Мессенджер РНК
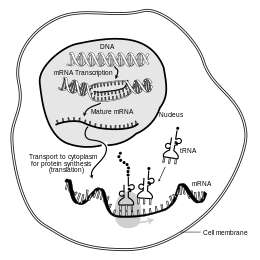
В молекулярной биологии информационная рибонуклеиновая кислота ( мРНК это одноцепочечная молекула РНК в которая соответствует генетической последовательности гена синтеза и считывается рибосомой процессе , белка ) — .
мРНК создается в процессе транскрипции , когда фермент ( РНК-полимераза ) преобразует ген в мРНК первичного транскрипта (также известную как пре-мРНК ). Эта пре-мРНК обычно все еще содержит интроны , участки, которые не будут кодировать окончательную аминокислотную последовательность . Они удаляются в процессе сплайсинга РНК , оставляя только экзоны — области, которые будут кодировать белок. Эта последовательность экзонов составляет зрелую мРНК . Затем зрелая мРНК считывается рибосомой, и рибосома создает белок, используя аминокислоты, переносимые транспортной РНК (тРНК). Этот процесс известен как перевод . Все эти процессы составляют часть центральной догмы молекулярной биологии , которая описывает поток генетической информации в биологической системе.
Как и в ДНК , генетическая информация в мРНК содержится в последовательности нуклеотидов , которые организованы в кодоны, состоящие из трех рибонуклеотидов каждый. Каждый кодон кодирует определенную аминокислоту , за исключением стоп-кодонов , которые завершают синтез белка. Для перевода кодонов в аминокислоты необходимы два других типа РНК: транспортная РНК, которая распознает кодон и обеспечивает соответствующую аминокислоту, и рибосомальная РНК (рРНК), центральный компонент механизма рибосомы по производству белков.
Концепция мРНК была разработана Сиднеем Бреннером и Фрэнсисом Криком в 1960 году во время беседы с Франсуа Жакобом . В 1961 году мРНК была идентифицирована и описана независимо одной командой, состоящей из Бреннера, Джейкоба и Мэтью Мезельсона , и другой командой под руководством Джеймса Уотсона . Анализируя данные при подготовке к публикации, Жакоб и Жак Моно придумали название «РНК-посланник».
Синтез [ править ]
Кратковременное существование молекулы мРНК начинается с транскрипции и в конечном итоге заканчивается деградацией. В течение своей жизни молекула мРНК также может подвергаться процессингу, редактированию и транспортировке перед трансляцией. Молекулы мРНК эукариот часто требуют интенсивного процессинга и транспорта, тогда как молекулы мРНК прокариот этого не делают. Молекула эукариотической мРНК и окружающие ее белки вместе называются информационным РНП . [ нужна ссылка ]
Транскрипция [ править ]
Транскрипция – это копирование РНК с ДНК. Во время транскрипции РНК-полимераза при необходимости копирует ген из ДНК в мРНК. Этот процесс несколько отличается у эукариот и прокариот. Одно заметное отличие состоит в том, что прокариотическая РНК-полимераза связывается с ферментами процессинга ДНК во время транскрипции, так что процессинг может продолжаться во время транскрипции. Таким образом, это приводит к тому, что новая цепь мРНК становится двухцепочечной за счет образования комплементарной цепи, известной как цепь тРНК, которые при объединении не могут образовывать структуры из спаривания оснований. Более того, матрицей для мРНК является комплементарная цепь тРНК, последовательность которой идентична последовательности антикодона, с которым связывается ДНК. Короткоживущий, необработанный или частично обработанный продукт называется мРНК-предшественником или пре-мРНК ; после полной обработки она называется зрелой мРНК . [ нужна ссылка ]
тимина урацилом Замена
мРНК использует в ДНК урацил (U) вместо тимина (Т). урацил (U) является комплементарным основанием аденину (А) во время транскрипции вместо тимина (Т). Поэтому при использовании шаблонной цепи ДНК для построения РНК тимин заменяется урацилом. Эта замена позволяет мРНК переносить соответствующую генетическую информацию от ДНК к рибосоме для трансляции. Что касается естественной истории, урацил был первым, чем тимин; данные свидетельствуют о том, что в эволюции РНК появилась раньше ДНК. [1] Гипотеза мира РНК предполагает, что жизнь началась с молекул РНК до появления геномов ДНК и кодируемых белков. В ДНК эволюционная замена урацила на тимин могла повысить стабильность ДНК и повысить эффективность репликации ДНК. [2] [3]
пре- эукариотической мРНК Процессинг

Процессинг мРНК сильно различается у эукариот , бактерий и архей . Неэукариотическая мРНК, по сути, созревает после транскрипции и не требует процессинга, за исключением редких случаев. [4] Однако эукариотическая пре-мРНК требует нескольких этапов обработки перед транспортировкой в цитоплазму и трансляцией рибосомой.
Сращивание [ править ]
Обширный процессинг эукариотической пре-мРНК, который приводит к образованию зрелой мРНК, представляет собой сплайсинг РНК , механизм, с помощью которого интроны или аутроны (некодирующие области) удаляются и экзоны (кодирующие области) соединяются. [ нужна ссылка ]
Добавление 5-футовой крышки [ править ]
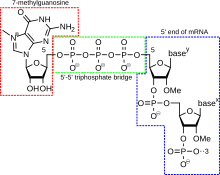
( 5'-кэп также называемый кэпом РНК, кэпом 7-метилгуанозина РНК или мк РНК). 7 G cap) представляет собой модифицированный гуаниновый нуклеотид, который был добавлен к «переднему» или 5'-концу эукариотической информационной РНК вскоре после начала транскрипции. 5'-кэп состоит из терминального остатка 7-метилгуанозина, который связан посредством 5'-5'-трифосфатной связи с первым транскрибируемым нуклеотидом. Его присутствие имеет решающее значение для распознавания рибосомой и защиты от РНКаз . [ нужна ссылка ]
Добавление кэпа связано с транскрипцией и происходит котранскрипционно, так что каждый из них влияет на другой. Вскоре после начала транскрипции 5'-конец синтезируемой мРНК связывается кэп-синтезирующим комплексом, связанным с РНК-полимеразой . Этот ферментативный комплекс катализирует химические реакции, необходимые для кэпирования мРНК. Синтез протекает как многостадийная биохимическая реакция. [ нужна ссылка ]
Редактирование [ править ]
В некоторых случаях мРНК будет редактироваться , изменяя нуклеотидный состав этой мРНК. Примером у человека является мРНК аполипопротеина B , которая редактируется в некоторых тканях, но не в других. В результате редактирования создается ранний стоп-кодон, который при трансляции образует более короткий белок.
Полиаденилирование [ править ]

Полиаденилирование – это ковалентная связь полиаденилильной группы с молекулой информационной РНК. В эукариотических организмах большинство молекул информационной РНК (мРНК) полиаденилированы на 3'-конце, но недавние исследования показали, что также распространены короткие участки уридина (олигоуридилирование). [5] Поли (А)-хвост и связанный с ним белок помогают защитить мРНК от деградации экзонуклеазами. Полиаденилирование также важно для терминации транскрипции, экспорта мРНК из ядра и трансляции. мРНК также может быть полиаденилирована в прокариотических организмах, где поли(А)-хвосты способствуют, а не препятствуют экзонуклеолитической деградации. [ нужна ссылка ]
Полиаденилирование происходит во время и/или сразу после транскрипции ДНК в РНК. После завершения транскрипции цепь мРНК расщепляется под действием эндонуклеазного комплекса, связанного с РНК-полимеразой. После расщепления мРНК к свободному 3'-концу в сайте расщепления добавляются около 250 остатков аденозина. Эту реакцию катализирует полиаденилат-полимераза . Как и при альтернативном сплайсинге , мРНК может иметь более одного варианта полиаденилирования.
Также происходят мутации сайта полиаденилирования. Первичный транскрипт РНК гена расщепляется в сайте добавления поли-А, и к 3'-концу РНК добавляются 100–200 А. Если этот сайт изменить, будет сформирована аномально длинная и нестабильная конструкция мРНК.
Транспорт [ править ]
Еще одно различие между эукариотами и прокариотами — транспорт мРНК. Поскольку транскрипция и трансляция эукариот разделены по компартментам, эукариотические мРНК должны экспортироваться из ядра в цитоплазму — процесс, который может регулироваться различными сигнальными путями. [6] Зрелые мРНК распознаются по их процессированным модификациям и затем экспортируются через ядерную пору путем связывания с кэп-связывающими белками CBP20 и CBP80. [7] а также комплекс транскрипции/экспорта (TREX). [8] [9] У эукариот идентифицировано множество путей экспорта мРНК. [10]
В пространственно сложных клетках некоторые мРНК транспортируются в определенные субклеточные места назначения. В зрелых нейронах определенные мРНК транспортируются из сомы в дендриты . Одним из сайтов трансляции мРНК являются полирибосомы, избирательно локализованные под синапсами. [11] мРНК Arc/Arg3.1 индуцируется синаптической активностью и избирательно локализуется вблизи активных синапсов на основе сигналов, генерируемых NMDA-рецепторами . [12] Другие мРНК также перемещаются в дендриты в ответ на внешние стимулы, например мРНК β-актина . [13] Для экспорта из ядра мРНК актина связывается с ZBP1. [14] и позже с субъединицей 40S . Комплекс связывается с моторным белком и транспортируется к месту-мишени ( отростку нейрита ) по цитоскелету . В конце концов ZBP1 фосфорилируется с помощью Src , чтобы инициировать трансляцию. [15] В развивающихся нейронах мРНК также транспортируются в растущие аксоны и особенно в конусы роста. Многие мРНК помечены так называемыми «почтовыми индексами», которые обеспечивают их транспортировку в определенное место. [16] [17] мРНК также могут передаваться между клетками млекопитающих через структуры, называемые туннельными нанотрубками . [18] [19]
Перевод [ править ]
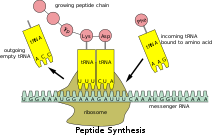
Поскольку прокариотическая мРНК не нуждается в процессинге или транспортировке, трансляция рибосомой может начаться сразу после окончания транскрипции. Следовательно, можно сказать, что прокариотическая трансляция связана с транскрипцией и происходит котранскрипционно . [ нужна ссылка ]
Эукариотическая мРНК, которая была процессирована и транспортирована в цитоплазму (т.е. зрелая мРНК), может затем транслироваться рибосомой. Трансляция может происходить на рибосомах, свободно плавающих в цитоплазме, или направляться в эндоплазматический ретикулум частицей, распознающей сигнал . Следовательно, в отличие от прокариот, эукариотическая трансляция не связана напрямую с транскрипцией. В некоторых случаях даже возможно, что снижение уровней мРНК сопровождается повышением уровней белка, как это наблюдалось для уровней мРНК/белка EEF1A1 при раке молочной железы . [20] [ нужен неосновной источник ]
Структура [ править ]

Кодирование регионов [ править ]
Кодирующие области состоят из кодонов , которые декодируются и транслируются в белки рибосомой; у эукариот обычно на одну, у прокариотов – на несколько. Кодирующие области начинаются со стартового кодона и заканчиваются стоп-кодоном . Как правило, стартовый кодон представляет собой триплет AUG, а стоп-кодон — UAG («янтарный»), UAA («охра») или UGA («опал»). Кодирующие области имеют тенденцию стабилизироваться внутренними парами оснований; это препятствует деградации. [21] [22] Помимо того, что они кодируют белок, части кодирующих областей могут служить регуляторными последовательностями в пре-мРНК в качестве экзонных энхансеров сплайсинга или экзонных сайленсеров сплайсинга .
Непереведенные регионы [ править ]

Нетранслируемые области (UTR) представляют собой участки мРНК перед стартовым кодоном и после стоп-кодона, которые не транслируются, называемые нетранслируемой областью с пятью штрихами (5'-UTR) и нетранслируемой областью с тремя штрихами (3'-UTR) соответственно. Эти области транскрибируются с кодирующей областью и, таким образом, являются экзонными, поскольку они присутствуют в зрелой мРНК. Нетранслируемым областям приписывают несколько ролей в экспрессии генов, включая стабильность мРНК, локализацию мРНК и эффективность трансляции . Способность НТО выполнять эти функции зависит от последовательности НТО и может различаться в зависимости от мРНК. Генетические варианты 3'-UTR также вовлечены в предрасположенность к заболеваниям из-за изменений в структуре РНК и трансляции белков. [23]
Стабильность мРНК можно контролировать с помощью 5'-UTR и/или 3'-UTR благодаря различному сродству к ферментам, разрушающим РНК, называемым рибонуклеазами , и к вспомогательным белкам, которые могут способствовать или ингибировать деградацию РНК. (См. также элемент стабильности, богатый C. )
Эффективность трансляции, включая иногда полное ингибирование трансляции, можно контролировать с помощью UTR. Белки, которые связываются либо с 3'-, либо с 5'-UTR, могут влиять на трансляцию, влияя на способность рибосомы связываться с мРНК. МикроРНК, связанные с 3'-UTR, также могут влиять на эффективность трансляции или стабильность мРНК.
Считается, что цитоплазматическая локализация мРНК является функцией 3'-UTR. Белки, необходимые в определенной области клетки, также могут транслироваться там; в таком случае 3'-UTR может содержать последовательности, которые позволяют локализовать транскрипт в этой области для трансляции.
Некоторые элементы, содержащиеся в нетранслируемых областях, при транскрипции в РНК образуют характерную вторичную структуру . Эти структурные элементы мРНК участвуют в регуляции мРНК. Некоторые из них, такие как элемент SECIS , являются мишенями для связывания белков. Один класс элементов мРНК, рибопереключатели , напрямую связывает небольшие молекулы, изменяя их складку, чтобы изменить уровни транскрипции или трансляции. В этих случаях мРНК регулирует себя.
Поли(А) хвост [ править ]
3'-поли(А)-хвост представляет собой длинную последовательность адениновых нуклеотидов (часто несколько сотен), добавленных к 3'-концу пре-мРНК. Этот хвост способствует экспорту из ядра и трансляции, а также защищает мРНК от деградации.
мРНК полицистронная и Моноцистронная
Молекулу мРНК называют моноцистронной, если она содержит генетическую информацию для трансляции только одной белковой цепи (полипептида). Это справедливо для большинства эукариотических мРНК. [24] [25] С другой стороны, полицистронная мРНК несет несколько открытых рамок считывания (ORF), каждая из которых транслируется в полипептид. Эти полипептиды обычно имеют родственную функцию (они часто представляют собой субъединицы, составляющие конечный комплексный белок), а их кодирующая последовательность группируется и регулируется вместе в регуляторной области, содержащей промотор и оператор . Большая часть мРНК, обнаруженная у бактерий и архей, является полицистронной. [24] как и митохондриальный геном человека. [26] Дицистронная или бицистронная мРНК кодирует только два белка .
циркуляризация мРНК [ править ]

У эукариот молекулы мРНК образуют кольцевые структуры из-за взаимодействия между eIF4E и поли(А)-связывающим белком , которые оба связываются с eIF4G , образуя мост мРНК-белок-мРНК. [27] Считается, что циркуляризация способствует циклическому циклу рибосом на мРНК, что приводит к более быстрой трансляции, а также может обеспечивать трансляцию только интактной мРНК (частично деградированная мРНК обычно не имеет кепки m7G или поли-А-хвоста). [28]
Существуют и другие механизмы циркуляризации, особенно в мРНК вируса. мРНК полиовируса использует участок клеверного листа на своем 5'-конце для связывания PCBP2, который связывает поли(А)-связывающий белок , образуя знакомый круг мРНК-белок-мРНК. Вирус желтой карликовости ячменя связывается между сегментами мРНК на 5'-конце и 3'-конце (так называемые петли целующегося стебля), образуя кольцевую структуру мРНК без участия каких-либо белков.
Геномы РНК-вирусов (+-цепи которых транслируются как мРНК) также обычно имеют кольцевую форму. [29] Во время репликации генома циркуляризация способствует увеличению скорости репликации генома, циклируя вирусную РНК-зависимую РНК-полимеразу почти так же, как предположительно циклизирует рибосома.
Деградация [ править ]
Различные мРНК в одной и той же клетке имеют разное время жизни (стабильность). В бактериальных клетках отдельные мРНК могут сохраняться от нескольких секунд до более часа. Однако время жизни составляет в среднем от 1 до 3 минут, что делает бактериальную мРНК гораздо менее стабильной, чем эукариотическая мРНК. [30] В клетках млекопитающих время жизни мРНК варьируется от нескольких минут до дней. [31] Чем выше стабильность мРНК, тем больше белка может быть произведено из этой мРНК. Ограниченное время жизни мРНК позволяет клетке быстро изменять синтез белка в ответ на меняющиеся потребности. Существует множество механизмов, приводящих к разрушению мРНК, некоторые из которых описаны ниже.
мРНК прокариот Деградация
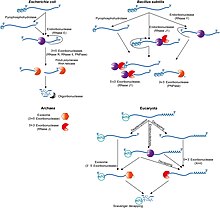
В целом у прокариот время жизни мРНК значительно короче, чем у эукариот. Прокариоты разрушают сообщения, используя комбинацию рибонуклеаз, включая эндонуклеазы , 3'- экзонуклеазы и 5'-экзонуклеазы. В некоторых случаях малые молекулы РНК (мРНК) длиной от десятков до сотен нуклеотидов могут стимулировать деградацию специфических мРНК путем спаривания оснований с комплементарными последовательностями и облегчения расщепления рибонуклеазой РНКазой III . Недавно было показано, что бактерии также имеют своего рода 5'-кэп, состоящий из трифосфата на 5'-конце . [32] Удаление двух фосфатов оставляет 5'-монофосфат, в результате чего сообщение разрушается экзонуклеазой РНКазой J, которая расщепляет 5'-конец до 3'.
Обмен мРНК эукариот [ править ]
Внутри эукариотических клеток существует баланс между процессами трансляции и распада мРНК. Сообщения, которые активно транслируются, связываются рибосомами , факторами инициации эукариот eIF-4E и eIF-4G , а также поли(А)-связывающим белком . eIF-4E и eIF-4G блокируют фермент декапирования ( DCP2 ), а поли(А)-связывающий белок блокирует экзосомный комплекс , защищая концы сообщения. Баланс между трансляцией и распадом отражается в размере и количестве цитоплазматических структур, известных как Р-тельца . [33] Поли (А)-хвост мРНК укорачивается специализированными экзонуклеазами, которые нацелены на специфические информационные РНК с помощью комбинации цис-регуляторных последовательностей на РНК и транс-действующих РНК-связывающих белков. Считается, что удаление хвоста поли(А) нарушает круговую структуру сообщения и дестабилизирует комплекс связывания кэпа . Затем сообщение подвергается деградации либо экзосомным комплексом , либо декапирующим комплексом . Таким образом, трансляционно неактивные сообщения могут быть быстро уничтожены, в то время как активные сообщения остаются нетронутыми. Механизм остановки трансляции и передачи сообщения комплексам распада детально не изучен.
Распад элементов, богатых AU [ править ]
Присутствие богатых AU элементов в мРНК некоторых млекопитающих имеет тенденцию дестабилизировать эти транскрипты за счет действия клеточных белков, которые связывают эти последовательности и стимулируют удаление поли(А) -хвоста. Считается, что потеря поли(А)-хвоста способствует деградации мРНК, облегчая атаку как экзосомного комплекса, так и [34] и декапирующий комплекс . [35] Быстрая деградация мРНК с помощью элементов, богатых AU, является критическим механизмом предотвращения перепроизводства мощных цитокинов, таких как фактор некроза опухоли (TNF) и фактор, стимулирующий колонии гранулоцитов-макрофагов (GM-CSF). [36] Богатые AU элементы также регулируют биосинтез протоонкогенных факторов транскрипции, таких как c-Jun и c-Fos . [37]
Бессмысленный распад
Эукариотические сообщения подлежат наблюдению с помощью нонсенс-опосредованного распада (NMD), который проверяет наличие в сообщении преждевременных стоп-кодонов (нонсенс-кодонов). Они могут возникнуть из-за неполного сплайсинга, рекомбинации V(D)J в адаптивной иммунной системе , мутаций в ДНК, ошибок транскрипции, негерметичного сканирования рибосомой, вызывающего сдвиг рамки считывания , и других причин. Обнаружение преждевременного стоп-кодона запускает деградацию мРНК путем декэпирования 5', удаления 3'- поли(А) -хвоста или эндонуклеолитического расщепления . [38]
РНК ( миРНК Малая интерферирующая )
У животных многоклеточных малые интерферирующие РНК (siRNA), процессируемые Dicer, включаются в комплекс, известный как РНК-индуцированный комплекс молчания или RISC. Этот комплекс содержит эндонуклеазу , которая расщепляет совершенно комплементарные сообщения, с которыми связывается миРНК. Полученные фрагменты мРНК затем разрушаются экзонуклеазами . миРНК обычно используется в лабораториях для блокирования функции генов в клеточной культуре. Считается, что он является частью врожденной иммунной системы и защищает от вирусов с двухцепочечной РНК. [39]
МикроРНК (миРНК) [ править ]
МикроРНК (миРНК) представляют собой небольшие РНК, которые обычно частично комплементарны последовательностям информационных РНК многоклеточных животных. [40] [41] Связывание микроРНК с сообщением может подавлять трансляцию этого сообщения и ускорять удаление поли(А)-хвоста, тем самым ускоряя деградацию мРНК. Механизм действия микроРНК является предметом активных исследований. [42] [43]
механизмы распада Другие
Существуют и другие способы ухудшения качества сообщений, включая , среди прочего, безостановочное затухание и подавление РНК, взаимодействующей с Piwi (piRNA).
Приложения [ править ]
Введение модифицированной нуклеозидами последовательности информационной РНК может заставить клетку вырабатывать белок, который, в свою очередь, может напрямую лечить заболевание или может действовать как вакцина ; более косвенно, белок может заставить эндогенные стволовые клетки дифференцироваться желаемым образом. [44] [45]
Основные задачи РНК-терапии заключаются в доставке РНК в соответствующие клетки. [46] Проблемы включают тот факт, что голые последовательности РНК естественным образом разрушаются после подготовки; организма они могут заставить иммунную систему атаковать их как захватчиков; и они непроницаемы для клеточной мембраны . [45] Оказавшись внутри клетки, они должны покинуть механизм клеточного транспорта, чтобы начать действовать в цитоплазме , в которой находятся необходимые рибосомы . [44]
Чтобы преодолеть эти проблемы, мРНК как терапевтическое средство была впервые предложена в 1989 году «после разработки широко применимой техники трансфекции in vitro». [47] В 1990-х годах были разработаны мРНК-вакцины для персонализированного рака, основанные на ненуклеозидно-модифицированной мРНК. Терапия на основе мРНК продолжает исследоваться как метод лечения или терапии как рака, так и аутоиммунных, метаболических и респираторных воспалительных заболеваний. Терапия редактирования генов, такая как CRISPR, также может принести пользу от использования мРНК, чтобы побудить клетки вырабатывать желаемый белок Cas . [48]
С 2010-х годов РНК-вакцины и другие РНК-терапевтические средства считаются «новым классом лекарств». [49] Первые вакцины на основе мРНК получили ограниченное разрешение и были распространены по всему миру во время пандемии COVID-19 компаниями Pfizer-BioNTech Covid-19 и компанией Moderna . , например, [50] 2023 года Нобелевская премия по физиологии и медицине была присуждена Каталин Карико и Дрю Вайсману за разработку эффективных мРНК-вакцин против COVID-19. [51] [52] [53]
История [ править ]
Несколько исследований молекулярной биологии, проведенных в 1950-х годах, показали, что РНК играет определенную роль в синтезе белка, но эта роль не была четко понята. Например, в одном из самых ранних отчетов Жак Моно и его команда показали, что синтез РНК необходим для синтеза белка, особенно во время производства фермента β-галактозидазы в бактерии E. coli . [54] Артур Парди также обнаружил подобное накопление РНК в 1954 году . [55] В 1953 году Альфред Херши , Джун Диксон и Марта Чейз описали некую цитозинсодержащую ДНК (указав, что это была РНК), которая быстро исчезла после ее синтеза в E. coli . [56] Оглядываясь назад, можно сказать, что это, возможно, было одно из первых наблюдений существования мРНК, но в то время оно как таковое не было признано. [57]
Идея мРНК была впервые высказана Сиднеем Бреннером и Фрэнсисом Криком 15 апреля 1960 года в Королевском колледже Кембриджа , когда Франсуа Жакоб рассказывал им о недавнем эксперименте, проведенном Артуром Парди , им самим и Моно (так называемый эксперимент PaJaMo, которые не доказали существование мРНК, но предположили возможность ее существования). При поддержке Крика Бреннер и Джейкоб немедленно приступили к проверке этой новой гипотезы и обратились за помощью к Мэтью Мезельсону из Калифорнийского технологического института . Летом 1960 года Бреннер, Джейкоб и Мезельсон провели эксперимент в лаборатории Мезельсона в Калифорнийском технологическом институте, который первым доказал существование мРНК. Той осенью Джейкоб и Моно придумали название «информационная РНК» и разработали первую теоретическую основу для объяснения ее функции. [57]
В феврале 1961 года Джеймс Уотсон сообщил, что его исследовательская группа из Гарварда последовала за ними, проведя серию экспериментов, результаты которых указывали примерно в том же направлении. Бреннер и другие согласились на просьбу Уотсона отложить публикацию результатов своего исследования. В результате статьи Бреннера и Уотсона были опубликованы одновременно в одном и том же выпуске журнала Nature в мае 1961 года, а в том же месяце Джейкоб и Моно опубликовали свои теоретические основы мРНК в Журнале молекулярной биологии . [57]
См. также [ править ]
- Тест удлинения Poly(A)
- GeneCalling — технология профилирования мРНК.
- Миссенс-мРНК
- дисплей мРНК
- надзор за мРНК
- Деградация прокариотической мРНК
- Транскриптом – сумма всех РНК в клетке.
Ссылки [ править ]
- ^ «Информация в ДНК декодируется посредством транскрипции | Изучайте науку в Scitable» . www.nature.com . Проверено 3 мая 2024 г.
- ^ «Мир РНК (статья) | Естественный отбор» . Ханская академия . Проверено 3 мая 2024 г.
- ^ «Присутствие тимина вместо урацила также придает ДНК дополнительную стабильность. Как?» . Топпр Спросите . Проверено 4 мая 2024 г.
- ^ Уотсон Джей Ди (22 февраля 2013 г.). Молекулярная биология гена, 7-е издание . Пирсон Высшее образование США. ISBN 9780321851499 .
- ^ Чой Ю.С., Патена В., Ливитт А.Д., Макманус М.Т. (март 2012 г.). «Распространенное олигоуридилирование 3'-конца РНК у млекопитающих» . РНК . 18 (3): 394–401. дои : 10.1261/rna.029306.111 . ПМЦ 3285928 . ПМИД 22291204 .
- ^ Куарежма А.Дж., Сиверт Р., Никерсон Дж.А. (апрель 2013 г.). «Регуляция экспорта мРНК с помощью пути передачи сигнала киназы PI3/AKT» . Молекулярная биология клетки . 24 (8): 1208–1221. doi : 10.1091/mbc.E12-06-0450 . ПМЦ 3623641 . ПМИД 23427269 .
- ^ Кержковски Д., Кмечак М., Пионтек П., Войташек П., Швейковска-Кулинска З., Ярмоловски А. (сентябрь 2009 г.). «Arabidopsis CBP20 нацеливает кэп-связывающий комплекс на ядро и стабилизируется CBP80» . Заводской журнал . 59 (5): 814–825. дои : 10.1111/j.1365-313X.2009.03915.x . ПМИД 19453442 .
- ^ Штрассер К., Масуда С., Мейсон П., Пфаннстил Дж., Оппицци М., Родригес-Наварро С., Рондон А.Г., Агилера А., Струл К., Рид Р., Хёрт Э. (май 2002 г.). «TREX представляет собой консервативный комплекс, сочетающий транскрипцию с экспортом информационной РНК». Природа . 417 (6886): 304–308. Бибкод : 2002Natur.417..304S . дои : 10.1038/nature746 . ПМИД 11979277 . S2CID 1112194 .
- ^ Катахира Дж., Йонеда Ю. (27 октября 2014 г.). «Роль комплекса TREX в ядерном экспорте мРНК» . Биология РНК . 6 (2): 149–152. дои : 10.4161/rna.6.2.8046 . ПМИД 19229134 .
- ^ Ченик С., Чуа Х.Н., Чжан Х., Тарнавски С.П., Акеф А., Дерти А., Тасан М., Мур М.Дж., Палаццо А.Ф., Рот Ф.П. (апрель 2011 г.). «Анализ генома показывает взаимодействие между интронами 5'UTR и экспортом ядерной мРНК для секреторных и митохондриальных генов» . ПЛОС Генетика . 7 (4): e1001366. дои : 10.1371/journal.pgen.1001366 . ПМК 3077370 . ПМИД 21533221 .
- ^ Стюард О, Леви ВБ (март 1982 г.). «Преимущественная локализация полирибосом под основанием дендритных шипиков в гранулярных клетках зубчатой извилины» . Журнал неврологии . 2 (3): 284–291. doi : 10.1523/JNEUROSCI.02-03-00284.1982 . ПМК 6564334 . ПМИД 7062109 .
- ^ Стюард О, Уорли ПФ (апрель 2001 г.). «Избирательное нацеливание вновь синтезированной мРНК Arc на активные синапсы требует активации рецептора NMDA» . Нейрон . 30 (1): 227–240. дои : 10.1016/s0896-6273(01)00275-6 . ПМИД 11343657 . S2CID 13395819 .
- ^ Джоб С., Эбервин Дж. (декабрь 2001 г.). «Локализация и трансляция мРНК в дендритах и аксонах». Обзоры природы. Нейронаука . 2 (12): 889–898. дои : 10.1038/35104069 . ПМИД 11733796 . S2CID 5275219 .
- ^ Олейников Ю., Певица Р.Х. (февраль 2003 г.). «Визуализация в реальном времени ассоциации ZBP1 с мРНК бета-актина во время транскрипции и локализации» . Современная биология . 13 (3): 199–207. Бибкод : 2003CBio...13..199O . дои : 10.1016/s0960-9822(03)00044-7 . ПМЦ 4765734 . ПМИД 12573215 .
- ^ Хюттельмайер С., Зенклузен Д., Ледерер М., Диктенберг Дж., Лоренц М., Менг Х. и др. (ноябрь 2005 г.). «Пространственная регуляция трансляции бета-актина путем Src-зависимого фосфорилирования ZBP1». Природа . 438 (7067): 512–515. Бибкод : 2005Natur.438..512H . дои : 10.1038/nature04115 . ПМИД 16306994 . S2CID 2453397 .
- ^ Олейников Ю., Певица Р.Х. (октябрь 1998 г.). «Локализация РНК: разные почтовые индексы, один и тот же почтальон?» . Тенденции в клеточной биологии . 8 (10): 381–383. дои : 10.1016/s0962-8924(98)01348-8 . ПМК 2136761 . ПМИД 9789325 .
- ^ Эйнгер К., Авосса Д., Диана А.С., Барри С., Барбарезе Э., Карсон Дж.Х. (сентябрь 1997 г.). «Элементы транспорта и локализации в мРНК основного белка миелина» . Журнал клеточной биологии . 138 (5): 1077–1087. дои : 10.1083/jcb.138.5.1077 . ПМК 2136761 . ПМИД 9281585 .
- ^ Хаймович Г., Экер С.М., Дунагин М.К., Эгган Э., Радж А., Герст Дж.Э., Сингер Р.Х. (ноябрь 2017 г.). «Межклеточный транспорт мРНК через расширения, подобные мембранным нанотрубкам, в клетках млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (46): E9873–E9882. Бибкод : 2017PNAS..114E9873H . дои : 10.1073/pnas.1706365114 . ПМК 5699038 . ПМИД 29078295 .
- ^ Хаймович Г., Дасгупта С., Герст Дж.Э. (февраль 2021 г.). «Перенос РНК через туннельные нанотрубки» . Труды Биохимического общества . 49 (1): 145–160. дои : 10.1042/BST20200113 . ПМИД 33367488 . S2CID 229689880 .
- ^ Лин Сай, Битти А, Барадаран Б, Дрей Э, Дуйф ПХ (сентябрь 2018 г.). «Противоположная неправильная экспрессия мРНК и белка EEF1A1 при протоковой карциноме молочной железы из-за регуляции клеточного цикла и клеточного стресса» . Научные отчеты . 8 (1): 13904. Бибкод : 2018NatSR...813904L . дои : 10.1038/s41598-018-32272-x . ПМК 6141510 . ПМИД 30224719 .
- ^ Шабалина С.А., Огурцов А.Ю., Спиридонов Н.А. (2006). «Периодический образец вторичной структуры мРНК, созданный генетическим кодом» . Исследования нуклеиновых кислот . 34 (8): 2428–2437. дои : 10.1093/nar/gkl287 . ПМЦ 1458515 . ПМИД 16682450 .
- ^ Кац Л., Бердж CB (сентябрь 2003 г.). «Широкий отбор локальной вторичной структуры РНК в кодирующих областях бактериальных генов» . Геномные исследования . 13 (9): 2042–2051. дои : 10.1101/гр.1257503 . ПМК 403678 . ПМИД 12952875 .
- ^ Лу Ю.Ф., Могер Д.М., Гольдштейн Д.Б., Урбан Т.Дж., Уикс К.М., Брэдрик С.С. (ноябрь 2015 г.). «Структура мРНК IFNL3 ремоделируется за счет функционального некодирующего полиморфизма, связанного с клиренсом вируса гепатита С» . Научные отчеты . 5 : 16037. Бибкод : 2015NatSR...516037L . дои : 10.1038/srep16037 . ПМК 4631997 . ПМИД 26531896 .
- ^ Jump up to: Перейти обратно: а б Козак М. (март 1983 г.). «Сравнение инициации синтеза белка у прокариот, эукариот и органелл» . Микробиологические обзоры . 47 (1): 1–45. дои : 10.1128/ММБР.47.1.1-45.1983 . ПМК 281560 . ПМИД 6343825 .
- ^ Ниерс К., Поллет Н. (декабрь 1999 г.). «Группы синеэкспрессии у эукариот». Природа . 402 (6761): 483–487. Бибкод : 1999Natur.402..483N . дои : 10.1038/990025 . ПМИД 10591207 . S2CID 4349134 .
- ^ Мерсер Т.Р., Неф С., Динджер М.Е., Кроуфорд Дж., Смит М.А., Ширвуд А.М., Хауген Э., Бракен С.П., Рэкхэм О., Стаматояннопулос Дж.А. , Филиповска А., Мэттик Дж.С. (август 2011 г.). «Митохондриальный транскриптом человека» . Клетка . 146 (4): 645–658. дои : 10.1016/j.cell.2011.06.051 . ПМК 3160626 . ПМИД 21854988 .
- ^ Wells SE, Hillner PE, Vale RD, Sachs AB (июль 1998 г.). «Циркуляризация мРНК с помощью эукариотических факторов инициации трансляции». Молекулярная клетка . 2 (1): 135–140. CiteSeerX 10.1.1.320.5704 . дои : 10.1016/S1097-2765(00)80122-7 . ПМИД 9702200 .
- ^ Лопес-Ластра М., Ривас А., Барриа М.И. (2005). «Синтез белка у эукариот: растущая биологическая значимость инициации трансляции, независимой от кэпа» . Биологические исследования . 38 (2–3): 121–146. дои : 10.4067/S0716-97602005000200003 . ПМИД 16238092 .
- ^ Чжан X, Лян Z, Ван C, Шэнь Z, Сунь С, Гун C, Ху X (2022). «Вирусные кольцевые РНК и их возможная роль во взаимодействии вирус-хозяин» . Границы в иммунологии . 13 : 939768. дои : 10.3389/fimmu.2022.939768 . ПМЦ 9247149 . PMID 35784275 .
- ^ Левин Б. , Кребс Дж.Э., Килпатрик С.Т., Гольдштейн Э.С., ред. (2011). Гены Левина X (10-е изд.). Садбери, Массачусетс: Джонс и Бартлетт. ISBN 9780763766320 . OCLC 456641931 .
- ^ Ю Дж., Рассел Дж. Э. (сентябрь 2001 г.). «Структурный и функциональный анализ комплекса мРНП, который обеспечивает высокую стабильность мРНК бета-глобина человека» . Молекулярная и клеточная биология . 21 (17): 5879–5888. дои : 10.1128/mcb.21.17.5879-5888.2001 . ПМЦ 87307 . ПМИД 11486027 .
- ^ Дина А., Селесник Х., Беласко Дж.Г. (январь 2008 г.). «Бактериальный фермент RppH запускает деградацию информационной РНК путем удаления 5'-пирофосфата». Природа . 451 (7176): 355–358. Бибкод : 2008Natur.451..355D . дои : 10.1038/nature06475 . ПМИД 18202662 . S2CID 4321451 .
- ^ Паркер Р., Шет У (март 2007 г.). «Р-тельца и контроль трансляции и деградации мРНК» . Молекулярная клетка . 25 (5): 635–646. doi : 10.1016/j.molcel.2007.02.011 . ПМИД 17349952 .
- ^ Чен С.И., Герци Р., Онг С.Э., Чан Э.Л., Райджмейкерс Р., Пруйн Г.Дж., Стеклин Г., Морони С., Манн М., Карин М. (ноябрь 2001 г.). «AU-связывающие белки привлекают экзосому для разрушения ARE-содержащих мРНК» . Клетка . 107 (4): 451–464. дои : 10.1016/S0092-8674(01)00578-5 . ПМИД 11719186 . S2CID 14817671 .
- ^ Фенгер-Грён М., Филлман С., Норрильд Б., Люкке-Андерсен Дж. (декабрь 2005 г.). «Множественные факторы обработки тела и ARE-связывающий белок TTP активируют декапирование мРНК» . Молекулярная клетка . 20 (6): 905–915. doi : 10.1016/j.molcel.2005.10.031 . ПМИД 16364915 .
- ^ Шоу Дж., Камен Р. (август 1986 г.). «Консервативная последовательность AU из 3'-нетранслируемой области мРНК GM-CSF опосредует селективную деградацию мРНК». Клетка . 46 (5): 659–667. дои : 10.1016/0092-8674(86)90341-7 . ПМИД 3488815 . S2CID 40332253 .
- ^ Чен С.И., Шю А.Б. (ноябрь 1995 г.). «Элементы, богатые AU: характеристика и значение в деградации мРНК». Тенденции биохимических наук . 20 (11): 465–470. дои : 10.1016/S0968-0004(00)89102-1 . ПМИД 8578590 .
- ^ Искен О, Макват Л.Е. (август 2007 г.). «Контроль качества эукариотической мРНК: защита клеток от аномальной функции мРНК» . Гены и развитие . 21 (15): 1833–1856. дои : 10.1101/gad.1566807 . ПМИД 17671086 .
- ^ Оббард DJ, Гордон К.Х., Бак А.Х., Джиггинс FM (январь 2009 г.). «Эволюция РНКи как защита от вирусов и мобильных элементов» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 364 (1513): 99–115. дои : 10.1098/rstb.2008.0168 . ПМЦ 2592633 . ПМИД 18926973 .
- ^ Роберт Э. Фаррелл-младший. Методологии РНК, 5-е издание. Академик Пресс, 2017.
- ^ Бреннеке Дж., Старк А., Рассел Р.Б., Коэн С.М. (март 2005 г.). «Принципы распознавания микроРНК-мишеней» . ПЛОС Биология . 3 (3): е85. doi : 10.1371/journal.pbio.0030085 . ПМЦ 1043860 . ПМИД 15723116 .
- ^ Тасуку Хондзё, Майкл Рет, Андреас Радбрух, Фредерик Альт. Молекулярная биология B-клеток, 2-е издание. Academic Press, 2014 г. (включая «обновленные исследования микроРНК»)
- ^ Эулалио А., Ханцингер Э., Нишихара Т., Ревинкель Дж., Фаузер М., Изаурральде Э. (январь 2009 г.). «Деаденилирование является широко распространенным эффектом регуляции микроРНК» . РНК . 15 (1): 21–32. дои : 10.1261/rna.1399509 . ПМЦ 2612776 . ПМИД 19029310 .
- ^ Jump up to: Перейти обратно: а б Хадж К.А., Уайтхед К.А. (12 сентября 2017 г.). «Инструменты трансляции: невирусные материалы для доставки терапевтической мРНК» . Материалы обзоров природы . 2 (10): 17056. Бибкод : 2017NatRM...217056H . дои : 10.1038/natrevmats.2017.56 .
- ^ Jump up to: Перейти обратно: а б Гусейнов Э., Козлов М., Скэнлан С. (15 сентября 2015 г.). «Терапия и вакцины на основе РНК» . Новости генной инженерии .
- ^ Качмарек Ю.К., Ковальски П.С., Андерсон Д.Г. (июнь 2017 г.). «Достижения в области РНК-терапии: от концепции к клинической реальности» . Геномная медицина . 9 (1): 60. дои : 10.1186/s13073-017-0450-0 . ПМЦ 5485616 . ПМИД 28655327 .
- ^ Шлейк Т., Тесс А., Фотин-Млечек М., Каллен К.Дж. (ноябрь 2012 г.). «Разработка технологий мРНК-вакцины» . Биология РНК . 9 (11): 1319–30. дои : 10.4161/rna.22269 . ПМЦ 3597572 . ПМИД 23064118 .
- ^ Хариди Р. (23 апреля 2021 г.). «Революция мРНК: как COVID-19 ускорил развитие экспериментальной технологии» . Новый Атлас . Проверено 26 апреля 2021 г.
- ^ Ковальска Дж, Выпиевска дель Ногаль А, Даржинкевич ЗМ, Бак Дж, Никола С, Кун А.Н., Лукашевич М, Зуберек Дж, Стренковска М, Земняк М, Мацейчик М, Боярска Е, Роудс Р.Э., Даржинкевич Е, Шахин У, Джемилити Дж ( 2014), «Терапия на основе мРНК – разработка нового класса лекарств», Nature Reviews Drug Discovery , vol 13, no. 10, с. 759–780, doi : 10.1093/nar/gku757 , PMC 4176373 , PMID 25150148
- ^ Барбье А.Дж., Цзян А.Ю., Чжан П., Вустер Р., Андерсон Д.Г. (июнь 2022 г.). «Клинический прогресс мРНК-вакцин и иммунотерапии» . Природная биотехнология . 40 (6): 840–854. дои : 10.1038/s41587-022-01294-2 . ПМИД 35534554 . S2CID 248667843 .
- ^ «Нобелевская премия по физиологии и медицине 2023 года» . NobelPrize.org . Проверено 3 октября 2023 г.
- ^ «Венгерские и американские учёные получили Нобелевскую премию за открытие вакцины против COVID-19» . Рейтер . 2023-10-02 . Проверено 3 октября 2023 г.
- ^ «Нобелевская премия по физиологии и медицине 2023 года» . NobelPrize.org . Проверено 3 октября 2023 г.
- ^ Моно Дж., Паппенхаймер А.М., Коэн-Базир Г. (1952). «Кинетика биосинтеза β-галактозидазы в E. coli в зависимости от роста». Biochimica et Biophysical Acta (на французском языке). 9 (6): 648–660. дои : 10.1016/0006-3002(52)90227-8 . ПМИД 13032175 .
- ^ Парди AB (май 1954 г.). «Предшественники нуклеиновых кислот и синтез белка» . Труды Национальной академии наук Соединенных Штатов Америки . 40 (5): 263–270. Бибкод : 1954PNAS...40..263P . дои : 10.1073/pnas.40.5.263 . ПМК 534118 . ПМИД 16589470 .
- ^ Херши А.Д., Диксон Дж., Чейз М. (июль 1953 г.). «Экономика нуклеиновых кислот у бактерий, инфицированных бактериофагом Т2. I. Пуриновый и пиримидиновый состав» . Журнал общей физиологии . 36 (6): 777–789. дои : 10.1085/jgp.36.6.777 . ПМК 2147416 . ПМИД 13069681 .
- ^ Jump up to: Перейти обратно: а б с Кобб М. (29 июня 2015 г.). «Кто открыл информационную РНК?» . Современная биология . 25 (13): Р526–Р532. Бибкод : 2015CBio...25.R526C . дои : 10.1016/j.cub.2015.05.032 . ПМИД 26126273 .
Дальнейшее чтение [ править ]
- Алсавид М., Лай К.Т., Хартманн П.Е., Геддес Д.Т., Какулас Ф. (февраль 2016 г.). «МиРНК человеческого молока в основном происходят из молочной железы, что приводит к уникальным профилям микроРНК фракционированного молока» . Научные отчеты . 6 (1): 20680. Бибкод : 2016NatSR...620680A . дои : 10.1038/srep20680 . ПМЦ 4745068 . ПМИД 26854194 .
- Лилликроп К.А. , Бердж Г.К. (октябрь 2012 г.). «Эпигенетические механизмы, связывающие раннее питание с долгосрочным здоровьем». Лучшие практики и исследования. Клиническая эндокринология и обмен веществ . 26 (5): 667–676. дои : 10.1016/j.beem.2012.03.009 . ПМИД 22980048 .
- Мельник Б.К., Какулас Ф., Геддес Д.Т., Хартманн П.Е., Джон С.М., Каррера-Бастос П., Корден Л., Шмитц Г. (21 июня 2016 г.). «МиРНК молока: простые питательные вещества или системные функциональные регуляторы?» . Питание и обмен веществ . 13 (1): 42. дои : 10.1186/s12986-016-0101-2 . ПМЦ 4915038 . ПМИД 27330539 .
- Викерс М.Х. (июнь 2014 г.). «Питание в раннем возрасте, эпигенетика и программирование болезней в более позднем возрасте» . Питательные вещества . 6 (6): 2165–2178. дои : 10.3390/nu6062165 . ПМК 4073141 . ПМИД 24892374 .
- Чжоу Q, Ли М, Ван X, Ли Q, Ван Т, Чжу Q, Чжоу X, Ван X, Гао X, Ли X (2012). «МикроРНК, связанные с иммунитетом, в изобилии содержатся в экзосомах грудного молока» . Международный журнал биологических наук . 8 (1): 118–123. дои : 10.7150/ijbs.8.118 . ПМЦ 3248653 . ПМИД 22211110 .
- Краузе В. (2023). «МРНК — от лечения COVID-19 до иммунотерапии рака» . Биомедицины . 11 (2): 308. doi : 10.3390/biomedicines11020308 . ПМЦ 9953480 . ПМИД 36830845 .
Внешние ссылки [ править ]
- Атлас RNAi : база данных библиотек RNAi и результатов их целевого анализа.
- miRSearch. Архивировано 4 декабря 2012 г. в Wayback Machine : Инструмент для поиска микроРНК, нацеленных на мРНК.
- Как кодируется мРНК? : видео на YouTube
- Что такое мРНК? : theconversation.com
Виды нуклеиновых кислот |
|---|