Пропионил-КоА
 | |
| Имена | |
|---|---|
| Название ИЮПАК 3'- O -фосфоноаденозин 5'-{(3R ) -3-гидрокси-2,2-диметил-4-оксо-4-[(3-оксо-3-{[2-(пропаноилсульфанил)этил]амино} пропил)амино]бутилдигидрофосфат} | |
| Систематическое название ИЮПАК ТО 1 -{[(2R , 3S , 4R , 5R ) -5-(6-амино-9Н- пурин -9-ил)-4-гидрокси-3-(фосфоноокси)оксолан-2-ил]метил } О 3 -{(3 R )-3-гидрокси-2,2-диметил-4-оксо-4-[(3-оксо-3-{[2-(пропаноилсульфанил)этил]амино}пропил)амино]бутил}дигидрофосфат | |
| Другие имена пропионил-коэнзим А; Пропаноил-коэнзим А | |
| Идентификаторы | |
3D model ( JSmol ) | |
| ЧЭБИ | |
| ХимическийПаук | |
| Информационная карта ECHA | 100.005.698 |
| МеШ | пропионил-кофермент+А |
ПабХим CID | |
| НЕКОТОРЫЙ | |
Панель управления CompTox ( EPA ) | |
| Характеристики | |
| С 24 Ч 40 Н 7 О 17 П 3 С | |
| Молярная масса | 823.60 g/mol |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |
Пропионил-КоА представляет собой кофермента А производное пропионовой кислоты . Он состоит из 24 углеродных цепей (без кофермента это 3-углеродная структура), и его производство и метаболическая судьба зависят от того, в каком организме он присутствует. [1] К его производству могут привести несколько различных путей, например, катаболизм определенных аминокислот или окисление жирных кислот с нечетной цепью . [2] Позже он может расщепляться пропионил-КоА-карбоксилазой или посредством метилцитратного цикла. [3] Однако в различных организмах пропионил-КоА может быть изолирован в контролируемых областях, чтобы снизить его потенциальную токсичность за счет накопления. [4] Генетические недостатки, связанные с выработкой и расщеплением пропионил-КоА, также имеют большое клиническое и человеческое значение. [5]
Производство
[ редактировать ]Существует несколько различных путей производства пропионил-КоА:
- Пропионил-КоА, трехуглеродная структура, считается второстепенной разновидностью пропионовой кислоты. Следовательно, нечетные цепи жирных кислот окисляются с образованием как пропионил-КоА, так и ацетил-КоА . Пропионил-КоА позже превращается в сукцинил-КоА посредством биотин-зависимой пропионил-КоА-карбоксилазы (PCC) и b12-зависимой метилмалонил-КоА-мутазы (MCM) последовательно. [2]
- Пропионил-КоА образуется не только в результате окисления жирных кислот с нечетной цепью, но также в результате окисления аминокислот, включая метионин , валин , изолейцин и треонин . Кроме того, катаболизм аминокислот также может быть результатом превращения пропионил-КоА в метилмалонил-КоА под действием пропионил-КоА-карбоксилазы . [1]
- Окисление холестерина , в результате которого образуются желчные кислоты , также образует пропионил-КоА в качестве побочного продукта. В эксперименте, проведенном Suld et al., при сочетании митохондрий печени и пропионовой кислоты с добавлением кофермента А деградировали меченые изотопы псионовой кислоты. Однако после инкубации с 5β-холестан-3α,7α,12α,26-тетрол-26,27-C14 пропионил-КоА удалось восстановить вместе с образованием желчи . [6]
Метаболическая судьба
[ редактировать ]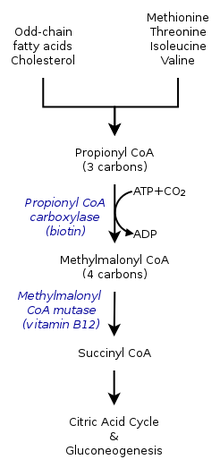
Метаболическая среде может иметь другую судьбу , (катаболическая судьба) пропионил-КоА зависит от того, в какой среде он синтезируется. Следовательно, пропионил-КоА в анаэробной чем в аэробном организме . Множественные пути, либо катаболизм пропионил-КоА-карбоксилазы, либо метилцитратсинтазы, также зависят от присутствия различных генов. [7]
Реакция с пропионил-КоА-карбоксилазой
[ редактировать ]В цикле лимонной кислоты у человека пропионил-КоА, который взаимодействует с оксалоацетатом с образованием метилцитрата, также может катализироваться с образованием метилмалонил-КоА посредством карбоксилирования пропионил -КоА-карбоксилазой (PCC). Метилмалонил-КоА позже трансформируется в сукцинил-КоА для дальнейшего использования в цикле трикарбоновых кислот . PCC не только катализирует карбоксилирование пропионил-КоА в метилмалонил-КоА, но также действует на несколько различных ацил-КоА. Тем не менее, его наибольшая аффинность связывания наблюдается с пропионил-КоА. Далее было показано, что трансформация пропионил-КоА ингибируется отсутствием нескольких маркеров TCA , таких как глутамат . Механизм показан на рисунке слева. [2]
Механизм
[ редактировать ]У млекопитающих пропионил-КоА превращается в ( S ) -метилмалонил-КоА под действием пропионил-КоА-карбоксилазы , биотин -зависимого фермента, также требующего бикарбоната и АТФ .
Этот продукт превращается в ( R )-метилмалонил-КоА под действием метилмалонил-КоА рацемазы .
( R )-Метилмалонил-КоА превращается в сукцинил-КоА , промежуточное соединение в цикле трикарбоновых кислот , под действием метилмалонил-КоА-мутазы , фермента, требующего

кобаламин катализирует миграцию углерод-углеродных связей.
Мутазный механизм метилмалонил -КоА начинается с разрыва связи между 5'- СН
2 - 5'-дезоксиаденозила и кобальта, находящегося в степени окисления 3+ (III), который дает 5'- дезоксиаденозильный радикал и кобаламин в восстановленной степени окисления Co(II).
Затем этот радикал отрывает атом водорода от метильной группы метилмалонил-КоА, в результате чего образуется радикал метилмалонил-КоА. Считается, что этот радикал образует углерод-кобальтовую связь с коферментом, за которой затем следует перестройка углеродного скелета субстрата, образуя радикал сукцинил-КоА. Затем этот радикал отрывает водород от ранее полученного 5'-дезоксиаденозина, снова создавая дезоксиаденозильный радикал, который атакует кофермент, реформируя исходный комплекс.
Дефект фермента мутазы метилмалонил-КоА приводит к метилмалоновой ацидурии — опасному заболеванию, которое вызывает снижение pH крови. [8]

Метилцитратный цикл
[ редактировать ]Накопление пропионил-КоА может оказаться токсичным для различных организмов. Поскольку были предложены различные циклы превращения пропионил-КоА в пируват, одним из изученных механизмов является метилцитратный цикл . Начальной реакцией является бета-окисление с образованием пропионил-КоА, который далее расщепляется в ходе цикла. В этом пути участвуют ферменты, связанные как с циклом метилцитрата, так и с циклом лимонной кислоты . Все это способствует общей реакции по детоксикации бактерий от вредного пропионил-КоА. Его также считают результатом катаболизма жирных кислот в микобактериях. [3] Чтобы продолжить, ген prpC кодирует метилцитратсинтазу, и если он отсутствует, метилцитратный цикл не произойдет. Вместо этого катаболизм протекает через пропионил-КоА-карбоксилазу. [7] Этот механизм показан ниже слева вместе с участвующими реагентами, продуктами, промежуточными продуктами и ферментами.
Бактериальный метаболизм
[ редактировать ]микобактерий туберкулеза Метаболизм
[ редактировать ]На окисление пропионил-КоА с образованием пирувата влияет его необходимость в микобактериях туберкулеза . Накопление пропионил-КоА может привести к токсическим эффектам. у Mycobacterium Tuberculosis Было высказано предположение, что метаболизм пропионил-КоА участвует в биогенезе клеточной стенки . Таким образом, отсутствие такого катаболизма может повысить восприимчивость клетки к различным токсинам, особенно к макрофагов антимикробным механизмам . Другая гипотеза относительно судьбы пропионил-КоА в M. Tuberculosis заключается в том, что, поскольку пропионил-КоА образуется в результате катаболизма жирных кислот с нечетной цепью бета, впоследствии активируется метилцитратный цикл, чтобы свести на нет любую потенциальную токсичность, действуя как буферный механизм. [11]
Возможная секвестрация у R. sphaeroides
[ редактировать ]Пропионил-КоА может оказывать множество неблагоприятных и токсических воздействий на различные виды, включая бактерии . Например, ингибирование пируватдегидрогеназы накоплением пропионил-КоА у Rhodobacter sphaeroides может оказаться смертельным. Кроме того, как и в случае с E. coli , приток пропионил-КоА у миобактерий может привести к токсичности, если не принять меры немедленно. Эта токсичность вызвана действием липидов, образующих бактерий клеточную стенку . Используя этерификацию длинноцепочечных жирных кислот, избыток пропионил-КоА может быть изолирован и сохранен в липиде триацилглицерине (ТАГ), что приводит к регулированию повышенных уровней пропионил-КоА. Такой процесс метильного разветвления жирных кислот заставляет их действовать как стоки для накопления пропиона. [4]
кишечной палочки Метаболизм
[ редактировать ]В исследовании, проведенном Луо и др., штаммы Escherichia coli использовались для изучения того, как метаболизм пропионил-КоА потенциально может привести к выработке 3-гидроксипропионовой кислоты (3-HP). Было показано, что мутация ключевого гена, участвующего в этом пути, сукцинат-КоА-трансферазы , приводит к значительному увеличению 3-HP. [7] Однако это все еще развивающаяся область, и информация по этой теме ограничена. [12]

Метаболизм растений
[ редактировать ]Метаболизм аминокислот в растениях считается спорным вопросом из-за отсутствия конкретных доказательств существования какого-либо конкретного пути. Однако было высказано предположение, что в этом участвуют ферменты, связанные с производством и использованием пропионил-КоА. С этим связан метаболизм изобутирил-КоА . Эти две молекулы считаются промежуточными продуктами метаболизма валина . Поскольку пропионат состоит в форме пропионил-КоА, было обнаружено, что пропионил-КоА превращается в β-гидроксипропионат посредством пероксисомального ферментативного пути β-окисления . Тем не менее, у растения Arabidopsis не наблюдалось ключевых ферментов превращения валина в пропионил-КоА. В ходе различных экспериментов, проведенных Лукасом и др., было высказано предположение, что у растений посредством пероксисомальных ферментов пропионил-КоА (и изобутирил-КоА ) участвуют в метаболизме многих различных субстратов (идентичность в настоящее время оценивается), а не просто валин . [13]

Метаболизм грибов
[ редактировать ]Производство пропионил-КоА посредством катаболизма также жирных кислот связано с тиоэтерификацией . В исследовании, касающемся Aspergillus nidulans , было обнаружено, что при ингибировании метилцитратсинтазы гена mcsA описанного выше пути продукция отдельных поликетидов также ингибируется . Следовательно, утилизация пропионил-КоА через метилцитратный цикл снижает его концентрацию, одновременно увеличивая концентрацию поликетидов. Поликетид — это структура, обычно встречающаяся у грибов, состоящая из ацетил- и малонил -КоА, образующая продукт с чередующимися карбонильными и метиленовыми группами . Поликетиды и производные поликетидов часто имеют очень сложную структуру, а некоторые из них очень токсичны. Это привело к исследованиям по ограничению токсичности поликетидов для сельскохозяйственных культур через фитопатогенные грибы . [14]
Пропионилирование белков
[ редактировать ]Пропионил-КоА также является субстратом для посттрансляционной модификации белков путем реакции с остатками лизина на белках, реакция, называемая пропионилированием белка . [15] [16] Считается, что из-за структурного сходства ацетил-КоА и пропионил-КоА в реакции пропионилирования используются многие из тех же ферментов, которые используются для ацетилирования белков. [16] Хотя функциональные последствия пропионилирования белка в настоящее время до конца не изучены, пропионилирование in vitro фермента пропионил-КоА-синтетазы контролирует его активность. [17]
Человеческое и клиническое значение
[ редактировать ]
Gen5
[ редактировать ]Подобно тому, как пероксисомальные ферменты растений связывают пропионил-КоА и изобутирил-КоА, Gen5, ацетилтрансфераза у человека, связывается с пропионил-КоА и бутирил-КоА . Они специфически связываются с каталитическим доменом Gen5L2 . Эта консервативная ацетилтрансфераза отвечает за регуляцию транскрипции путем лизина ацетилирования гистонов хвостов N-концевых . Эта функция ацетилирования имеет гораздо более высокую скорость реакции, чем пропионилирование или бутирилирование . Из-за структуры пропионил-КоА Gen5 различает разные молекулы ацил-КоА . Фактически было обнаружено, что пропильная группа бутирл-КоА не может связываться из-за отсутствия стереоспецифичности с активным сайтом связывания Gen5 из-за ненасыщенных ацильных цепей . С другой стороны, третий атом углерода пропионил-КоА может вписаться в активный центр Gen5 с правильной ориентацией. [18]
Пропионовая ацидемия
[ редактировать ]На новорожденного этапах развития пропионовая ацидемия , которая представляет собой медицинскую проблему, определяемую как недостаток пропионил-КоА-карбоксилазы, может вызывать нарушения, умственную отсталость и множество других проблем. Это вызвано накоплением пропионил-КоА, поскольку он не может быть преобразован в метилмалонил-КоА . Новорожденных проверяют на повышенный уровень пропионилкарнитина . Дальнейшие способы диагностики этого заболевания включают анализ мочи. Используемые лекарства помогают обратить вспять и предотвратить повторение симптомов, включая использование добавок для снижения выработки пропионата . [5]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Дасгупта А (01.01.2019). «Глава 2 - Биотин: фармакология, патофизиология и оценка статуса биотина». В Дасгупте А (ред.). Биотин и другие помехи в иммуноанализах . Эльзевир. стр. 17–35. дои : 10.1016/B978-0-12-816429-7.00002-2 . ISBN 9780128164297 .
- ^ Перейти обратно: а б с Вонгкиттихоте П., Ах Мью Н., Чепмен К.А. (декабрь 2017 г.). «Пропионил-КоА-карбоксилаза - обзор» . Молекулярная генетика и обмен веществ . 122 (4): 145–152. дои : 10.1016/j.ymgme.2017.10.002 . ПМЦ 5725275 . ПМИД 29033250 .
- ^ Перейти обратно: а б Аптон AM, МакКинни JD (декабрь 2007 г.). «Роль метилцитратного цикла в метаболизме пропионата и детоксикации у Mycobacterium smegmatis» . Микробиология . 153 (Часть 12): 3973–82. дои : 10.1099/mic.0.2007/011726-0 . ПМИД 18048912 .
- ^ Перейти обратно: а б Долан С.К., Виджая А., Геддис С.М., Спринг Д.Р., Сильва-Роча Р., Уэлч М. (март 2018 г.). «Любить яд: метилцитратный цикл и бактериальный патогенез» . Микробиология . 164 (3): 251–259. дои : 10.1099/mic.0.000604 . ПМИД 29458664 .
- ^ Перейти обратно: а б Щелочков О.А., Каррильо Н., Вендитти С. (1993). «Пропионовая ацидемия» . Адам М.П., Ардингер Х.Х., Пагон Р.А., Уоллес С.Е., Бин Л.Дж., Стивенс К., Амемия А. (ред.). GeneReviews® . Вашингтонский университет, Сиэтл. ПМИД 22593918 . Проверено 13 июня 2019 г.
- ^ Сульд Х.М., Стейпл Э., Гурин С. (февраль 1962 г.). «Механизм образования желчных кислот из холестерина: окисление 5бита-холестана-3альфа,7альфа,12альфа-триола и образование пропионовой кислоты из боковой цепи митохондриями печени крысы» (PDF) . Журнал биологической химии . 237 (2): 338–44. дои : 10.1016/S0021-9258(18)93925-6 . ПМИД 13918291 .
- ^ Перейти обратно: а б с Луо Х, Чжоу Д, Лю Х, Не З, Кирога-Санчес ДЛ, Чанг Ю (26 мая 2016 г.). «Производство 3-гидроксипропионовой кислоты по пути пропионил-КоА с использованием рекомбинантных штаммов Escherichia coli» . ПЛОС ОДИН . 11 (5): e0156286. Бибкод : 2016PLoSO..1156286L . дои : 10.1371/journal.pone.0156286 . ПМК 4882031 . ПМИД 27227837 .
- ^ Халарнкар П.П., Бломквист Г.Дж. (1 января 1989 г.). «Сравнительные аспекты метаболизма пропионата». Сравнительная биохимия и физиология. Б. Сравнительная биохимия . 92 (2): 227–31. дои : 10.1016/0305-0491(89)90270-8 . ПМИД 2647392 .
- ^ Лю ВБ, Лю XX, Шен MJ, Ше ГЛ, Е BC (апрель 2019 г.). «Регулятор азота GlnR напрямую контролирует транскрипцию оперона prpDBC, участвующего в метилцитратном цикле у Mycobacterium smegmatis» . Журнал бактериологии . 201 (8). дои : 10.1128/JB.00099-19 . ПМЦ 6436344 . ПМИД 30745367 .
- ^ Райан Д.Г., Мерфи, член парламента, Фрезза С., Праг Х.А., Чучани Э.Т., О'Нил Л.А., Миллс Э.Л. (январь 2019 г.). «Связь метаболитов цикла Кребса с передачей сигналов при иммунитете и раке» . Природный метаболизм . 1 (1): 16–33. дои : 10.1038/s42255-018-0014-7 . ПМЦ 6485344 . ПМИД 31032474 .
- ^ Муньос-Элиас Э.Дж., Аптон А.М., Чериан Дж., МакКинни Дж.Д. (июнь 2006 г.). «Роль метилцитратного цикла в метаболизме, внутриклеточном росте и вирулентности микобактерий туберкулеза». Молекулярная микробиология . 60 (5): 1109–22. дои : 10.1111/j.1365-2958.2006.05155.x . ПМИД 16689789 .
- ^ Хан Дж., Хоу Дж., Чжан Ф., Ай Г., Ли М., Цай С. и др. (май 2013 г.). «Множественные пути снабжения пропионил-коэнзима А для производства биопластического поли(3-гидроксибутират-ко-3-гидроксивалерата) в Haloferax mediterranei» . Прикладная и экологическая микробиология . 79 (9): 2922–31. Бибкод : 2013ApEnM..79.2922H . дои : 10.1128/АЕМ.03915-12 . ПМЦ 3623125 . ПМИД 23435886 .
- ^ Лукас К.А., Филли-младший, Эрб Дж.М., Грейбилл Э.Р., Хоуз Дж.В. (август 2007 г.). «Пероксисомальный метаболизм пропионовой и изомасляной кислот в растениях» . Журнал биологической химии . 282 (34): 24980–9. дои : 10.1074/jbc.M701028200 . ПМИД 17580301 .
- ^ Чжан Ю.К., Брок М., Келлер Н.П. (октябрь 2004 г.). «Связь метаболизма пропионил-КоА с биосинтезом поликетидов у Aspergillus nidulans» . Генетика . 168 (2): 785–94. doi : 10.1534/genetics.104.027540 . ПМЦ 1448837 . ПМИД 15514053 .
- ^ Чен Ю., Спрунг Р., Тан Ю., Болл Х., Санграс Б., Ким С.К. и др. (май 2007 г.). «Пропионилирование и бутирилирование лизина являются новыми посттрансляционными модификациями гистонов» . Молекулярная и клеточная протеомика . 6 (5): 812–9. дои : 10.1074/mcp.M700021-MCP200 . ПМК 2911958 . ПМИД 17267393 .
- ^ Перейти обратно: а б Ченг З, Тан Ю, Чен Ю, Ким С, Лю Х, Ли СС и др. (январь 2009 г.). «Молекулярная характеристика пропионилизинов в негистоновых белках» . Молекулярная и клеточная протеомика . 8 (1): 45–52. дои : 10.1074/mcp.M800224-MCP200 . ПМК 2621001 . ПМИД 18753126 .
- ^ Гаррити Дж., Гарднер Дж.Г., Хоуз В., Вольбергер С., Эскаланте-Семерена Дж.К. (октябрь 2007 г.). «Пропионилирование N-лизина контролирует активность пропионил-КоА-синтетазы» . Журнал биологической химии . 282 (41): 30239–45. дои : 10.1074/jbc.m704409200 . ПМИД 17684016 .
- ^ Рингель А.Е., Вольбергер С. (июль 2016 г.). «Структурная основа дискриминации ацильной группы человеческим Gcn5L2» . Acta Crystallographica Раздел D. 72 (Часть 7): 841–8. Бибкод : 2016AcCrD..72..841R . дои : 10.1107/S2059798316007907 . ПМЦ 4932917 . ПМИД 27377381 .