Орнитиндекарбоксилаза
| Орнитиндекарбоксилаза | |||
|---|---|---|---|
 Димер орнитиндекарбоксилазы, человек | |||
| Идентификаторы | |||
| Номер ЕС. | 4.1.1.17 | ||
| Номер CAS. | 9024-60-6 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| орнитиндекарбоксилаза | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ОДК1 | ||
| ген NCBI | 4953 | ||
| HGNC | 8109 | ||
| МОЙ БОГ | 165640 | ||
| RefSeq | НМ_002539 | ||
| ЮниПрот | P11926 | ||
| Другие данные | |||
| Номер ЕС | 4.1.1.17 | ||
| Локус | Хр. 2 стр25 | ||
| |||
Фермент орнитиндекарбоксилаза ( EC 4.1.1.17 катализирует декарбоксилирование орнитина , ODC ) (продукта цикла мочевины ) с образованием путресцина . Эта реакция является обязательным этапом синтеза полиаминов . [1] У человека этот белок содержит 461 аминокислоту и образует гомодимер . [2]
У человека орнитиндекарбоксилаза (ODC) экспрессируется геном ODC1 . Белок ODC иногда называют «ODC1» в исследованиях, касающихся людей и мышей, но некоторые виды, такие как дрозофила ( dODC2 ), [3] виды Пасленовые семейства растений ( ODC2 ), [4] и молочнокислые бактерии Paucilactobacillus wasatchensis ( odc2 ). [5] Было показано, что у них есть второй ген ODC.
Механизм реакции
[ редактировать ]Лизин 69 на орнитиндекарбоксилазе (ODC) связывает кофактор пиридоксальфосфат с образованием основания Шиффа . [6] Орнитин замещает лизин с образованием основания Шиффа, присоединенного к ортонину, который декарбоксилируется с образованием хиноидного промежуточного соединения. Это промежуточное соединение перестраивается с образованием основания Шиффа, присоединенного к путресцину , который подвергается атаке лизина с высвобождением путресцинового продукта и реформированием PLP-связанного ODC. [7] Это первый и лимитирующий этап производства полиаминов у людей , соединений, необходимых для деления клеток .

Затем спермидинсинтаза может катализировать превращение путресцина в спермидин путем присоединения аминопропильного фрагмента. [8] Спермидин является предшественником других полиаминов, таких как спермин и его структурный изомер термоспермин .
Структура
[ редактировать ]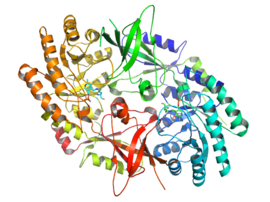
Активная форма орнитиндекарбоксилазы представляет собой гомодимер . Каждый мономер содержит бочкообразный домен, состоящий из альфа-бета-цилиндра , и листовой домен, состоящий из двух бета-листов . Домены соединены петлями. Мономеры соединяются друг с другом посредством взаимодействия между стволом одного мономера и слоем другого. Связывание между мономерами относительно слабое, и ODC быстро преобразуется между мономерными и димерными формами в клетке. [1]
Кофактор пиридоксальфосфат связывает лизин 69 на С- конце бочкообразного домена. Активный центр находится на границе раздела двух доменов, в полости, образованной петлями обоих мономеров. [1]
Функция
[ редактировать ]Реакция декарбоксилирования орнитина, катализируемая орнитиндекарбоксилазой, является первой и важной стадией синтеза полиаминов , особенно путресцина , спермидина и спермина . Полиамины важны для стабилизации структуры ДНК ДНК , пути восстановления двухцепочечного разрыва и в качестве антиоксидантов . Таким образом, орнитиндекарбоксилаза является важным ферментом для роста клеток, производящим полиамины, необходимые для стабилизации вновь синтезированной ДНК. Недостаток ODC вызывает апоптоз клеток у эмбрионов мышей, вызванный повреждением ДНК. [10]
Протеосомная деградация
[ редактировать ]ODC является наиболее хорошо изученным клеточным белком, подверженным убиквитин -независимой протеасомной деградации. Хотя большинство белков сначала необходимо пометить несколькими молекулами убиквитина, прежде чем они связываются и разрушаются протеасомой , деградация ODC вместо этого опосредована несколькими сайтами узнавания на белке и антизимом его дополнительного фактора . Процесс деградации ODC регулируется по принципу отрицательной обратной связи продуктами его реакции. [11]
До отчета Sheaff et al. (2000), [12] которые продемонстрировали, что ингибитор циклин-зависимой киназы (Cdk) p21Cip1 также расщепляется протеасомой убиквитин-независимым образом, ODC был единственным ярким примером убиквитин-независимой протеасомной деградации. [13]
Клиническое значение
[ редактировать ]ODC является транскрипционной мишенью онкогена Myc . [14] и его активность повышается при различных видах рака. Полиаминные продукты пути, инициализируемого ODC, связаны с усиленным ростом клеток и снижением апоптоза . [15] Ультрафиолетовый свет , [16] асбест [17] и андрогены, выделяемые предстательной железой [18] Известно, что все они вызывают повышенную активность ODC, связанную с раком. ингибиторы ODC, такие как эфлорнитин, эффективно уменьшают раковые заболевания на животных моделях. Было показано, что [19] и препараты, нацеленные на ODC, проходят испытания на предмет потенциального клинического использования.Механизм, с помощью которого ODC способствует канцерогенезу , сложен и до конца не изучен. Помимо прямого влияния на стабильность ДНК, полиамины также активируют щелевых соединений. гены [20] и подавлять плотных соединений гены . Гены щелевых соединений участвуют в коммуникации между канцерогенными клетками, а гены плотных соединений действуют как супрессоры опухолей. [15]
мутации гена ODC1 вызывают синдром Бахмана-Буппа (BABS), редкое нейрометаболическое расстройство, характеризующееся глобальной задержкой развития, алопецией, макроцефалией, дисморфическими особенностями и поведенческими отклонениями. Было показано, что [21] BABS обычно вызывается аутосомно-доминантным de novo вариантом ODC1 . [21]
Экспрессия гена ODC индуцируется большим количеством биологических стимулов, включая судорожную активность в головном мозге. [22] Инактивация ODC дифторметилорнитином (ДМФО, эфлорнитин ) используется для лечения рака и роста волос на лице у женщин в постменопаузе.
ODC также является ферментом, незаменимым для таких паразитов , как Trypanosoma , Giardia и Plasmodium , и этот факт используется в препарате эфлорнитин . [23]
Иммунологическое значение
[ редактировать ]В Т-клетках, активированных антигеном, ферментативная активность ODC увеличивается после активации, что соответствует увеличению синтеза полиаминов в Т-клетках после активации. [24] Как и в случае с ODC и раком, MYC , также называемый c-Myc от клеточного Myc, является главным регулятором биосинтеза полиаминов в Т-клетках. [25]
Исследование 2020 года, проведенное Wu et al. специфичных для Т-клеток Использование мышей cKO ODC, , показало, что Т-клетки могут нормально функционировать и пролиферировать in vivo , а другие пути синтеза полиаминов могут компенсировать недостаток ODC. [26] Однако блокирование синтеза полиаминов посредством ODC с помощью DMFO и поглощения полиаминов с помощью AMXT 1501 истощало пул полиаминов, ингибировало пролиферацию Т-клеток и подавляло воспаление Т-клеток. [26]
Недавние исследования показали важность синтеза ODC и полиаминов в определении судьбы Т-хелперных клеток . [27] Исследование 2021 года, проведенное Puleston et al. показали, что TH 1 ) клетки и TH клетки 2 экспрессируют более высокие уровни ODC, чем регуляторные T (T reg и клетки TH , что соответствует более высоким 17 уровням биосинтеза полиаминов в TH 1 и TH 2 . [28] Исследование Wagner et al., проведенное в 2021 году. показал продвижение программы Treg в Odc1 -/- мыши. [29] Они пришли к выводу, что экспрессия ферментов, связанных с полиамином, усиливается в патогенных TH 17 и подавляется в клетках Treg. [29]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Керн А.Д., Оливейра М.А., Коффино П., Хакерт М.Л. (май 1999 г.). «Структура орнитиндекарбоксилазы млекопитающих при разрешении 1,6 А: стереохимическое значение PLP-зависимых декарбоксилаз аминокислот» . Структура . 7 (5): 567–581. дои : 10.1016/S0969-2126(99)80073-2 . ПМИД 10378276 .
- ^ Пегг А.Е. (май 2006 г.). «Регуляция орнитиндекарбоксилазы» . Журнал биологической химии . 281 (21): 14529–14532. дои : 10.1074/jbc.R500031200 . ПМИД 16459331 .
- ^ Ром Э, Кахана С (1993). «Выделение и характеристика локуса орнитиндекарбоксилазы дрозофилы: доказательства присутствия двух транскрибируемых генов ODC в геноме дрозофилы». ДНК и клеточная биология . 12 (6): 499–508. дои : 10.1089/dna.1993.12.499 . ПМИД 8329117 .
- ^ Цинь X, Chetelat RT (май 2021 г.). «Гены орнитиндекарбоксилазы способствуют независимому от S-РНКазы отторжению пыльцы» . Физиология растений . 186 (1): 452–468. дои : 10.1093/plphys/kiab062 . ПМК 8154068 . PMID 33576789 .
- ^ Берту Х., Векслер Д., Ирмлер С. (03.03.2022). «Производство путресцина и кадаверина Paucilactobacillus wasatchensis » . Границы микробиологии . 13 : 842403. дои : 10.3389/fmicb.2022.842403 . ПМЦ 8928434 . ПМИД 35308356 .
- ^ Оливейра Э.Ф., Серкейра Н.М., Фернандес П.А., Рамос М.Дж. (октябрь 2011 г.). «Механизм образования внутреннего альдимина в пиридоксаль-5'-фосфатзависимых ферментах». Журнал Американского химического общества . 133 (39): 15496–15505. дои : 10.1021/ja204229m . ПМИД 21854048 .
- ^ Брукс Х.Б., Филлипс Массачусетс (декабрь 1997 г.). «Характеристика механизма реакции орнитиндекарбоксилазы Trypanosoma brucei с помощью многоволновой спектроскопии с остановленным потоком». Биохимия . 36 (49): 15147–15155. дои : 10.1021/bi971652b . ПМИД 9398243 .
- ^ Тети Д., Визалли М., Макнейр Х. (декабрь 2002 г.). «Анализ полиаминов как маркеров (пато) физиологических состояний». Журнал хроматографии. Б. Аналитические технологии в биомедицине и науках о жизни . 781 (1–2): 107–149. дои : 10.1016/S1570-0232(02)00669-4 . ПМИД 12450656 .
- ^ PDB : 1d7k ; Алмруд Дж.Дж., Оливейра М.А., Керн А.Д., Гришин Н.В., Филлипс М.А., Хакерт М.Л. (январь 2000 г.). «Кристаллическая структура орнитиндекарбоксилазы человека при разрешении 2,1 А: структурное понимание связывания антизима». Журнал молекулярной биологии . 295 (1): 7–16. дои : 10.1006/jmbi.1999.3331 . ПМИД 10623504 . ; рендерится через PyMOL .
- ^ Пендевиль Х., Карпино Н., Марин Дж. К., Такахаши Ю., Мюллер М., Марсьяль Дж. А., Кливленд Дж. Л. (октябрь 2001 г.). «Ген орнитиндекарбоксилазы необходим для выживания клеток на ранних стадиях развития мышей» . Молекулярная и клеточная биология . 21 (19): 6549–6558. дои : 10.1128/MCB.21.19.6549-6558.2001 . ПМК 99801 . ПМИД 11533243 .
- ^ Чжан М., Пиккарт С.М., Коффино П. (апрель 2003 г.). «Детерминанты распознавания протеасом орнитиндекарбоксилазы, убиквитин-независимого субстрата» . Журнал ЭМБО . 22 (7): 1488–1496. дои : 10.1093/emboj/cdg158 . ПМК 152902 . ПМИД 12660156 .
- ^ Шифф Р.Дж., Сингер Дж.Д., Свангер Дж., Смитерман М., Робертс Дж.М., Клерман Б.Е. (февраль 2000 г.). «Протеосомный оборот p21Cip1 не требует убиквитинирования p21Cip1» . Молекулярная клетка . 5 (2): 403–410. дои : 10.1016/S1097-2765(00)80435-9 . ПМИД 10882081 .
- ^ Верма Р., Деше Р.Дж. (май 2000 г.). «Протеасомный хаудунит: случай отсутствия сигнала» . Клетка . 101 (4): 341–344. дои : 10.1016/S0092-8674(00)80843-0 . ПМИД 10830160 . S2CID 18425370 .
- ^ Белло-Фернандес К., Пакхэм Дж., Кливленд Дж.Л. (август 1993 г.). «Ген орнитиндекарбоксилазы является мишенью транскрипции c-Myc» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (16): 7804–7808. Бибкод : 1993PNAS...90.7804B . дои : 10.1073/pnas.90.16.7804 . ПМЦ 47231 . ПМИД 8356088 .
- ^ Перейти обратно: а б Гернер Э.В., Мейскенс, Флорида (октябрь 2004 г.). «Полиамины и рак: старые молекулы, новое понимание» . Обзоры природы. Рак . 4 (10): 781–792. дои : 10.1038/nrc1454 . ПМИД 15510159 . S2CID 37647479 .
- ^ Ахмад Н., Гиллиам А.С., Катияр С.К., О'Брайен Т.Г., Мухтар Х (сентябрь 2001 г.). «Определенная роль орнитиндекарбоксилазы в фотоканцерогенезе» . Американский журнал патологии . 159 (3): 885–892. дои : 10.1016/S0002-9440(10)61764-6 . ПМК 1850478 . ПМИД 11549581 .
- ^ Марш Дж.П., Моссман Б.Т. (январь 1991 г.). «Роль асбеста и активных форм кислорода в активации и экспрессии орнитиндекарбоксилазы в эпителиальных клетках трахеи хомяка». Исследования рака . 51 (1): 167–173. ПМИД 1846307 .
- ^ Кроза А., Палвимо Дж. Дж., Юлкунен М., Янне О. А. (март 1992 г.). «Сравнение андрогенной регуляции экспрессии генов орнитиндекарбоксилазы и S-аденозилметиониндекарбоксилазы в почках и дополнительных половых органах грызунов». Эндокринология . 130 (3): 1131–1144. дои : 10.1210/endo.130.3.1537280 . ПМИД 1537280 .
- ^ Мейскенс Флорида, Гернер Э.В. (май 1999 г.). «Разработка дифторметилорнитина (ДФМО) в качестве химиопрофилактического средства». Клинические исследования рака . 5 (5): 945–951. ПМИД 10353725 .
- ^ Шор Л., Маклин П., Гилмор С.К., Ходжинс М.Б., Финбоу М.Э. (июль 2001 г.). «Полиамины регулируют связь через щелевые соединения в клетках, экспрессирующих коннексин 43» . Биохимический журнал . 357 (Часть 2): 489–495. дои : 10.1042/0264-6021:3570489 . ПМК 1221976 . ПМИД 11439099 .
- ^ Перейти обратно: а б Бапп С., Майкл Дж., ВанСикл Э., Раджасекаран С., Бахманн А.С. (1993). «Синдром Бахмана-Буппа». Адам М.П., Мирзаа Г.М., Пагон Р.А., Уоллес С.Е. (ред.). Джин Обзоры . Сиэтл (Вашингтон): Вашингтонский университет, Сиэтл. ПМИД 36007106 . Проверено 28 июня 2023 г.
- ^ Херберг Л.Дж., Роуз И.С., де Беллерош Дж.С., Минц М. (март 1992 г.). «Индукция орнитиндекарбоксилазы и синтез полиаминов при возникновении судорог: эффект альфа-дифторметилорнитина». Исследования эпилепсии . 11 (1): 3–7. дои : 10.1016/0920-1211(92)90015-Л . ПМИД 1563337 . S2CID 1221264 .
- ^ Хеби О., Перссон Л., Рентала М. (август 2007 г.). «Нацеливание на ферменты биосинтеза полиаминов: многообещающий подход к терапии африканской сонной болезни, болезни Шагаса и лейшманиоза». Аминокислоты . 33 (2): 359–366. дои : 10.1007/s00726-007-0537-9 . ПМИД 17610127 . S2CID 26273053 .
- ^ Хестерберг Р.С., Кливленд Дж.Л., Эплинг-Бернетт ПК (март 2018 г.). «Роль полиаминов в функциях иммунных клеток» . Медицинские науки . 6 (1): 22. doi : 10.3390/medsci6010022 . ПМЦ 5872179 . ПМИД 29517999 .
- ^ Миллер Д.М., Томас С.Д., Ислам А., Мюнх Д., Седорис К. (октябрь 2012 г.). «c-Myc и метаболизм рака» . Клинические исследования рака . 18 (20): 5546–5553. дои : 10.1158/1078-0432.CCR-12-0977 . ПМЦ 3505847 . ПМИД 23071356 .
- ^ Перейти обратно: а б Ву Р., Чен X, Кан С., Ван Т., Гнанапракасам Дж.Р., Яо Ю. и др. (декабрь 2020 г.). «Путь синтеза и спасения de novo координально регулирует гомеостаз полиаминов и определяет пролиферацию и функцию Т-клеток» . Достижения науки . 6 (51). Бибкод : 2020SciA....6.4275W . дои : 10.1126/sciadv.abc4275 . ПМЦ 7744078 . ПМИД 33328226 .
- ^ Ши Х, Чи Х (август 2021 г.). «Полиамин: метаболический компас для направления судьбы Т-хелперных клеток» . Клетка . 184 (16): 4109–4112. дои : 10.1016/j.cell.2021.07.012 . ПМИД 34358466 .
- ^ Пулестон Д.Д., Байшаули Ф., Санин Д.Е., Эдвардс-Хикс Дж., Вилла М., Кабат А.М. и др. (август 2021 г.). «Метаболизм полиаминов является центральным определяющим фактором верности линии Т-хелперных клеток» . Клетка . 184 (16): 4186–4202.e20. дои : 10.1016/j.cell.2021.06.007 . ПМЦ 8358979 . ПМИД 34216540 .
- ^ Перейти обратно: а б Вагнер А., Ван С., Фесслер Дж., ДеТомасо Д., Авила-Пачеко Дж., Камински Дж. и др. (август 2021 г.). «Метаболическое моделирование одиночных клеток Th17 выявляет регуляторы аутоиммунитета» . Клетка . 184 (16): 4168–4185.e21. дои : 10.1016/j.cell.2021.05.045 . ПМК 8621950 . ПМИД 34216539 .
Внешние ссылки
[ редактировать ]- Орнитиндекарбоксилаза на сайте herkules.oulu.fi
- Орнитин + декарбоксилаза Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)