Полиамин
Полиамин имеющее – органическое соединение, более двух аминогрупп . Алкилполиамины встречаются в природе, но некоторые из них являются синтетическими. Алкилполиамины бесцветны, гигроскопичны и водорастворимы. При нейтральном pH они существуют в виде производных аммония. [1] Большинство ароматических полиаминов представляют собой кристаллические твердые вещества при комнатной температуре.
Природные полиамины
[ редактировать ]Низкомолекулярные линейные полиамины встречаются во всех формах жизни. Основными примерами являются триамин спермидин и тетраамин спермин . Структурно и биосинтетически они родственны диаминам путресцину и кадаверину . Метаболизм полиаминов регулируется активностью фермента орнитиндекарбоксилазы (ОДК). [2] Полиамины обнаруживаются в высоких концентрациях в млекопитающих мозге . [3]
- Природные полиамины


Синтетические полиамины
[ редактировать ]Этиленамины представляют собой коммерчески важный класс синтетических полиаминов с этиленом ( −CH 2 CH 2 − связи); в 2001 году мировая производственная мощность оценивалась в 385 000 тонн. [4] Они представляют собой химические промежуточные продукты, часто используемые для производства поверхностно-активных веществ и в качестве сшивающих агентов для эпоксидных смол. [5] Некоторые интересные представители этого класса включают в себя:
- Этилендиамин , первый представитель этого ряда. Он сам по себе является хелатирующим лигандом и предшественником популярного секвестранта металлов ЭДТА (этилендиаминтетрауксусной кислоты). Перметилированный этилендиамин дает тетраметилэтилендиамин (ТМЕДА), который имеет очень высокое сродство к ионам лития. [6]
- Макроциклические полиамины, аналогичные краун-эфирам : 1,4,7-триазациклононан ( (NHCH 2 CH 2 ) 3 ) и циклы ( (NHCH 2 CH 2 ) 4 ). Родственным тетрааза макроциклом является циклам .
- Трис(2-аминоэтил)амин ( N(CH 2 CH 2 NH 2 ) 3 ) представляет собой разветвленный полиамин, который является второстепенным побочным продуктом процесса получения полиэтиленамина . Родственным триподальным полиамином является 1,1,1-трис(аминометил)этан . Это интересные хелатирующие лиганды.
- Полиэтиленимин представляет собой полимер, полученный из азиридина .
Другие синтетические полиамины включают 1,3,5-триазинан (не путать с 1,3,5-триазином ) и N-замещенные аналоги. Метилен ( −CH 2 ) связи образуются из формальдегида . Продукт реакции моноэтаноламина и формальдегида известен в промышленности как «МЭА триазин» (на самом деле это триазинан) и служит водорастворимым поглотителем сероводорода . [7] Гексаметилентетрамин (гексамин) — еще один продукт формальдегида и аммиака, имеющий различное применение в промышленности. В домашних условиях его используют как твердое походное топливо. В лаборатории он реагирует с алкилгалогенидами для селективного получения первичных аминов по реакции Делепина .
- Синтетические полиамины

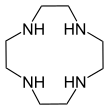
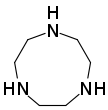

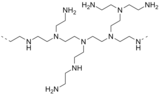 Субъединица полиэтиленимина
Субъединица полиэтиленимина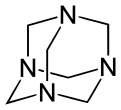






Биологическая функция
[ редактировать ]Хотя известно, что биосинтез полиаминов строго регулируется, биологическая функция полиаминов изучена лишь частично. В катионной аммониевой форме они связываются с ДНК и по структуре представляют собой соединения, катионы которых находятся через равные промежутки времени (в отличие от Mg 2+
или Ca 2+
, которые являются точечными зарядами). Также было обнаружено, что они действуют как промоторы запрограммированного сдвига рамки рибосомы во время трансляции. [8]
Ингибирование биосинтеза полиаминов замедляет или останавливает рост клеток . Поступление экзогенных полиаминов восстанавливает рост этих клеток. Большинство эукариотических клеток экспрессируют АТФазу, транспортирующую полиамины на своей клеточной мембране , которая облегчает интернализацию экзогенных полиаминов. Эта система высокоактивна в быстро пролиферирующих клетках и является мишенью некоторых химиотерапевтических препаратов, находящихся в стадии разработки. [9]
Полиамины также являются модуляторами различных ионных каналов , включая рецепторы NMDA и рецепторы AMPA . Они блокируют внутренние выпрямляющие калиевые каналы , так что токи каналов выпрямляются внутрь, тем самым, клеточная энергия, т. е. K +
Градиент ионов через клеточную мембрану сохраняется. Кроме того, полиамины участвуют в инициации экспрессии SOS-ответа оперона колицина E7 и подавляют белки, которые необходимы для поглощения колицина E7, тем самым обеспечивая преимущество в выживании продуцирующей колицин E. coli в условиях стресса. [10]
Полиамины могут повышать проницаемость гематоэнцефалического барьера . [11]
Они участвуют в модуляции старения органов растений и поэтому считаются растительными гормонами . [12] Кроме того, они принимают непосредственное участие в регуляции запрограммированной гибели клеток. [13]
Репарация ДНК, направленная на гомологию
[ редактировать ]Полиамины способствуют опосредованному гомологичной рекомбинацией (HR) восстановлению двухцепочечного разрыва (DSB), . [14] Полиамины усиливают обменную активность RAD51 рекомбиназы . Истощение полиаминов повышает чувствительность клеток к генотоксичным веществам, таким как ионизирующая радиация и ультрафиолетовое излучение. Влияние полиаминов на RAD51 обусловлено их способностью усиливать захват гомологичной дуплексной ДНК и способствовать опосредованному RAD-51 спариванию гомологичных ДНК и обменной активности. [14] Полиамины, по-видимому, играют эволюционно консервативную роль в регуляции активности рекомбиназы.
Биосинтез спермидина, спермина, термоспермина.
[ редактировать ]
Спермидин синтезируется из путресцина с использованием аминопропильной группы декарбоксилированного S -аденозил-L-метионина (SAM), S -аденозилметионинамина . Реакцию катализирует спермидинсинтаза . [15]
Спермин синтезируется в результате реакции спермидина с SAM в присутствии фермента сперминсинтазы .
Полиамины подвергаются быстрому взаимопревращению в полиаминовом цикле, в котором путресцин приводит к синтезу спермидина и спермина с деградацией этих полиаминов с образованием путресцина, который может начать цикл снова. [15]
Термоспермин ( NH 2 -(CH 2 ) 3 -NH-(CH 2 ) 3 -NH-(CH 2 ) 4 -NH 2 ) представляет собой структурный изомер спермина и новый тип регулятора роста растений. Он производится из спермидина под действием термосперминсинтазы , которая кодируется геном ACAULIS5 (ACL5). [16]
Аналоги полиаминов
[ редактировать ]Критическая роль полиаминов в росте клеток привела к разработке ряда агентов, которые мешают метаболизму полиаминов. Эти агенты используются в терапии рака . Аналоги полиаминов повышают регуляцию р53 в клетке, что приводит к ограничению пролиферации и апоптозу . [17] Он также снижает экспрессию альфа-рецептора эстрогена при ER-положительном раке молочной железы. [18]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Эллер, Карстен; Хенкес, Эрхард; Россбахер, Роланд; Хёке, Хартмут (2005). «Амины алифатические». Энциклопедия промышленной химии Ульмана . Вайнхайм: Wiley-VCH. дои : 10.1002/14356007.a02_001 . ISBN 978-3527306732 .
- ^ Пегг, А.Е.; Макканн, П.П. (1982). «Метаболизм и функция полиаминов». Американский журнал физиологии . 243 (5): 212–221. дои : 10.1152/ajpcell.1982.243.5.C212 . ПМИД 6814260 . S2CID 21063248 .
- ^ Зайлер, Н. (1982). «Полиамины» . В Лайте, А. (ред.). Химическая и клеточная архитектура . Справочник по нейрохимии. Том. 1 (2-е изд.). Спрингер. стр. 223–255. дои : 10.1007/978-1-4757-0614-7_9 . ISBN 978-1-4757-0614-7 .
- ^ Шривасан Шридхар; Ричард Г. Картер (2001). «Диамины и высшие амины алифатические». Энциклопедия химической технологии Кирка-Отмера . Нью-Йорк: Джон Уайли. дои : 10.1002/0471238961.0409011303011820.a01.pub2 . ISBN 9780471238966 .
- ^ Лоуренс, Стивен А. (2004). Амины: синтез, свойства и применение . Издательство Кембриджского университета. п. 64. ИСБН 978-0-521-78284-5 .
- ^ Хейнс, РК; Фонвиллер, Южная Каролина; Людерер, MR (2006). «N,N,N',N'-тетраметилэтилендиамин». В Пакетте, Л. (ред.). N , N , N ', N' -Тетраметилэтилендиамин . Энциклопедия реагентов для органического синтеза . Уайли. дои : 10.1002/047084289X.rt064.pub2 . ISBN 0-471-93623-5 .
- ^ Дж. Н. Тейлор; Джей Джей Уайлд; Т. Мюллер; Дж. Мьюрисон; Ф. Шнайдер (2017). Свежий взгляд на механизм удаления H2S МЭА-триазина и ММА-триазина . Международная конференция SPE по нефтепромысловой химии. Монтгомери, Техас. дои : 10.2118/184529-MS .
- ^ Рато С; Амирова С.Р.; Бейтс Д.Г.; Стэнсфилд I; Уоллес Х.М. (июнь 2011 г.). «Трансляционное перекодирование как контроллер обратной связи: системные подходы выявляют специфичные для полиаминов эффекты на сдвиг рамки антизимной рибосомы» . Нуклеиновые кислоты Рез . 39 (11): 4587–97. дои : 10.1093/нар/gkq1349 . ПМК 3113565 . ПМИД 21303766 .
- ^ Ван С., Делькрос Дж.Г., Кэннон Л., Конате Ф., Кариас Х., Биггерстафф Дж., Гарднер Р.А., Фэнстиел IV О (ноябрь 2003 г.). «Определение молекулярных требований для селективной доставки конъюгатов полиаминов в клетки, содержащие активные переносчики полиаминов». J Med Chem . 46 (24): 5129–38. дои : 10.1021/jm030223a . ПМИД 14613316 .
- ^ И-Сюань Пан; Чен-Чунг Ляо (май 2006 г.). «Важнейшая роль полиаминов, регулирующих выработку ColE7 и ограничивающих поглощение ColE7 продуцирующей колицин Escherichia coli» . Ж. Биол. Хим . 281 (19): 13083–91. дои : 10.1074/jbc.M511365200 . ПМИД 16549429 .
- ^ Чжан Л., Ли Х.К., Прюсс Т.Х., Уайт Х.С., Булай Г. (март 2009 г.). «Синтез и применение полиаминовых аминокислотных остатков: повышение биологической активности анальгетического нейропептида нейротензина» . Дж. Мед. Хим . 52 (6): 1514–7. дои : 10.1021/jm801481y . ПМК 2694617 . ПМИД 19236044 .
- ^ Панди С., Ранаде С.А., Нагар П.К., Кумар Н. (сентябрь 2000 г.). «Роль полиаминов и этилена как модуляторов старения растений». Дж. Биоски . 25 (3): 291–9. дои : 10.1007/BF02703938 . ПМИД 11022232 . S2CID 21925829 .
- ^ Москва, ПН; Рубелакис-Ангелакис, штат Калифорния (11 ноября 2013 г.). «Полиамины и запрограммированная смерть клеток» . Журнал экспериментальной ботаники . 65 (5): 1285–96. дои : 10.1093/jxb/ert373 . ПМИД 24218329 .
- ^ Перейти обратно: а б Ли САЙ, Су Г.К., Хуан Вайоминг, Ко МЮ, Йе ХИ, Чанг Г.Д., Линь С.Дж., Чи П (январь 2019 г.). «Стимулирование репарации ДНК, направленной на гомологию, с помощью полиаминов» . Нат Коммун . 10 (1): 65. Бибкод : 2019NatCo..10...65L . дои : 10.1038/s41467-018-08011-1 . ПМК 6325121 . ПМИД 30622262 .
- ^ Перейти обратно: а б Пал М, Салаи Г, Янда Т (2015). «Предположение: полиамины играют важную роль в передаче сигналов абиотического стресса» (PDF) . Наука о растениях . 237 : 16–23. Бибкод : 2015PlnSc.237...16P . doi : 10.1016/j.plantsci.2015.05.003 . ПМИД 26089148 .
- ^ Такано, А; Какехи, Дж; Такахаши, Т. (апрель 2012 г.). «Термоспермин не является второстепенным полиамином в царстве растений» . Физиол растительной клетки . 53 (4): 606–16. дои : 10.1093/pcp/pcs019 . ПМИД 22366038 .
- ^ Хуан, И; Пледжи, Эллисон; Рубин, Этель; Мартон, Лоуренс Дж.; Уостер, Патрик М.; Сукумар, Сарасвати; Касеро, Роберт А.; Дэвидсон, Нэнси Э. (сентябрь 2005 г.). «Роль p53/p21 (Waf1/Cip1) в регуляции индуцированного аналогом полиамина ингибирования роста и гибели клеток в клетках рака молочной железы человека» . Биология и терапия рака . 4 (9): 1006–13. дои : 10.4161/cbt.4.9.1970 . ПМЦ 3639297 . ПМИД 16131835 .
- ^ Хуанг, Ю; Кин, Дж. К.; Пледжи, А; Мартон, LJ; Чжу, Т; Сукумар, С; Парк, Британская Колумбия; Блэр, Б; Бреннер, К; Касеро, РА-младший; Дэвидсон, Нью-Йорк (2006). «Аналоги полиаминов подавляют экспрессию альфа-рецептора эстрогена в клетках рака молочной железы человека» . J Биол Хим . 281 (28): 19055–63. дои : 10.1074/jbc.M600910200 . ПМЦ 3623667 . ПМИД 16679312 .
Внешние ссылки
[ редактировать ]- Полиамины в пролиферации клеточного цикла и гибели клеток
- Килпеляйнен, Пекка (2002). Экспрессия и регуляция в мозге крыс и трансгенных мышей . Кафедра биохимии Университета Оулу. hdl : 10024/37387 . ISBN 951-42-6631-5 . Обширный обзор литературы за 2001 г. по структуре, свойствам, метаболизму полиаминов у млекопитающих, а также их физиологической и патофизиологической роли (см. статью «Содержание»).