогенеза происходит сдвиг экспрессии с PKM1 на PKM2. Микроокружение опухоли, такое как гипоксия, активирует факторы транскрипции, такие как фактор, индуцируемый гипоксией, для стимулирования транскрипции PKM2, который образует петлю положительной обратной связи для усиления собственной транскрипции. [8]
Альтернативы [ править ]
Обратимый фермент с аналогичной функцией, пируватфосфатдикиназа (PPDK), обнаружен у некоторых бактерий и перенесен в ряд анаэробных групп эукариот (например, Streblomastix , Giardia , Entamoeba и Trichomonas ), по-видимому, через горизонтальный ген. переводить два или более раз. В некоторых случаях в одном и том же организме присутствуют как пируваткиназа, так и ППДК. [40]
Ссылки [ править ]
- ^ Гупта В., Бамезай Р.Н. (ноябрь 2010 г.). «Пируваткиназа М2 человека: многофункциональный белок» . Белковая наука . 19 (11): 2031–44. дои : 10.1002/pro.505 . ПМК 3005776 . ПМИД 20857498 .
- ^ Гудман Х.М. (2009). Базовая медицинская эндокринология (4-е изд.). Эльзевир. п. 132 . ISBN 978-0-12-373975-9 .
- ^ Мюрхед Х. (апрель 1990 г.). «Изоферменты пируваткиназы». Труды Биохимического общества . 18 (2): 193–6. дои : 10.1042/bst0180193 . ПМИД 2379684 . S2CID 3262531 .
- ^ Эйгенбродт Э., Райнахер М., Шиферс-Борчел Ю., Шиферс Х., Фриис Р. (1 января 1992 г.). «Двойная роль пируваткиназы типа М2 в расширении пулов фосфометаболитов, обнаруженных в опухолевых клетках». Критические обзоры онкогенеза . 3 (1–2): 91–115. ПМИД 1532331 .
- ^ Ногучи Т., Иноуэ Х., Танака Т. (октябрь 1986 г.). «Изоферменты пируваткиназы крысы M1 и M2 производятся из одного и того же гена путем альтернативного сплайсинга РНК» . Журнал биологической химии . 261 (29): 13807–12. дои : 10.1016/S0021-9258(18)67091-7 . ПМИД 3020052 .
- ^ Домбраукас Дж.Д., Сантарсьеро Б.Д., Месекар А.Д. (июль 2005 г.). «Структурная основа аллостерической регуляции и катализа опухолевой пируваткиназы М2». Биохимия . 44 (27): 9417–29. дои : 10.1021/bi0474923 . ПМИД 15996096 . S2CID 24625677 .
- ^ Чен М., Чжан Дж., Мэнли Дж.Л. (ноябрь 2010 г.). «Включение топливного переключателя рака: белки hnRNP регулируют альтернативный сплайсинг мРНК пируваткиназы» . Исследования рака . 70 (22): 8977–80. дои : 10.1158/0008-5472.CAN-10-2513 . ПМЦ 2982937 . ПМИД 20978194 .
- ↑ Перейти обратно: Перейти обратно: а б с Пракасам Г., Икбал М.А., Бамезай Р.Н., Мазурек С. (2018). «Посттрансляционные модификации пируваткиназы M2: изменения, приносящие пользу раку» . Границы онкологии . 8:22 . doi : 10.3389/fonc.2018.00022 . ПМЦ 5808394 . ПМИД 29468140 .
- ↑ Перейти обратно: Перейти обратно: а б Валентини Дж., Кьярелли Л., Фортин Р., Сперанца М.Л., Галицци А., Маттеви А. (июнь 2000 г.). «Аллостерическая регуляция пируваткиназы» . Журнал биологической химии . 275 (24): 18145–52. дои : 10.1074/jbc.M001870200 . ПМИД 10751408 .
- ^ Рамсейер Т.М., Негре Д., Кортай Дж.К., Скарабель М., Коццоне А.Дж., Сайер М.Х. (ноябрь 1993 г.). «Связывание плейотропного регулятора транскрипции FruR in vitro с оперонами fru, pps, ace, pts и icd Escherichia coli и Salmonella typhimurium». Журнал молекулярной биологии . 234 (1): 28–44. дои : 10.1006/jmbi.1993.1561 . ПМИД 8230205 .
- ^ Рамсейер Т.М., Бледиг С., Мичоти В., Фегхали Р., Сайер М.Х. (июнь 1995 г.). «Глобальный регуляторный белок FruR модулирует направление потока углерода в Escherichia coli». Молекулярная микробиология . 16 (6): 1157–69. дои : 10.1111/j.1365-2958.1995.tb02339.x . ПМИД 8577250 . S2CID 45447144 .
- ^ Сайер М.Х., Рамсейер Т.М. (июнь 1996 г.). «Белок-репрессор/активатор катаболитов (Cra) кишечных бактерий» . Журнал бактериологии . 178 (12): 3411–7. дои : 10.1128/jb.178.12.3411-3417.1996 . ПМК 178107 . ПМИД 8655535 .
- ^ Сабнис Н.А., Ян Х., Ромео Т. (декабрь 1995 г.). «Плейотропная регуляция центрального углеводного обмена у Escherichia coli посредством гена csrA» . Журнал биологической химии . 270 (49): 29096–104. дои : 10.1074/jbc.270.49.29096 . ПМИД 7493933 .
- ^ Кумар С., Барт А. (май 2010 г.). «Связывание фосфоенолпирувата и Mg2+ с пируваткиназой, контролируемое с помощью инфракрасной спектроскопии» . Биофизический журнал . 98 (9): 1931–40. Бибкод : 2010BpJ....98.1931K . дои : 10.1016/j.bpj.2009.12.4335 . ПМК 2862152 . ПМИД 20441757 .
- ^ Коггинс AJ, владелец MW (апрель 2017 г.). «Пребиотический синтез фосфоенолпирувата путем контролируемого α-фосфорилированием триозогликолиза» . Природная химия . 9 (4): 310–317. дои : 10.1038/nchem.2624 . ПМИД 28338685 . S2CID 205296677 .
- ^ Болленбах Т.Дж., Новак Т. (октябрь 2001 г.). «Анализ кинетических связанных функций мультилигандных взаимодействий на Mg (2+)-активированной дрожжевой пируваткиназы». Биохимия . 40 (43): 13097–106. дои : 10.1021/bi010126o . ПМИД 11669648 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Берг Дж. М., Тимочко Дж. Л., Страйер Дж., Кларк Н. Д. (2002). Биохимия (пятое изд.). Нью-Йорк, штат Нью-Йорк: WH Freeman. ISBN 978-0-7167-3051-4 .
- ^ Карбонелл Дж., Фелиу Дж.Е., Марко Р., Солс А. (август 1973 г.). «Пируваткиназа. Классы регуляторных изоферментов в тканях млекопитающих». Европейский журнал биохимии . 37 (1): 148–56. дои : 10.1111/j.1432-1033.1973.tb02969.x . hdl : 10261/78345 . ПМИД 4729424 .
- ^ Ян Дж, Лю Х, Лю X, Гу С, Луо Р, Чен Х.Ф. (июнь 2016 г.). «Синергетический аллостерический механизм фруктозо-1,6-бисфосфата и серина для пируваткиназы M2 с помощью сетевого анализа динамических колебаний» . Журнал химической информации и моделирования . 56 (6): 1184–1192. doi : 10.1021/acs.jcim.6b00115 . ПМК 5115163 . ПМИД 27227511 .
- ^ Чанетон Б., Хиллманн П., Чжэн Л., Мартин А.С., Мэддокс О.Д., Чоккатукалам А. и др. (ноябрь 2012 г.). «Серин является природным лигандом и аллостерическим активатором пируваткиназы М2» . Природа . 491 (7424): 458–462. Бибкод : 2012Natur.491..458C . дои : 10.1038/nature11540 . ПМЦ 3894725 . ПМИД 23064226 .
- ^ Накацу Д., Хориучи Ю., Кано Ф., Ногучи Ю., Сугавара Т., Такамото И. и др. (март 2015 г.). «L-цистеин обратимо ингибирует вызванную глюкозой двухфазную секрецию инсулина и выработку АТФ, инактивируя PKM2» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (10): Е1067-76. Бибкод : 2015PNAS..112E1067N . дои : 10.1073/pnas.1417197112 . ПМК 4364213 . ПМИД 25713368 .
- ^ Ишвар А (24 февраля 2015 г.). «Выявление взаимодействий в сайте связывания фруктозо-1,6-бисфосфата пируваткиназы печени человека, которые способствуют аллостерии» . Биохимия . 54 (7): 1516–24. дои : 10.1021/bi501426w . ПМЦ 5286843 . ПМИД 25629396 .
- ^ Юрица М.С., Месекар А., Хит П.Дж., Ши В., Новак Т., Стоддард Б.Л. (февраль 1998 г.). «Аллостерическая регуляция пируваткиназы с помощью фруктозо-1,6-бисфосфата» . Структура . 6 (2): 195–210. дои : 10.1016/S0969-2126(98)00021-5 . ПМИД 9519410 .
- ^ Ли Ю.Х., Ли К.Ф., Лю Дж.Т., Ван Х., Фань Л.Л., Ли Дж., Сунь Г.П. (август 2018 г.). «PKM2, потенциальная мишень для регулирования рака». Джин . 668 : 48–53. дои : 10.1016/j.gene.2018.05.038 . ПМИД 29775756 . S2CID 205030574 .
- ^ Бирнбаум М.Дж., Фейн Дж.Н. (январь 1977 г.). «Активация протеинкиназы и гликогенфосфорилазы в изолированных клетках печени крысы глюкагоном и катехоламинами» . Журнал биологической химии . 252 (2): 528–35. дои : 10.1016/S0021-9258(17)32749-7 . ПМИД 188818 .
- ^ Фелиу Ж.Э., Хюэ Л., Херс Х.Г. (1976). «Гормональный контроль активности пируваткиназы и глюконеогенеза в изолированных гепатоцитах» . Труды Национальной академии наук Соединенных Штатов Америки . 73 (8): 2762–6. Бибкод : 1976PNAS...73.2762F . дои : 10.1073/pnas.73.8.2762 . ПМК 430732 . ПМИД 183209 .
- ^ Арго Д., Рот Х., Вирнспергер Н., Леверв Х.М. (1993). «Метформин снижает глюконеогенез за счет усиления потока пируваткиназы в изолированных гепатоцитах крысы» . Европейский журнал биохимии . 213 (3): 1341–8. дои : 10.1111/j.1432-1033.1993.tb17886.x . ПМИД 8504825 .
- ^ Клоуэр К.В., Чаттерджи Д., Ван З., Кэнтли Л.К., Вандер Хайден М.Г., Крайнер А.Р. (февраль 2010 г.). «Альтернативные репрессоры сплайсинга hnRNP A1/A2 и PTB влияют на экспрессию изоформы пируваткиназы и клеточный метаболизм» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (5): 1894–9. Бибкод : 2010PNAS..107.1894C . дои : 10.1073/pnas.0914845107 . ПМЦ 2838216 . ПМИД 20133837 .
- ^ Икбал М.А., Сиддики Ф.А., Гупта В., Чаттопадхьяй С., Гопинатх П., Кумар Б. и др. (июль 2013 г.). «Инсулин усиливает метаболические способности раковых клеток за счет двойной регуляции гликолитического фермента пируваткиназы М2» . Молекулярный рак . 12 (1): 72. дои : 10.1186/1476-4598-12-72 . ПМК 3710280 . ПМИД 23837608 .
- ↑ Перейти обратно: Перейти обратно: а б с Кавагути Т., Такэношита М., Кабашима Т., Уеда К. (ноябрь 2001 г.). «Глюкоза и цАМФ регулируют ген пируваткиназы L-типа путем фосфорилирования/дефосфорилирования белка, связывающего элемент углеводного ответа» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (24): 13710–5. Бибкод : 2001PNAS...9813710K . дои : 10.1073/pnas.231370798 . ПМК 61106 . ПМИД 11698644 .
- ↑ Перейти обратно: Перейти обратно: а б Ортега-Прието, Паула; Постич, Кэтрин (2019). «Ощущение углеводов посредством фактора транскрипции ChREBP» . Границы генетики . 10 : 472. дои : 10.3389/fgene.2019.00472 . ISSN 1664-8021 . ПМК 6593282 . ПМИД 31275349 .
- ^ Ричардс, Пол; Ураба, Сара; Монтань, Жак; Бёрноль, Анн-Франсуаза; Постик, Кэтрин; Гильмо, Сандра (2017). «MondoA/ChREBP: обычные подозреваемые в транскрипционном распознавании глюкозы; значение в патофизиологии» . Метаболизм: клинический и экспериментальный . 70 : 133–151. дои : 10.1016/j.metabol.2017.01.033 . ISSN 1532-8600 . ПМИД 28403938 .
- ^ Грейс РФ, Занелла А., Нойфельд Э.Дж., Мортон Д.Х., Эбер С., Яиш Х., Глэйдер Б. (сентябрь 2015 г.). «Дефицит пируваткиназы эритроцитов: отчет о состоянии дел за 2015 год» . Американский журнал гематологии . 90 (9): 825–30. дои : 10.1002/ajh.24088 . ПМЦ 5053227 . ПМИД 26087744 .
- ^ Климент Ф, Розет Ф, Реписо А, Перес де ла Осса П (июнь 2009 г.). «Нарушения гликолитических ферментов эритроцитов, вызванные мутациями: обновленная информация». Целевые лекарственные средства для лечения сердечно-сосудистых и гематологических заболеваний . 9 (2): 95–106. дои : 10.2174/187152909788488636 . ПМИД 19519368 .
- ^ Анастасиу Д., Пулогианнис Г., Асара Дж.М., Боксер М.Б., Цзян Дж.К., Шен М., Беллинджер Г., Сасаки А.Т., Локасейл Дж.В., Олд Д.С., Томас С.Дж., Вандер Хайден М.Г., Кэнтли Л.С. (декабрь 2011 г.). «Ингибирование пируваткиназы М2 активными формами кислорода способствует клеточным антиоксидантным реакциям» . Наука . 334 (6060): 1278–83. Бибкод : 2011Sci...334.1278A . дои : 10.1126/science.1211485 . ПМЦ 3471535 . ПМИД 22052977 .
- ^ Кристофк Х.Р., Вандер Хайден М.Г., Харрис М.Х., Раманатан А., Герстен Р.Э., Вей Р., Флеминг М.Д., Шрайбер С.Л., Кантли Л.С. (март 2008 г.). «Изоформа сплайсинга M2 пируваткиназы важна для метаболизма рака и роста опухоли». Природа . 452 (7184): 230–3. Бибкод : 2008Natur.452..230C . дои : 10.1038/nature06734 . ПМИД 18337823 . S2CID 16111842 .
- ^ Миллер А.Л., Хокинс Р.А., Вич Р.Л. (март 1973 г.). «Фенилкетонурия: фенилаланин ингибирует пируваткиназу мозга in vivo». Наука . 179 (4076): 904–6. Бибкод : 1973Sci...179..904M . дои : 10.1126/science.179.4076.904 . ПМИД 4734564 . S2CID 12776382 .
- ^ Вебер Г. (август 1969 г.). «Ингибирование пируваткиназы и гексокиназы человеческого мозга фенилаланином и фенилпируватом: возможная связь с фенилкетонурическим повреждением головного мозга» . Труды Национальной академии наук Соединенных Штатов Америки . 63 (4): 1365–9. Бибкод : 1969PNAS...63.1365W . дои : 10.1073/pnas.63.4.1365 . ПМК 223473 . ПМИД 5260939 .
- ^ Ян В., Чжэн Ю., Ся Ю., Цзи Х., Чэнь Х., Го Ф. и др. (декабрь 2012 г.). «ERK1/2-зависимое фосфорилирование и ядерная транслокация PKM2 способствуют эффекту Варбурга» . Природная клеточная биология . 14 (12): 1295–304. дои : 10.1038/ncb2629 . ПМК 3511602 . ПМИД 23178880 .
- ^ Ляпунова Н.А., Хампл В., Гордон П.М., Сенсен К.В., Гедаму Л., Дакс Дж.Б. (декабрь 2006 г.). «Реконструкция мозаичного гликолитического пути анаэробных эукариот Monocercomonoides» (Бесплатный полный текст) . Эукариотическая клетка . 5 (12): 2138–46. дои : 10.1128/EC.00258-06 . ПМК 1694820 . ПМИД 17071828 .
Внешние ссылки [ править ]
- Пируват + киназа Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)









