Сдвиг рамки рибосом
рибосомы Сдвиг рамки , также известный как сдвиг рамки трансляции или трансляционное перекодирование , представляет собой биологическое явление, возникающее во время трансляции и приводящее к образованию множества уникальных белков из одной мРНК . [ 1 ] Этот процесс может быть запрограммирован нуклеотидной последовательностью мРНК, а иногда на него влияет вторичная трехмерная структура мРНК . [ 2 ] Он описан главным образом у вирусов (особенно ретровирусов ), ретротранспозонов и бактериальных инсерционных элементов, а также в некоторых клеточных генах . [ 3 ]
Также было обнаружено, что небольшие молекулы, белки и нуклеиновые кислоты стимулируют сдвиг рамки считывания. В декабре 2023 года сообщалось, что in vitro транскрибируемые (IVT) мРНК в ответ на вакцину BNT162b2 (Pfizer-BioNTech) против COVID-19 вызвали сдвиг рамки рибосомы. [ 4 ]
Обзор процесса
[ редактировать ]Белки транслируются путем считывания тринуклеотидов на цепи мРНК, также известных как кодоны , от одного конца мРНК к другому (от 5'- конца к 3'-концу ), начиная с аминокислоты метионина в качестве начала (инициации). кодон AUG. Каждый кодон транслируется в одну аминокислоту . Сам код считается вырожденным , то есть определенная аминокислота может обозначаться более чем одним кодоном. Однако сдвиг любого количества нуклеотидов, не кратного 3, в рамке считывания приведет к тому, что последующие кодоны будут читаться по-другому. [ 5 ] рибосом Это эффективно изменяет рамку считывания .
Пример предложения
[ редактировать ]В этом примере следующее предложение, состоящее из трехбуквенных слов, имеет смысл, если читать его с начала:
|Start|THE CAT AND THE MAN ARE FAT ... |Start|123 123 123 123 123 123 123 ...
Однако, если рамка считывания смещается на одну букву между T и H первого слова (фактически сдвиг кадра на +1, если считать, что позиция 0 является начальной позицией T ),
T|Start|HEC ATA NDT HEM ANA REF AT... -|Start|123 123 123 123 123 123 12...
тогда предложение читается по-другому и теряет смысл.
пример ДНК
[ редактировать ]В этом примере следующая последовательность представляет собой область митохондриального генома человека с двумя перекрывающимися генами MT-ATP8 и MT-ATP6 . При чтении с самого начала эти кодоны имеют смысл для рибосомы и могут транслироваться в аминокислоты (АА) под митохондриальным кодом позвоночных :
|Start|AAC GAA AAT CTG TTC GCT TCA ... |Start|123 123 123 123 123 123 123 ... | AA | N E N L F A S ...
Однако давайте изменим рамку считывания, начав на один нуклеотид ниже (фактически «сдвиг рамки на +1», если считать позицию 0 начальной позицией A ):
A|Start|ACG AAA ATC TGT TCG CTT CA... -|Start|123 123 123 123 123 123 12... | AA | T K I C S L ...
Из-за этого сдвига рамки +1 последовательность ДНК читается по-другому. Таким образом, разные рамки считывания кодонов дают разные аминокислоты.
Эффект
[ редактировать ]В случае транслирующей рибосомы сдвиг рамки считывания может привести либо к бессмысленной мутации , преждевременному появлению стоп-кодона после сдвига рамки считывания, либо к созданию совершенно нового белка после сдвига рамки считывания. В случае, когда сдвиг рамки приводит к нонсенсу, путь нонсенс-опосредованного распада мРНК (NMD) может разрушить транскрипт мРНК, поэтому сдвиг рамки может служить методом регулирования уровня экспрессии соответствующего гена. [ 6 ]
Если вырабатывается новый или нецелевой белок, это может вызвать другие неизвестные последствия. [ 4 ]
Функция у вирусов и эукариот
[ редактировать ]У вирусов это явление может быть запрограммировано на возникновение определенных участков и позволяет вирусу кодировать несколько типов белков одной и той же мРНК. Яркие примеры включают ВИЧ-1 (вирус иммунодефицита человека), [ 7 ] РСВ ( вирус саркомы Рауса ) [ 8 ] и вирус гриппа (грипп), [ 9 ] все они основаны на сдвиге рамки считывания для создания правильного соотношения белков 0-кадра (нормальная трансляция) и «транс-фрейма» (кодируемого последовательностью со сдвигом рамки считывания). Его использование в вирусах в первую очередь предназначено для уплотнения большего количества генетической информации в более короткое количество генетического материала.
У эукариот он, по-видимому, играет роль в регуляции уровня экспрессии генов, вызывая преждевременную остановку и производя нефункциональные транскрипты. [ 3 ] [ 10 ]
Типы смены кадров
[ редактировать ]Наиболее распространенным типом сдвига кадра является -1 сдвиг кадра или запрограммированный -1 рибосомальный сдвиг кадра (-1 PRF) . Другие, более редкие типы сдвига кадра включают сдвиг кадра +1 и -2. [ 2 ] Считается, что сдвиг кадров -1 и +1 управляется разными механизмами, которые обсуждаются ниже. Оба механизма имеют кинетический привод .
Запрограммированный сдвиг рамки -1 рибосомы
[ редактировать ]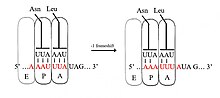
При сдвиге рамки -1 рибосома отодвигает один нуклеотид назад и продолжает трансляцию в кадре -1. Обычно существует три элемента, которые содержат сигнал сдвига рамки -1: скользящая последовательность , спейсерная область и вторичная структура РНК. Скользкая последовательность соответствует мотиву X_XXY_YYH, где XXX — это любые три идентичных нуклеотида (хотя встречаются и некоторые исключения), YYY обычно представляет собой UUU или AAA, а H — это A, C или U. Поскольку структура этого мотива содержит 2 соседних 3-нуклеотида Повторы полагают, что сдвиг рамки -1 описывается моделью тандемного проскальзывания, в которой антикодон тРНК рибосомального P-сайта повторно спаривается с XXY на XXX, а антикодон A-сайта повторно спаривается с YYH на YYY одновременно. Эти новые пары идентичны парам 0-кадров, за исключением их третьих позиций. Это различие незначительно ухудшает связывание антикодонов, поскольку третий нуклеотид в кодоне, известный как положение качания , имеет более слабую специфичность связывания антикодона тРНК, чем первый и второй нуклеотиды. [ 2 ] [ 11 ] В этой модели структура мотива объясняется тем фактом, что первая и вторая позиции антикодонов должны иметь возможность идеально спариваться как в кадрах 0, так и в -1. Следовательно, нуклеотиды 2 и 1 должны быть идентичными, а нуклеотиды 3 и 2 также должны быть идентичными, что приводит к необходимой последовательности из 3 идентичных нуклеотидов для каждой проскальзывающей тРНК. [ 12 ]
+1 сдвиг рамки рибосомы
[ редактировать ]Скользящая последовательность для сигнала сдвига рамки +1 не имеет того же мотива и вместо этого, по-видимому, действует, приостанавливая рибосому на последовательности, кодирующей редкую аминокислоту. [ 13 ] Рибосомы не транслируют белки с постоянной скоростью, независимо от последовательности. неравное количество тРНК этого конкретного кодона Трансляция некоторых кодонов занимает больше времени, поскольку в цитозоле . [ 14 ] Из-за этого отставания на небольших участках существуют последовательности кодонов, которые контролируют скорость сдвига рамки рибосомы. В частности, рибосома должна сделать паузу, чтобы дождаться прибытия редкой тРНК, и это увеличивает кинетическую благоприятность рибосомы и связанной с ней тРНК, проскользнувшей в новую рамку. [ 13 ] [ 15 ] В этой модели изменение рамки считывания вызвано одним проскальзыванием тРНК, а не двумя.
Механизмы управления
[ редактировать ]Сдвиг рамки рибосомы может контролироваться механизмами, обнаруженными в последовательности мРНК (цис-действующими). Обычно это относится к скользкой последовательности, вторичной структуре РНК или к тому и другому. Сигнал сдвига рамки -1 состоит из обоих элементов, разделенных спейсерной областью, обычно длиной 5–9 нуклеотидов. [ 2 ] Сдвиг рамки считывания также может быть индуцирован другими молекулами, которые взаимодействуют с рибосомой или мРНК (транс-действуя).
Элементы сигнала сдвига кадра
[ редактировать ]
Скользкая последовательность
[ редактировать ]Скользкие последовательности потенциально могут заставить считывающую рибосому «скользить» и пропускать определенное количество нуклеотидов (обычно только 1) и после этого читать совершенно другой кадр. При запрограммированном сдвиге рамки рибосомы -1 скользкая последовательность соответствует мотиву X_XXY_YYH, где XXX — любые три идентичных нуклеотида (хотя встречаются и некоторые исключения), YYY обычно представляет собой UUU или AAA, а H представляет собой A, C или U. В случае + 1, скользкая последовательность содержит кодоны, для которых соответствующая тРНК более редка, и сдвиг рамки предпочтителен, поскольку кодон в новом рамка имеет более общую ассоциированную тРНК. [ 13 ] Одним из примеров скользкой последовательности является полиА на мРНК, которая, как известно, индуцирует проскальзывание рибосом даже в отсутствие каких-либо других элементов. [ 16 ]
Вторичная структура РНК
[ редактировать ]Эффективный сдвиг рамки рибосом обычно требует присутствия вторичной структуры РНК для усиления эффекта скользкой последовательности. [ 12 ] Считается , что структура РНК (которая может представлять собой петлю -стебель или псевдоузел ) приостанавливает рибосому на скользком участке во время трансляции, заставляя ее перемещаться и продолжать репликацию из положения -1. Считается, что это происходит потому, что структура физически блокирует движение рибосомы, застревая в туннеле мРНК рибосомы. [ 2 ] Эта модель подтверждается тем фактом, что сила псевдоузла положительно коррелирует с уровнем сдвига рамки считывания ассоциированной мРНК. [ 3 ] [ 17 ]
Ниже приведены примеры предсказанных вторичных структур для элементов сдвига рамки считывания, которые, как показано, стимулируют сдвиг рамки считывания у различных организмов. Большинство показанных структур представляют собой стебель-петли, за исключением структуры псевдоузла ALIL (апикальная петля-внутренняя петля). На этих изображениях большие и неполные кружки мРНК представляют собой линейные области. Вторичные структуры «стебель-петля», где «стебли» образованы участком спаривания оснований мРНК с другим участком на той же цепи, показаны выступающими из линейной ДНК. Линейная область сигнала сдвига рамки рибосомы ВИЧ содержит высококонсервативную скользкую последовательность UUU UUU A; многие из других предсказанных структур также содержат кандидатов на скользкие последовательности.
Последовательности мРНК на изображениях можно прочитать в соответствии с рядом правил. Хотя A, T, C и G обозначают конкретный нуклеотид в определенной позиции, существуют также буквы, обозначающие неоднозначность, которые используются, когда в этой позиции может находиться более одного типа нуклеотидов. Правила Международного союза теоретической и прикладной химии ( IUPAC ) заключаются в следующем: [ 18 ]
| Символ [ 18 ] | Описание | Представленные базы | Дополнить | ||||
|---|---|---|---|---|---|---|---|
| А | Денин | А | 1 | Т | |||
| С | Цитозин | С | Г | ||||
| Г | Гуанин | Г | С | ||||
| Т | Т химин | Т | А | ||||
| В | Ты расовый | В | А | ||||
| В | Слабый | А | Т | 2 | В | ||
| С | Сильный | С | Г | С | |||
| М | Мино | А | С | К | |||
| К | К это | Г | Т | М | |||
| Р | Пу Рин | А | Г | Р | |||
| И | p Y римидин | С | Т | И | |||
| Б | не А ( Б идет после А) | С | Г | Т | 3 | V | |
| Д | не C ( D идет после C) | А | Г | Т | ЧАС | ||
| ЧАС | не G ( H идет после G) | А | С | Т | Д | ||
| V | не T ( V идет после T и U) | А | С | Г | Б | ||
| Н | любой нуклеотид (не пробел) | А | С | Г | Т | 4 | Н |
| С | ноль | 0 | С | ||||
Эти символы также действительны для РНК, за исключением того, что U (урацил) заменяет Т (тимин). [ 18 ]
Галерея изображений вторичной структуры | |
|---|---|
|




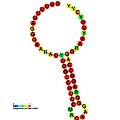


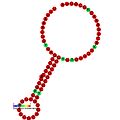




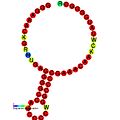

| Тип | Распределение | Ссылка. |
|---|---|---|
| Псевдоузел ALIL | Бактерии | [ 19 ] |
| Элемент стимуляции сдвига рамки антизимной РНК | Беспозвоночные | [ 20 ] |
| Элемент стимуляции сдвига кадра коронавируса | Coronavirus | [ 21 ] |
| Элемент сдвига рамки рибосомы DnaX | Эукариоты , бактерии | [ 22 ] |
| Сигнал сдвига рамки рибосомы ВИЧ | Вирусы | |
| Вставочная последовательность рибосомального элемента сдвига рамки IS1222 | Эукариоты , бактерии | |
| Сдвиг рамки рибосом | Вирусы |
Транзактные элементы
[ редактировать ]Было обнаружено, что небольшие молекулы, белки и нуклеиновые кислоты стимулируют сдвиг рамки считывания. Например, механизм отрицательной обратной связи в пути синтеза полиаминов основан на том, что уровни полиаминов стимулируют увеличение сдвига рамки +1, что приводит к выработке ингибирующего фермента . Было также показано, что некоторые белки, которые необходимы для распознавания кодонов или которые непосредственно связываются с последовательностью мРНК, модулируют уровни сдвига рамки считывания. Молекулы микроРНК (миРНК) могут гибридизоваться со вторичной структурой РНК и влиять на ее прочность. [ 6 ]
См. также
[ редактировать ]- Элемент стимуляции сдвига рамки антизимной РНК
- Элемент стимуляции сдвига кадра коронавируса
- Элемент сдвига рамки рибосомы DnaX
- Мутация сдвига рамки
- Сигнал сдвига рамки рибосомы ВИЧ
- Вставочная последовательность рибосомального элемента сдвига рамки IS1222
- Перекодировать базу данных
- Рибосомальная пауза
- Скользкая последовательность
Ссылки
[ редактировать ]- ^ Аткинс Дж. Ф., Логран Дж., Бхатт П. Р., Ферт А. Е., Баранов П. В. (сентябрь 2016 г.). «Рибосомальный сдвиг рамки и проскальзывание транскрипции: от генетической стеганографии и криптографии к случайному использованию» . Исследования нуклеиновых кислот . 44 (15): 7007–7078. дои : 10.1093/nar/gkw530 . ПМЦ 5009743 . ПМИД 27436286 .
- ^ Перейти обратно: а б с д и Наптин С., Линг Р., Финч Л.К., Джонс Дж.Д., Белл С., Брайерли И., Ферт А.Е. (июнь 2017 г.). «Белковый сдвиг рамки рибосомы регулирует во времени экспрессию генов» . Природные коммуникации . 8 : 15582. Бибкод : 2017NatCo...815582N . дои : 10.1038/ncomms15582 . ПМЦ 5472766 . ПМИД 28593994 .
- ^ Перейти обратно: а б с Кеттелер Р. (2012). «О запрограммированном сдвиге рамки рибосом: альтернативные протеомы» . Границы генетики . 3 : 242. дои : 10.3389/fgene.2012.00242 . ПМК 3500957 . ПМИД 23181069 .
- ^ Перейти обратно: а б Малруни, Томас Э.; Пойри, Туйя; Ям-Пук, Хуан Карлос; Руст, Мария; Харви, Роберт Ф.; Кальмар, Лайош; Хорнер, Эмили; Бут, Люси; Феррейра, Александр П.; Стоунли, Марк; Саваркар, Ритвик; Ментцер, Александр Дж.; Лилли, Кэтрин С.; Смейлс, К. Марк; фон дер Хаар, Тобиас (6 декабря 2023 г.). «N1-метилпсевдоуридилирование мРНК вызывает сдвиг рамки рибосомы на +1» . Природа . 625 (7993): 189–194. дои : 10.1038/s41586-023-06800-3 . ISSN 1476-4687 . ПМЦ 10764286 . ПМИД 38057663 .
- ^ Иванов И.П., Аткинс Дж.Ф. (2007). «Сдвиг рамки рибосом при декодировании антизимных мРНК от дрожжей и простейших к человеку: около 300 случаев демонстрируют значительное разнообразие, несмотря на лежащую в основе консервативность» . Исследования нуклеиновых кислот . 35 (6): 1842–1858. дои : 10.1093/нар/gkm035 . ПМК 1874602 . ПМИД 17332016 .
- ^ Перейти обратно: а б Девер Т.Е., Динман Дж.Д., Грин Р. (август 2018 г.). «Удлинение трансляции и перекодирование у эукариот» . Перспективы Колд-Спринг-Харбор в биологии . 10 (8): а032649. doi : 10.1101/cshperspect.a032649 . ПМК 6071482 . ПМИД 29610120 .
- ^ Перейти обратно: а б Джекс Т., Пауэр МД, Масиарз ФР, Люцив П.А., Барр П.Дж., Вармус Х.Э. (январь 1988 г.). «Характеристика сдвига рамки рибосомы при экспрессии gag-pol ВИЧ-1». Природа . 331 (6153): 280–283. Бибкод : 1988Natur.331..280J . дои : 10.1038/331280a0 . ПМИД 2447506 . S2CID 4242582 .
- ^ Перейти обратно: а б Джекс Т., Мадхани Х.Д., Масиарз Ф.Р., Вармус Х.Э. (ноябрь 1988 г.). «Сигналы для сдвига рамки рибосомы в области gag-pol вируса саркомы Рауса» . Клетка . 55 (3): 447–458. дои : 10.1016/0092-8674(88)90031-1 . ПМЦ 7133365 . ПМИД 2846182 .
- ^ Джаггер Б.В., Уайз Х.М., Каш Дж.К., Уолтерс К.А., Уиллс Н.М., Сяо Ю.Л., Данфи Р.Л., Шварцман Л.М., Озински А., Белл Г.Л., Далтон Р.М., Ло А., Эфстатиу С., Аткинс Дж.Ф., Ферт А.Э., Таубенбергер Дж.К., Дигард П. (июль 2012 г.). «Перекрывающаяся кодирующая белок область в сегменте 3 вируса гриппа А модулирует реакцию хозяина» . Наука . 337 (6091): 199–204. Бибкод : 2012Sci...337..199J . дои : 10.1126/science.1222213 . ПМЦ 3552242 . ПМИД 22745253 .
- ^ Адвани В.М., Динман Дж.Д. (январь 2016 г.). «Перепрограммирование генетического кода: новая роль сдвига рамки рибосом в регуляции клеточной экспрессии генов» . Биоэссе . 38 (1): 21–26. doi : 10.1002/bies.201500131 . ПМЦ 4749135 . ПМИД 26661048 .
- ^ Крик Ф.Х. (август 1966 г.). «Спаривание кодонов и антикодонов: гипотеза колебания». Журнал молекулярной биологии . 19 (2): 548–555. дои : 10.1016/S0022-2836(66)80022-0 . ПМИД 5969078 .
- ^ Перейти обратно: а б Брайерли I (август 1995 г.). «Рибосомальные вирусные РНК со сдвигом рамки» . Журнал общей вирусологии . 76 (Часть 8) (8): 1885–1892. дои : 10.1099/0022-1317-76-8-1885 . ПМИД 7636469 .
- ^ Перейти обратно: а б с д Харгер Дж.В., Мескаускас А., Динман Дж.Д. (сентябрь 2002 г.). «Интегрированная модель» запрограммированного сдвига рамки считывания рибосом» . Тенденции биохимических наук . 27 (9): 448–454. дои : 10.1016/S0968-0004(02)02149-7 . ПМИД 12217519 .
- ^ Гурвич О.Л., Баранов П.В., Гестеланд Р.Ф., Аткинс Дж.Ф. (июнь 2005 г.). «Уровни экспрессии влияют на сдвиг рамки рибосомы в тандемных редких кодонах аргинина AGG_AGG и AGA_AGA в Escherichia coli» . Журнал бактериологии . 187 (12): 4023–4 дои : 10.1128/JB.187.12.4023–4032.2005 . ПМЦ 1151738 . ПМИД 15937165 .
- ^ Калискан Н., Катунин В.И., Белардинелли Р., Песке Ф., Роднина М.В. (июнь 2014 г.). «Запрограммированный сдвиг рамки -1 за счет кинетического разделения во время затрудненной транслокации» . Клетка . 157 (7): 1619–1631. дои : 10.1016/j.cell.2014.04.041 . ПМЦ 7112342 . ПМИД 24949973 .
- ^ Артур Л., Павлович-Джуранович С., Смит-Кутмоу К., Грин Р., Щесны П., Джуранович С. (июль 2015 г.). «Контроль трансляции с помощью лизин-кодирующих А-богатых последовательностей» . Достижения науки . 1 (6): e1500154. Бибкод : 2015SciA....1E0154A . дои : 10.1126/sciadv.1500154 . ПМЦ 4552401 . ПМИД 26322332 .
- ^ Хансен Т.М., Рейхани С.Н., Оддершеде Л.Б., Соренсен М.А. (апрель 2007 г.). «Корреляция между механической прочностью псевдоузлов информационной РНК и сдвигом рамки рибосомы» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (14): 5830–5835. Бибкод : 2007PNAS..104.5830H . дои : 10.1073/pnas.0608668104 . ПМК 1838403 . ПМИД 17389398 .
- ^ Перейти обратно: а б с Номенклатурный комитет Международного биохимического союза (NC-IUB) (1984). «Номенклатура не полностью определенных оснований в последовательностях нуклеиновых кислот» . Проверено 4 февраля 2008 г.
- ^ Мазаурик М.Х., Личнар П., Прер М.Ф., Canal I, Fayet O (июль 2008 г.). «Псевдоузлы РНК апикальная петля-внутренняя петля: новый тип стимулятора трансляционного сдвига рамки -1 у бактерий» . Журнал биологической химии . 283 (29): 20421–20432. дои : 10.1074/jbc.M802829200 . ПМИД 18474594 .
- ^ Иванов И.П., Андерсон С.Б., Гестеланд РФ, Аткинс Дж.Ф. (июнь 2004 г.). «Идентификация нового антизимного мРНК +1, стимулирующего сдвиг рамки считывания, у подмножества разнообразных беспозвоночных и его очевидное отсутствие у промежуточных видов» . Журнал молекулярной биологии . 339 (3): 495–504. дои : 10.1016/j.jmb.2004.03.082 . ПМЦ 7125782 . ПМИД 15147837 .
- ^ Баранов П.В., Хендерсон К.М., Андерсон С.Б., Гестеланд Р.Ф., Аткинс Дж.Ф., Ховард М.Т. (февраль 2005 г.). «Программированный сдвиг рамки рибосом при расшифровке генома SARS-CoV» . Вирусология . 332 (2): 498–510. дои : 10.1016/j.virol.2004.11.038 . ПМЦ 7111862 . ПМИД 15680415 .
- ^ Ларсен Б., Гестеланд Р.Ф., Аткинс Дж.Ф. (август 1997 г.). «Структурное зондирование и мутагенный анализ стволовой петли, необходимой для рибосомального сдвига рамки ДНК ДНК Escherichia coli: запрограммированная эффективность 50%» . Журнал молекулярной биологии . 271 (1): 47–60. дои : 10.1006/jmbi.1997.1162 . ПМК 7126992 . ПМИД 9300054 .
Внешние ссылки
[ редактировать ]- Сдвиг рамки, + рибосомы Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- Wise2 — выравнивает белок по последовательности ДНК , допуская сдвиги рамки и интроны.
- FastY — сравнение последовательности ДНК с базой данных последовательностей белков , с учетом пропусков и сдвигов рамки считывания.
- Путь. Архивировано 19 июля 2011 г. в Wayback Machine — инструменте, который сравнивает два сдвига рамки считывания белка ( обратной трансляции ). принцип
- Recode2 — База данных перекодированных генов, в том числе тех, которые требуют запрограммированного трансляционного сдвига рамки считывания.