Экзон

Экзон — это любая часть гена , которая образует часть окончательной зрелой РНК , продуцируемой этим геном, после интронов удаления в результате сплайсинга РНК . Термин экзон относится как к последовательности ДНК внутри гена, так и к соответствующей последовательности в транскриптах РНК. При сплайсинге РНК интроны удаляются, а экзоны ковалентно соединяются друг с другом в рамках образования зрелой РНК . Точно так же, как весь набор генов вида составляет геном , весь набор экзонов составляет экзом .
История
[ редактировать ]Термин экзон происходит от экспрессируемой области и был придуман американским биохимиком Уолтером Гилбертом в 1978 году: «Понятие цистрона ... должно быть заменено понятием транскрипционной единицы, содержащей области, которые будут потеряны из зрелого мессенджера – что я предлагаю называть интроны (для внутригенных областей) – чередуясь с областями, которые будут экспрессироваться – экзонами». [1]
Первоначально это определение было дано для транскриптов, кодирующих белки, которые подвергаются сплайсингу перед трансляцией. Позже этот термин стал включать последовательности, удаленные из рРНК. [2] и тРНК , [3] и другие нкРНК [4] позже его также использовали для молекул РНК, происходящих из разных частей генома, которые затем лигировались путем транс-сплайсинга. [5]
Вклад в геномы и распределение по размерам
[ редактировать ]Хотя у одноклеточных эукариот, таких как дрожжи, интроны либо отсутствуют, либо их очень мало, геномы многоклеточных животных и особенно позвоночных содержат большую долю некодирующей ДНК . Например, в геноме человека только 1,1% генома состоит из экзонов, тогда как 24% приходится на интроны, при этом 75% генома представляет собой межгенная ДНК . [6] Это может обеспечить практическое преимущество в омики с использованием здравоохранении (например, в точной медицине ), поскольку делает коммерческое секвенирование всего экзома меньшей и менее дорогостоящей задачей, чем коммерческое секвенирование всего генома . Большие различия в размере генома и значении C у разных форм жизни поставили интересную задачу, получившую название « загадка значения C» .
Во всех эукариотических генах в GenBank насчитывалось (в 2002 г.) в среднем 5,48 экзонов на ген, кодирующий белок. Средний экзон кодирует 30-36 аминокислот . [7] Хотя самый длинный экзон в геноме человека имеет длину 11555 п.н. , было обнаружено, что длина некоторых экзонов составляет всего 2 п.о. [8] Сообщалось об однонуклеотидном экзоне из генома арабидопсиса . [9] У людей, как и белок, кодирующий мРНК , большинство некодирующих РНК также содержат несколько экзонов. [10]
Структура и функции
[ редактировать ]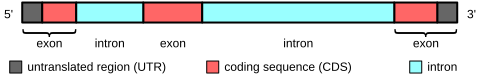
В генах, кодирующих белок, экзоны включают как кодирующую белок последовательность, так и 5'- и 3'- нетранслируемые области (UTR). Часто первый экзон включает в себя как 5'-UTR, так и первую часть кодирующей последовательности, но в некоторых генах встречаются экзоны, содержащие только участки 5'-UTR или (реже) 3'-UTR, т.е. UTR могут содержать интроны. . [11] Некоторые некодирующие транскрипты РНК также имеют экзоны и интроны.
Зрелые мРНК, происходящие из одного и того же гена, не обязательно должны включать одни и те же экзоны, поскольку разные интроны в пре-мРНК могут быть удалены в процессе альтернативного сплайсинга .
Экзонизация — это создание нового экзона в результате мутации интронов . [12]
Экспериментальные подходы с использованием экзонов
[ редактировать ]Захват экзонов или « захват генов » — это метод молекулярной биологии , который использует существование сплайсинга интрон-экзон для поиска новых генов. [13] Первый экзон «захваченного» гена соединяется с экзоном, содержащимся во вставочной ДНК . Этот новый экзон содержит ORF для репортерного гена , который теперь может быть экспрессирован с использованием энхансеров , контролирующих целевой ген. Ученый знает, что новый ген оказывается в ловушке, когда экспрессируется репортерный ген.
Сплайсинг можно экспериментально модифицировать таким образом, чтобы целевые экзоны исключались из зрелых транскриптов мРНК путем блокирования доступа малых ядерных рибонуклеопротеиновых частиц (мяРНП), направляющих сплайсинг, к пре-мРНК с использованием антисмысловых олигонуклеотидов морфолино . [14] Это стало стандартным методом в биологии развития . Морфолиноолиголигоиды также могут быть нацелены на предотвращение связывания молекул, которые регулируют сплайсинг (например, энхансеры сплайсинга, супрессоры сплайсинга), с пре-мРНК, изменяя характер сплайсинга.
Распространенное неправильное использование термина
[ редактировать ]Распространенное неправильное использование термина «экзон» заключается в том, что «экзоны кодируют белок», «экзоны кодируют аминокислоты» или «экзоны транслируются». Однако такого рода определения охватывают только гены, кодирующие белки , и опускают те экзоны, которые становятся частью некодирующей РНК. [15] или область мРНК . нетранслируемая [16] [17] Такие неверные определения до сих пор встречаются во в целом авторитетных вторичных источниках. [18] [19]
См. также
[ редактировать ]- ДБАСС3/5
- Экситрон
- Экзон-интронная база данных
- Перетасовка экзонов
- Прерванный ген
- Аутрон
- Твинтрон
- Непереведенный регион (UTR)
Ссылки
[ редактировать ]- ^ Гилберт В. (февраль 1978 г.). «Почему гены разбиты на кусочки?» . Природа . 271 (5645): 501. Бибкод : 1978Natur.271..501G . дои : 10.1038/271501a0 . ПМИД 622185 .
- ^ Кистер К.П., Эккерт В.А. (март 1987 г.). «Характеристика аутентичного промежуточного продукта в процессе самосплайсинга рибосомальной РНК-предшественника в макроядрах Tetrahymena thermophila» . Исследования нуклеиновых кислот . 15 (5): 1905–20. дои : 10.1093/нар/15.5.1905 . ПМК 340607 . ПМИД 3645543 .
- ^ Валенсуэла П., Венегас А., Вайнберг Ф., Бишоп Р., Раттер В.Дж. (январь 1978 г.). «Структура генов дрожжевой фенилаланин-тРНК: промежуточный сегмент ДНК внутри области, кодирующей тРНК» . Труды Национальной академии наук Соединенных Штатов Америки . 75 (1): 190–4. Бибкод : 1978PNAS...75..190В . дои : 10.1073/pnas.75.1.190 . ПМК 411211 . ПМИД 343104 .
- ^ Хан, MR; Веллингер, Р.Дж.; Лоран, Б. (август 2021 г.). «Изучение альтернативного сплайсинга длинных некодирующих РНК». Тенденции в генетике . 37 (8): 695–698. дои : 10.1016/j.tig.2021.03.010 . ПМИД 33892960 . S2CID 233382870 .
- ^ Лю А.Ю., Ван дер Плог Л.Х., Рейсевийк Ф.А., Борст П. (июнь 1983 г.). «Единица транспозиции вариантного гена поверхностного гликопротеина 118 Trypanosoma brucei. Наличие повторяющихся элементов на ее границе и отсутствие последовательностей, связанных с промотором». Журнал молекулярной биологии . 167 (1): 57–75. дои : 10.1016/S0022-2836(83)80034-5 . ПМИД 6306255 .
- ^ Вентер Дж.К .; и др. (2000). «Последовательность генома человека» . Наука . 291 (5507): 1304–51. Бибкод : 2001Sci...291.1304V . дои : 10.1126/science.1058040 . ПМИД 11181995 .
- ^ Сахаркар М., Пассетти Ф., де Соуза Дж.Э., Лонг М., де Соуза С.Дж. (2002). «ExInt: база данных экзонов-интронов» . Нуклеиновые кислоты Рез . 30 (1): 191–4. дои : 10.1093/нар/30.1.191 . ПМК 99089 . ПМИД 11752290 .
- ^ Сахаркар МК; Чоу В.Т.; Кангеан П. (2004). «Распределение экзонов и интронов в геноме человека». В Силико Биол . 4 (4): 387–93. ПМИД 15217358 .
- ^ Го Лей, Лю Чун-Мин (2015). « Однонуклеотидный экзон, обнаруженный у Arabidopsis» . Научные отчеты . 5 : 18087. Бибкод : 2015NatSR...518087G . дои : 10.1038/srep18087 . ПМЦ 4674806 . ПМИД 26657562 .
- ^ Дерриен, Т; Джонсон, Р.; Буссотти, Дж; Танцер, А; Джебали, С; Тилгнер, Х; Гернек, Г; Мартин, Д; Меркель, А; Ноулз, генеральный директор; Лагард, Дж; Виравалли, Л; Руан, Х; Руан, Ю; Лассманн, Т; Карнинчи, П; Браун, Дж. Б.; Липович, Л; Гонсалес, Дж. М.; Томас, М; Дэвис, Калифорния; Шихаттар, Р; Гингерас, ТР; Хаббард, Ти Джей; Нотредам, К; Харроу, Дж; Гиго, Р. (сентябрь 2012 г.). «Каталог длинных некодирующих РНК человека GENCODE v7: анализ их генной структуры, эволюции и экспрессии» . Геномные исследования . 22 (9): 1775–89. дои : 10.1101/гр.132159.111 . ПМЦ 3431493 . ПМИД 22955988 .
- ^ Бикнелл, А.А. (декабрь 2012 г.). «Интроны в UTR: почему мы должны перестать их игнорировать» . Биоэссе . 34 (12): 1025–1034. doi : 10.1002/bies.201200073 . ПМИД 23108796 . S2CID 5808466 .
- ^ Сорек Р. (октябрь 2007 г.). «Рождение новых экзонов: механизмы и эволюционные последствия» . РНК . 13 (10): 1603–8. дои : 10.1261/rna.682507 . ЧВК 1986822 . ПМИД 17709368 .
- ^ Дуйк Г.М.; Ким СВ; Майерс Р.М.; Кокс Д. Р. (1990). «Захват экзонов: генетический скрининг для идентификации потенциальных транскрибируемых последовательностей в клонированной геномной ДНК млекопитающих» . Труды Национальной академии наук . 87 (22): 8995–8999. Бибкод : 1990PNAS...87.8995D . дои : 10.1073/pnas.87.22.8995 . ПМК 55087 . ПМИД 2247475 .
- ^ Моркос, Пенсильвания (июнь 2007 г.). «Достижение целевого и поддающегося количественной оценке изменения сплайсинга мРНК с олигонуклеотидами морфолино». Связь с биохимическими и биофизическими исследованиями . 358 (2): 521–7. дои : 10.1016/j.bbrc.2007.04.172 . ПМИД 17493584 .
- ^ Хан, MR; Веллингер, Р.Дж.; Лоран, Б. (август 2021 г.). «Изучение альтернативного сплайсинга длинных некодирующих РНК». Тенденции в генетике . 37 (8): 695–698. дои : 10.1016/j.tig.2021.03.010 . ПМИД 33892960 . S2CID 233382870 .
- ^ Лу, Дж; Уильямс, Дж.А.; Люк, Дж; Чжан, Ф; Чу, К; Кей, Массачусетс (январь 2017 г.). «5'-некодирующий экзон, содержащий сконструированный интрон, усиливает экспрессию трансгена из рекомбинантных векторов AAV in vivo» . Генная терапия человека . 28 (1): 125–134. дои : 10.1089/hum.2016.140 . ПМЦ 5278795 . ПМИД 27903072 .
- ^ Чунг, BY; Саймонс, К; Ферт, А.Е.; Браун, СМ; Хелленс, РП (19 мая 2006 г.). «Влияние интронов 5'UTR на экспрессию генов Arabidopsis thaliana» . БМК Геномика . 7 : 120. дои : 10.1186/1471-2164-7-120 . ПМК 1482700 . ПМИД 16712733 .
- ^ «Экзон» . Genome.gov . Архивировано из оригинала 16 марта 2023 г. Проверено 23 марта 2023 г.
- ^ «Экзон» . www.nature.com . Возбудимый. Архивировано из оригинала 23 марта 2023 г. Проверено 23 марта 2023 г.
Библиография
[ редактировать ]- Чжан MQ (май 1998 г.). «Статистические особенности экзонов человека и их фланкирующих областей» . Молекулярная генетика человека . 7 (5): 919–32. дои : 10.1093/hmg/7.5.919 . ПМИД 9536098 .
- Танарадж Т.А., Робинсон А.Дж. (ноябрь 2000 г.). «Предсказание точных границ экзонов» . Краткий. Биоинформ . 1 (4): 343–56. дои : 10.1093/нагрудник/1.4.343 . ПМИД 11465052 .
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|