Бактерии
| Бактерии | |
|---|---|
 | |
| Сканирующая электронная микрофотография палочек Escherichia coli . | |
| Научная классификация | |
| Домен: | Бактерии Вёзе и др. 1990 год |
| Фила | |
См . § Фила. | |
| Синонимы | |
| |
Бактерии ( / b æ k ˈ t ɪər i ə / ; сг. :бактерия) — повсеместно распространённые, преимущественно свободноживущие организмы, часто состоящие из одной биологической клетки . обширную область прокариотических микроорганизмов . Они составляют несколько микрометров Бактерии, обычно длиной , были одними из первых форм жизни, появившихся на Земле , и присутствуют в большинстве ее сред обитания . Бактерии обитают в почве, воде, кислых горячих источниках , радиоактивных отходах и глубокой биосфере земной коры . Бактерии играют жизненно важную роль на многих стадиях круговорота питательных веществ , перерабатывая питательные вещества и фиксируя азот из атмосферы . Круговорот питательных веществ включает разложение мертвых тел ; Бактерии ответственны за стадию гниения в этом процессе. В биологических сообществах, окружающих гидротермальные источники и холодные выходы , экстремофильные бактерии обеспечивают питательные вещества, необходимые для поддержания жизни, преобразуя растворенные соединения, такие как сероводород и метан , в энергию. Бактерии также живут в мутуалистических , комменсальных и паразитических отношениях с растениями и животными. Большинство бактерий не охарактеризовано, и существует множество видов, которые невозможно охарактеризовать. выращенный в лаборатории. Изучение бактерий известно как бактериология , раздел микробиологии .
Как и все животные, люди переносят огромное количество (около 10 13 до 10 14 ) бактерий. [2] Большинство из них находится в кишечнике , хотя их много и на коже. Большинство бактерий внутри и на теле безвредны или становятся таковыми в результате защитного воздействия иммунной системы , а многие из них полезны . [3] особенно те, что в кишечнике. Однако некоторые виды бактерий являются патогенными и вызывают инфекционные заболевания , в том числе холеру , сифилис , сибирскую язву , проказу , туберкулез , столбняк и бубонную чуму . Наиболее распространенными смертельными бактериальными заболеваниями являются респираторные инфекции . Антибиотики используются для лечения бактериальных инфекций , а также в сельском хозяйстве, что делает устойчивость к антибиотикам растущей проблемой. Бактерии играют важную роль в очистке сточных вод и ликвидации разливов нефти , производстве сыра и йогурта посредством ферментации , извлечении золота, палладия , меди и других металлов в горнодобывающем секторе, а также в биотехнологии и производстве антибиотиков и другие химикаты.
Когда-то бактерии считались растениями , входящими в класс Schizomycetes («грибы деления»), а теперь относят к прокариотам . В отличие от клеток животных и других эукариот , бактериальные клетки не содержат ядра и редко мембраносвязанные содержат органеллы . Хотя термин «бактерии» традиционно включал в себя всех прокариотов, научная классификация изменилась после открытия в 1990-х годах того, что прокариоты состоят из двух очень разных групп организмов, которые произошли от одного древнего общего предка . Эти эволюционные домены называются Бактерии и Археи . [4]
Этимология

Слово бактерии» — это множественное число от неолатинского слова бактерия , которое является латинизацией древнегреческого « βακτήριον ( baktḗrion ), [5] уменьшительное от бактерия baktēría ( ), что означает «посох, трость», [6] потому что первые из них, которые были обнаружены, имели форму стержня . [7] [8]
Происхождение и ранняя эволюция

Предками бактерий были одноклеточные микроорганизмы, которые были первыми формами жизни , появившимися на Земле около 4 миллиардов лет назад. [10] На протяжении примерно 3 миллиардов лет большинство организмов были микроскопическими, а доминирующими формами жизни были бактерии и археи. [11] [12] [13] Хотя бактериальные окаменелости существуют, такие как строматолиты , отсутствие у них отличительной морфологии не позволяет использовать их для изучения истории эволюции бактерий или для определения времени происхождения определенного вида бактерий. Однако последовательности генов могут быть использованы для реконструкции бактериальной филогении , и эти исследования показывают, что бактерии сначала отделились от архейной/эукариотической линии. [14] Самый недавний общий предок (MRCA) бактерий и архей, вероятно, был гипертермофилом , жившим около 2,5–3,2 миллиарда лет назад. [15] [16] [17] Самой ранней жизнью на суше, возможно, были бактерии около 3,22 миллиарда лет назад. [18]
Бактерии также участвовали во втором великом эволюционном расхождении – археях и эукариотах. [19] [20] Здесь эукариоты возникли в результате вступления древних бактерий в эндосимбиотические ассоциации с предками эукариотических клеток, которые сами, возможно, были родственниками архей. [21] [22] протоэукариотическими клетками Это включало поглощение альфа-протеобактериальных симбионтов с образованием либо митохондрий , либо гидрогеносом , которые до сих пор встречаются у всех известных эукариот (иногда в сильно редуцированной форме , например, у древних «амитохондриальных» простейших). Позже некоторые эукариоты, уже содержащие митохондрии, также поглотили цианобактерии -подобные организмы, что привело к образованию хлоропластов в водорослях и растениях. Это известно как первичный эндосимбиоз . [23]
среда обитания
Бактерии распространены повсеместно: они живут во всех возможных средах обитания на планете, включая почву, под водой, глубоко в земной коре и даже в таких экстремальных средах, как кислые горячие источники и радиоактивные отходы. [24] [25] Считается, что их примерно 2×10. 30 бактерии на Земле, [26] образуя биомассу , которую превосходят только растения. [27] Их много в озерах и океанах, арктических льдах и геотермальных источниках. [28] где они обеспечивают питательные вещества, необходимые для поддержания жизни, путем преобразования растворенных соединений, таких как сероводород и метан , в энергию. [29] Они живут на растениях и животных. Большинство из них не вызывают заболеваний, полезны для окружающей среды и необходимы для жизни. [3] [30] Почва является богатым источником бактерий, и в нескольких граммах их содержится около тысячи миллионов. Все они необходимы для экологии почвы, расщепления токсичных отходов и переработки питательных веществ. Они встречаются даже в атмосфере, а в одном кубическом метре воздуха содержится около ста миллионов бактериальных клеток. Океаны и моря имеют гавани примерно 3 х 10 26 бактерии, которые обеспечивают до 50% кислорода, которым дышит человек. [31] Полностью изучено лишь около 2% видов бактерий. [32]
| среда обитания | Разновидность | Ссылка |
|---|---|---|
| Холод (минус 15 °C Антарктида) | Криптоэндолиты | [33] |
| 70–100 °C Горячие ( гейзеры ) | Термус водный | [32] |
| Радиация, 5М Рад | Дейнококк радиодуранс | [33] |
| Соленый раствор, 47% соли ( Мертвое море , Большое Соленое озеро ) | несколько видов | [32] [33] |
| Кислота pH 3 | несколько видов | [24] |
| Щелочная pH 12,8 | бетапротеобактерии | [33] |
| Космос (6 лет на спутнике НАСА ) | Бацилла субтилис | [33] |
| 3,2 км под землей | несколько видов | [33] |
| Высокое давление ( Марианская впадина – 1200 атм .) | Морителла, Шеванелла и другие. | [33] |
Морфология
Размер . Бактерии имеют большое разнообразие форм и размеров. Бактериальные клетки составляют примерно одну десятую размера эукариотических клеток и обычно имеют 0,5–5,0 микрометров длину . Однако некоторые виды видны невооруженным глазом — например, Thiomargarita namibiensis имеет длину до полмиллиметра, [34] Epulopiscium fishelsoni достигает 0,7 мм, [35] и Thiomargarita magnifica могут достигать даже 2 см в длину, что в 50 раз больше, чем у других известных бактерий. [36] [37] Среди самых маленьких бактерий есть представители рода Mycoplasma , размер которых составляет всего 0,3 микрометра, что соответствует размеру самых крупных вирусов . [38] Некоторые бактерии могут быть еще меньше, но эти ультрамикробактерии недостаточно изучены. [39]
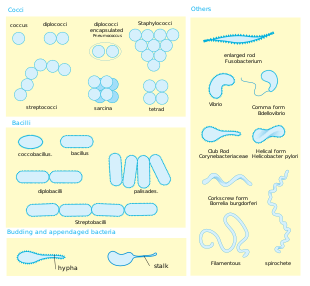
Форма . Большинство видов бактерий имеют либо сферическую форму, называемую кокками ( единственное число coccus , от греческого kókkos , зерно, семя), либо палочковидную форму, называемую бациллой ( sing . bacillus, от латинского baculus , палочка). [40] Некоторые бактерии, называемые вибрионами , имеют форму слегка изогнутых палочек или запятых; другие могут иметь спиралевидную форму, называемые спириллами , или плотно свернутые, называемые спирохетами . Описано небольшое количество других необычных форм, например, звездчатых бактерий. [41] Такое широкое разнообразие форм определяется клеточной стенкой и цитоскелетом бактерий и важно, поскольку оно может влиять на способность бактерий приобретать питательные вещества, прикрепляться к поверхностям, плавать в жидкостях и спасаться от хищников . [42] [43]

Многоклеточность . Большинство видов бактерий существуют в виде отдельных клеток; другие объединяются по характерным закономерностям: Neisseria образует диплоиды (пары), стрептококки образуют цепочки, а стафилококки группируются в гроздья «грозди винограда». Бактерии также могут группироваться, образуя более крупные многоклеточные структуры, такие как удлиненные нити видов Actinomycetota , агрегаты видов Myxobacteria и сложные гифы видов Streptomyces . [45] Эти многоклеточные структуры часто можно увидеть только в определенных условиях. Например, при недостатке аминокислот миксобактерии обнаруживают окружающие клетки в процессе, известном как ощущение кворума , мигрируют навстречу друг другу и объединяются, образуя плодовые тела длиной до 500 микрометров, содержащие примерно 100 000 бактериальных клеток. [46] В этих плодовых телах бактерии выполняют отдельные задачи; например, примерно одна из десяти клеток мигрирует в верхушку плодового тела и дифференцируется в специализированное спящее состояние, называемое миксоспорой, которое более устойчиво к высыханию и другим неблагоприятным условиям окружающей среды. [47]
Биопленки . Бактерии часто прикрепляются к поверхностям и образуют плотные скопления, называемые биопленками. [48] и более крупные образования, известные как микробные маты . [49] Эти биопленки и маты могут иметь толщину от нескольких микрометров до глубины до полуметра и могут содержать несколько видов бактерий, простейших и архей. Бактерии, живущие в биопленках, демонстрируют сложное расположение клеток и внеклеточных компонентов, образуя вторичные структуры, такие как микроколонии , через которые проходят сети каналов, обеспечивающие лучшую диффузию питательных веществ. [50] [51] В естественной среде, такой как почва или поверхность растений, большинство бактерий связаны с поверхностями в виде биопленок. [52] Биопленки также важны в медицине, поскольку эти структуры часто присутствуют во время хронических бактериальных инфекций или инфекций имплантированных медицинских устройств , а бактерии, защищенные биопленками, гораздо труднее убить, чем отдельные изолированные бактерии. [53]
Клеточная структура

Внутриклеточные структуры
Бактериальная клетка окружена клеточной мембраной , состоящей преимущественно из фосфолипидов . Эта мембрана закрывает содержимое клетки и действует как барьер для удержания питательных веществ, белков и других важных компонентов цитоплазмы внутри клетки. [54] В отличие от эукариотических клеток , у бактерий обычно отсутствуют крупные мембраносвязанные структуры в цитоплазме, такие как ядро , митохондрии , хлоропласты и другие органеллы, присутствующие в эукариотических клетках. [55] Однако у некоторых бактерий в цитоплазме есть связанные с белками органеллы, которые разделяют аспекты бактериального метаболизма. [56] [57] например, карбоксисома . [58] Кроме того, бактерии имеют многокомпонентный цитоскелет , позволяющий контролировать локализацию белков и нуклеиновых кислот внутри клетки и управлять процессом клеточного деления . [59] [60] [61]
Многие важные биохимические реакции, такие как выработка энергии, происходят из-за градиентов концентрации на мембранах, создавая разность потенциалов, аналогичную батарее. Общее отсутствие внутренних мембран у бактерий означает, что эти реакции, такие как транспорт электронов , происходят через клеточную мембрану между цитоплазмой и внешней частью клетки или периплазмой . [62] Однако у многих фотосинтезирующих бактерий плазматическая мембрана сильно складчата и заполняет большую часть клетки слоями светособирающей мембраны. [63] Эти светособирающие комплексы могут даже образовывать заключенные в липиды структуры, называемые хлоросомами у зеленых серных бактерий . [64]
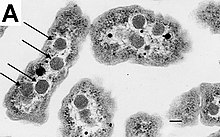
Бактерии не имеют мембраносвязанного ядра, и их генетический материал обычно представляет собой одну кольцевую бактериальную хромосому , ДНК расположенную в цитоплазме в теле неправильной формы, называемом нуклеоидом . [65] Нуклеоид содержит хромосому со связанными с ней белками и РНК . Как и все другие организмы , бактерии содержат рибосомы для производства белков, но структура бактериальной рибосомы отличается от структуры эукариот и архей. [66]
Некоторые бактерии производят внутриклеточные гранулы для хранения питательных веществ, таких как гликоген , [67] полифосфат , [68] сера [69] или полигидроксиалканоаты . [70] Бактерии, такие как фотосинтезирующие цианобактерии , производят внутренние газовые вакуоли , которые они используют для регулирования своей плавучести, позволяя им перемещаться вверх или вниз в слои воды с различной интенсивностью света и уровнями питательных веществ. [71]
Внеклеточные структуры
Вокруг клеточной мембраны снаружи расположена клеточная стенка . Стенки бактериальных клеток состоят из пептидогликана (также называемого муреином), который состоит из полисахаридных цепей, сшитых пептидами, содержащими D- аминокислоты . [72] Стенки бактериальных клеток отличаются от клеточных стенок растений и грибов , которые состоят из целлюлозы и хитина соответственно. [73] Клеточная стенка бактерий также отличается от клеточной стенки ахей, которые не содержат пептидогликана. Клеточная стенка необходима для выживания многих бактерий, а антибиотик пенициллин (продуцируемый грибом Penicillium ) способен убивать бактерии, ингибируя этап синтеза пептидогликана. [73]
Вообще говоря, у бактерий есть два разных типа клеточной стенки, которые делят бактерии на грамположительные и грамотрицательные бактерии . Названия происходят от реакции клеток на окраску по Граму — давний тест для классификации видов бактерий. [74]
Грамположительные бактерии обладают толстой клеточной стенкой, содержащей множество слоев пептидогликана и тейхоевых кислот . Напротив, грамотрицательные бактерии имеют относительно тонкую клеточную стенку, состоящую из нескольких слоев пептидогликана, окруженных второй липидной мембраной, содержащей липополисахариды и липопротеины . Большинство бактерий имеют грамотрицательную клеточную стенку, и только представители группы Bacillota и actinomycetota (ранее известные как грамположительные бактерии с низким G + C и высоким G + C соответственно) имеют альтернативное грамположительное расположение. [75] Эти различия в структуре могут привести к различиям в чувствительности к антибиотикам; например, ванкомицин может убивать только грамположительные бактерии и неэффективен против грамотрицательных патогенов , таких как Haemophilus influenzae или Pseudomonas aeruginosa . [76] Некоторые бактерии имеют структуры клеточной стенки, которые не являются ни классическими грамположительными, ни грамотрицательными. Сюда входят клинически важные бактерии, такие как микобактерии , которые имеют толстую клеточную стенку из пептидогликана, как у грамположительных бактерий, а также второй внешний слой липидов. [77]
У многих бактерий S-слой жестко расположенных белковых молекул покрывает внешнюю поверхность клетки. [78] Этот слой обеспечивает химическую и физическую защиту поверхности клетки и может действовать как макромолекулярный диффузионный барьер . S-слои имеют разнообразные функции и, как известно, действуют как факторы вирулентности у видов Campylobacter и содержат поверхностные ферменты у Bacillus stearothermophilus . [79] [80]
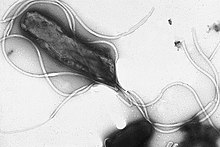
Жгутики представляют собой жесткие белковые структуры диаметром около 20 нанометров и длиной до 20 микрометров, которые используются для подвижности . Жгутики приводятся в движение энергией, выделяемой при переносе ионов по электрохимическому градиенту через клеточную мембрану. [81]
Фимбрии (иногда называемые « прикрепительными пилями ») представляют собой тонкие белковые нити, обычно диаметром 2–10 нанометров и длиной до нескольких микрометров. Они распределены по поверхности клетки и под электронным микроскопом напоминают тонкие волоски . [82] Считается, что фимбрии участвуют в прикреплении к твердым поверхностям или к другим клеткам и необходимы для вирулентности некоторых бактериальных патогенов. [83] Пили ( sing . pilus) — это клеточные придатки, немного крупнее фимбрии, которые могут переносить генетический материал между бактериальными клетками в процессе, называемом конъюгацией , где они называются конъюгационными пилями или половыми пилями (см. Бактериальную генетику ниже). [84] Они также могут генерировать движение, где их называют пилями IV типа . [85]
Гликокаликс вырабатывается многими бактериями и окружает их клетки. [86] и различается по структурной сложности: от неорганизованного слоя слизи внеклеточных полимерных веществ до высокоструктурированной капсулы . Эти структуры могут защищать клетки от поглощения эукариотическими клетками, такими как макрофаги (часть иммунной системы человека ). [87] Они также могут действовать как антигены и участвовать в распознавании клеток, а также способствовать прикреплению к поверхностям и образованию биопленок. [88]
Сборка этих внеклеточных структур зависит от систем секреции бактерий . Они переносят белки из цитоплазмы в периплазму или в среду вокруг клетки. Известны многие типы систем секреции, и эти структуры часто необходимы для вирулентности патогенов, поэтому они интенсивно изучаются. [88]
Эндоспоры
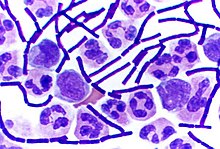
Некоторые роды грамположительных бактерий, такие как Bacillus , Clostridium , Sporohalobacter , Anaerobacter и Heliobacterium , могут образовывать высокорезистентные, спящие структуры, называемые эндоспорами . [90] Эндоспоры развиваются в цитоплазме клетки; обычно в каждой клетке развивается одна эндоспора. [91] Каждая эндоспора содержит ядро из ДНК и рибосом, окруженное слоем коры и защищенное многослойной жесткой оболочкой, состоящей из пептидогликана и различных белков. [91]
Эндоспоры не проявляют заметного метаболизма и могут выдерживать экстремальные физические и химические стрессы, такие как высокие уровни ультрафиолетового излучения , гамма-излучения , моющих средств , дезинфицирующих средств , тепла, замораживания, давления и высыхания . [92] В этом состоянии покоя эти организмы могут оставаться жизнеспособными в течение миллионов лет. [93] [94] [95] Эндоспоры даже позволяют бактериям выживать в условиях вакуума и радиации космического пространства , что приводит к возможности распространения бактерий по Вселенной с помощью космической пыли , метеороидов , астероидов , комет , планетоидов или направленной панспермии . [96] [97]
Бактерии, образующие эндоспоры, могут вызывать заболевания; например, сибирской язвой можно заразиться при вдыхании эндоспор Bacillus anthracis , а заражение глубоких колотых ран эндоспорами Clostridium tetani вызывает столбняк , который, как и ботулизм , вызывается токсином, выделяемым бактериями, растущими из спор. [98] Clostridioides difficile Инфекция , распространенная проблема в медицинских учреждениях, вызывается спорообразующими бактериями. [99]
Метаболизм
Бактерии демонстрируют чрезвычайно широкое разнообразие типов метаболизма . [100] Распределение метаболических признаков внутри группы бактерий традиционно использовалось для определения их таксономии , но эти признаки часто не соответствуют современным генетическим классификациям. [101] Бактериальный метаболизм классифицируется на группы питания на основе трех основных критериев: источник энергии , используемые доноры электронов и источник углерода , используемый для роста. [102]
Фототрофные бактерии получают энергию из света с помощью фотосинтеза , тогда как хемотрофные бактерии расщепляют химические соединения путем окисления . [103] управление метаболизмом путем переноса электронов от данного донора электронов к терминальному акцептору электронов в ходе окислительно-восстановительной реакции . Хемотрофы далее делятся по типам соединений, которые они используют для переноса электронов. Бактерии, которые получают электроны из неорганических соединений, таких как водород, окись углерода или аммиак , называются литотрофами , а те, которые используют органические соединения, называются органотрофами . [103] Тем не менее, более конкретно, аэробные организмы используют кислород в качестве терминального акцептора электронов, в то время как анаэробные организмы используют другие соединения, такие как нитрат , сульфат или углекислый газ. [103]
Многие бактерии, называемые гетеротрофами , получают углерод из другого органического углерода . Другие, такие как цианобактерии и некоторые пурпурные бактерии , являются автотрофами , то есть они получают клеточный углерод путем фиксации углекислого газа . [104] В необычных обстоятельствах газообразный метан может использоваться метанотрофными бактериями как источник электронов и субстрат для углеродного анаболизма . [105]
| Тип питания | Источник энергии | Источник углерода | Примеры |
|---|---|---|---|
| Фототрофы | Солнечный свет | Органические соединения (фотогетеротрофы) или фиксация углерода (фотоавтотрофы) | Цианобактерии , зеленые серобактерии , хлорофлексота или пурпурные бактерии. |
| Литотрофы | Неорганические соединения | Органические соединения (литогетеротрофы) или фиксация углерода (литоавтотрофы) | Thermodesulfobacteriota , Hydrogenophilaceae или Nitrospirota. |
| Органотрофы | Органические соединения | Органические соединения (хемогетеротрофы) или фиксация углерода (хемоавтотрофы) | Bacillus , Clostridium или Enterobacteriaceae |
Во многих отношениях бактериальный метаболизм обеспечивает свойства, полезные для экологической стабильности и человеческого общества. Например, диазотрофы обладают способностью фиксировать газообразный азот с помощью фермента нитрогеназы . [106] Эта особенность, присущая бактериям большинства перечисленных выше метаболических типов, [107] приводит к экологически важным процессам денитрификации , сульфатредукции и ацетогенеза соответственно. [108] Бактериальные метаболические процессы являются важными движущими силами биологических реакций на загрязнение ; например, сульфатвосстанавливающие бактерии в значительной степени ответственны за производство высокотоксичных форм ртути ( метил- и диметилртути ) в окружающей среде. [109] Нереспираторные анаэробы используют ферментацию для выработки энергии и снижения мощности, выделяя побочные продукты метаболизма (например, этанол в пивоварении) в качестве отходов. Факультативные анаэробы могут переключаться между ферментацией и различными акцепторами терминальных электронов в зависимости от условий окружающей среды, в которых они находятся. [110]
Рост и размножение



В отличие от многоклеточных организмов, увеличение размера клеток ( рост клеток ) и размножение путем деления клеток у одноклеточных организмов тесно связаны. Бактерии вырастают до фиксированного размера, а затем размножаются посредством бинарного деления , формы бесполого размножения . [112] В оптимальных условиях бактерии могут расти и делиться чрезвычайно быстро, а некоторые популяции бактерий могут удваиваться каждые 17 минут. [113] При делении клеток два идентичных клона образуются дочерних клеток. Некоторые бактерии, продолжая размножаться бесполым путем, образуют более сложные репродуктивные структуры, которые помогают рассеять вновь образовавшиеся дочерние клетки. Примеры включают образование плодовых тел миксобактериями и воздушных гиф образование видами Streptomyces или почкование. Почкование предполагает, что клетка образует выступ, который отрывается и производит дочернюю клетку. [114]
В лаборатории бактерии обычно выращивают на твердых или жидких средах. [115] Твердые питательные среды , такие как чашки с агаром , используются для выделения чистых культур бактериального штамма. Однако жидкие питательные среды используются, когда требуется измерение роста или больших объемов клеток. Рост в перемешиваемых жидких средах происходит в виде равномерной клеточной суспензии, что позволяет легко делить и переносить культуры, хотя изолировать отдельные бактерии из жидких сред сложно. Использование селективных сред (сред с добавлением или недостатком определенных питательных веществ или с добавлением антибиотиков) может помочь идентифицировать конкретные организмы. [116]
Большинство лабораторных методов выращивания бактерий используют высокие уровни питательных веществ для дешевого и быстрого производства большого количества клеток. [115] Однако в естественной среде количество питательных веществ ограничено, а это означает, что бактерии не могут размножаться бесконечно. Это ограничение питательных веществ привело к развитию различных стратегий роста (см. теорию отбора r/K ). Некоторые организмы могут расти чрезвычайно быстро, когда питательные вещества становятся доступными, например, образование цветения водорослей и цианобактерий , которое часто происходит в озерах летом. [117] Другие организмы адаптируются к суровым условиям окружающей среды, например, стрептомицеты производят множество антибиотиков , которые подавляют рост конкурирующих микроорганизмов. [118] В природе многие организмы живут сообществами (например, биопленками ), что может обеспечить увеличение снабжения питательными веществами и защиту от стрессов окружающей среды. [52] Эти взаимоотношения могут иметь существенное значение для роста конкретного организма или группы организмов ( синтрофия ). [119]
Рост бактерий проходит четыре фазы. Когда популяция бактерий впервые попадает в среду с высоким содержанием питательных веществ, обеспечивающую рост, клеткам необходимо адаптироваться к новой среде. Первая фаза роста — это лаг-фаза , период медленного роста, когда клетки адаптируются к среде с высоким содержанием питательных веществ и готовятся к быстрому росту. Лаг-фаза имеет высокие темпы биосинтеза, поскольку вырабатываются белки, необходимые для быстрого роста. [120] [121] Вторая фаза роста — логарифмическая фаза , также известная как экспоненциальная фаза. Лог-фаза характеризуется быстрым экспоненциальным ростом . Скорость, с которой клетки растут во время этой фазы, известна как скорость роста ( k ), а время, необходимое клеткам для удвоения, известно как время генерации ( g ). Во время лог-фазы питательные вещества метаболизируются с максимальной скоростью, пока одно из питательных веществ не истощается и не начинает ограничивать рост. Третья фаза роста является стационарной фазой и вызвана истощением питательных веществ. Клетки снижают свою метаболическую активность и потребляют несущественные клеточные белки. Стационарная фаза представляет собой переход от быстрого роста к состоянию реакции на стресс, при этом увеличивается экспрессия генов, участвующих в репарации ДНК , антиоксидантном метаболизме и транспорте питательных веществ . [122] Заключительная фаза — это фаза смерти , когда у бактерий заканчиваются питательные вещества и они умирают. [123]
Генетика
Большинство бактерий имеют одну кольцевую хромосому , размер которой может варьироваться от 160 000 пар оснований у эндосимбиотических бактерий Carsonella ruddii . [125] до 12 200 000 пар оснований (12,2 МБП) у почвенных бактерий Sorangium cellulosum . [126] Из этого есть много исключений; например, некоторые виды Streptomyces и Borrelia содержат одну линейную хромосому, [127] [128] в то время как некоторые виды вибрионов содержат более одной хромосомы. [129] Некоторые бактерии содержат плазмиды , небольшие внехромосомные молекулы ДНК, которые могут содержать гены для различных полезных функций, таких как устойчивость к антибиотикам , метаболические способности или различные факторы вирулентности . [130]
Геномы бактерий обычно кодируют от нескольких сотен до нескольких тысяч генов. Гены в бактериальных геномах обычно представляют собой один непрерывный участок ДНК. Хотя у бактерий действительно существует несколько различных типов интронов , они встречаются гораздо реже, чем у эукариот. [131]
Бактерии, как бесполые организмы, наследуют идентичную копию родительского генома и являются клональными . Однако все бактерии могут развиваться путем отбора изменений в ДНК их генетического материала, вызванных генетической рекомбинацией или мутациями . Мутации возникают в результате ошибок, допущенных во время репликации ДНК или в результате воздействия мутагенов . Скорость мутаций широко варьируется среди разных видов бактерий и даже среди разных клонов одного вида бактерий. [132] Генетические изменения в бактериальных геномах возникают либо в результате случайной мутации во время репликации, либо в результате «мутации, направленной на стресс», когда гены, участвующие в определенном процессе, ограничивающем рост, имеют повышенную частоту мутаций. [133]
Некоторые бактерии переносят генетический материал между клетками. Это может произойти тремя основными способами. Во-первых, бактерии могут поглощать экзогенную ДНК из окружающей среды в процессе, называемом трансформацией . [134] Многие бактерии могут естественным образом поглощать ДНК из окружающей среды, в то время как другие должны быть химически изменены, чтобы заставить их поглощать ДНК. [135] Развитие компетентности в природе обычно связано со стрессовыми условиями окружающей среды и, по-видимому, является адаптацией, способствующей восстановлению повреждений ДНК в клетках-реципиентах. [136] Во-вторых, бактериофаги могут интегрироваться в бактериальную хромосому, вводя чужеродную ДНК в процессе, известном как трансдукция . Существует множество типов бактериофагов; некоторые заражают и лизируют бактерии - хозяева , а другие встраиваются в бактериальную хромосому. [137] Бактерии сопротивляются фаговой инфекции посредством систем рестрикционной модификации , которые разрушают чужеродную ДНК. [138] и систему, которая использует последовательности CRISPR для сохранения фрагментов геномов фагов, с которыми бактерии контактировали в прошлом, что позволяет им блокировать репликацию вируса посредством формы РНК-интерференции . [139] [140] В-третьих, бактерии могут передавать генетический материал посредством прямого контакта с клетками посредством конъюгации . [141]
В обычных обстоятельствах трансдукция, конъюгация и трансформация включают перенос ДНК между отдельными бактериями одного и того же вида, но иногда перенос может происходить между особями разных видов бактерий, и это может иметь серьезные последствия, такие как передача устойчивости к антибиотикам. [142] [143] В таких случаях приобретение генов от других бактерий или окружающей среды называется горизонтальным переносом генов и может быть обычным явлением в естественных условиях. [144]
Поведение
Движение

Многие бактерии подвижны (способны передвигаться самостоятельно) и делают это, используя различные механизмы. Наиболее изученными из них являются жгутики — длинные нити, которые вращаются двигателем у основания, создавая движение, подобное пропеллеру. [145] Бактериальный жгутик состоит примерно из 20 белков, еще примерно 30 белков необходимы для его регуляции и сборки. [145] Жгутик представляет собой вращающуюся структуру, приводимую в движение обратимым двигателем в основании, который использует электрохимический градиент на мембране для получения энергии. [146]

Бактерии могут использовать жгутики по-разному для создания разных видов движений. Многие бактерии (например, кишечная палочка ) имеют два различных режима движения: движение вперед (плавание) и кувыркание. Кувыркание позволяет им переориентироваться и превращает их движение в трехмерное случайное блуждание . [147] Виды бактерий различаются числом и расположением жгутиков на поверхности; у одних один жгутик ( монотрихозный ), по жгутику на каждом конце ( амфитрихозный ), скопления жгутиков у полюсов клетки ( лофотрихозный ), у других жгутики распределены по всей поверхности клетки ( перитрихозный ). Жгутики группы бактерий спирохет расположены между двумя мембранами в периплазматическом пространстве. У них характерное спиральное тело, которое вращается при движении. [145]
Два других типа бактериального движения называются подергивающейся подвижностью , которая основана на структуре, называемой пилусом IV типа . [148] и скользящая моторика , использующая другие механизмы. При подергивающейся подвижности палочковидный пилус выдвигается из клетки, связывает некоторый субстрат, а затем втягивается, тянуя клетку вперед. [149]
Подвижные бактерии привлекаются или отталкиваются определенными стимулами в поведении, называемом налогами : к ним относятся хемотаксис , фототаксис , энергетическое такси и магнитотаксис . [150] [151] [152] В одной своеобразной группе, миксобактериях, отдельные бактерии перемещаются вместе, образуя волны клеток, которые затем дифференцируются, образуя плодовые тела, содержащие споры. [47] Миксобактерии передвигаются только на твердых поверхностях, в отличие от кишечной палочки , которая подвижна в жидких или твердых средах. [153]
Некоторые виды Listeria и Shigella перемещаются внутри клеток-хозяев, узурпируя цитоскелет , который обычно используется для перемещения органелл внутри клетки. Способствуя актина полимеризации на одном полюсе своих клеток, они могут образовывать своего рода хвост, который продвигает их через цитоплазму клетки-хозяина. [154]
Коммуникация
У некоторых бактерий есть химические системы, генерирующие свет. Эта биолюминесценция часто возникает у бактерий, живущих вместе с рыбами, и свет, вероятно, служит для привлечения рыб или других крупных животных. [155]
Бактерии часто функционируют как многоклеточные агрегаты, известные как биопленки , обмениваясь различными молекулярными сигналами для межклеточной коммуникации и участвуя в скоординированном многоклеточном поведении. [156] [157]
Общие преимущества многоклеточного сотрудничества включают клеточное разделение труда , доступ к ресурсам, которые не могут эффективно использоваться отдельными клетками, коллективную защиту от антагонистов и оптимизацию выживания популяции за счет дифференцировки в отдельные типы клеток. [156] Например, бактерии в биопленках могут иметь более чем в пятьсот раз повышенную устойчивость к антибактериальным агентам, чем отдельные «планктонные» бактерии того же вида. [157]
Один из типов межклеточной коммуникации с помощью молекулярного сигнала называется «чувством кворума» , который служит для определения того, достаточна ли плотность местной популяции для поддержки инвестиций в процессы, которые успешны только в том случае, если большое количество подобных организмов ведут себя одинаково, например, выделяют пищеварительные ферменты. или излучающий свет. [158] [159] Чувство кворума позволяет бактериям координировать экспрессию генов , а также производить, высвобождать и обнаруживать аутоиндукторы или феромоны , которые накапливаются по мере роста популяции клеток. [160]
Классификация и идентификация


Классификация направлена на описание разнообразия видов бактерий путем наименования и группировки организмов на основе сходства. Бактерии можно классифицировать на основе клеточной структуры, клеточного метаболизма или на основе различий в клеточных компонентах, таких как ДНК , жирные кислоты , пигменты, антигены и хиноны . [116] Хотя эти схемы позволяли идентифицировать и классифицировать бактериальные штаммы, было неясно, представляют ли эти различия различия между отдельными видами или между штаммами одного и того же вида. Эта неопределенность была связана с отсутствием отличительных структур у большинства бактерий, а также с латеральным переносом генов между неродственными видами. [162] Из-за латерального переноса генов некоторые близкородственные бактерии могут иметь очень разную морфологию и метаболизм. Чтобы преодолеть эту неопределенность, современная классификация бактерий делает упор на молекулярную систематику с использованием генетических методов, таких как гуанина и цитозина определение соотношения , гибридизация геном-геном, а также секвенирование генов, которые не подверглись обширному латеральному переносу генов, таких как ген рРНК . [163] Классификация бактерий определяется публикацией в Международном журнале систематической бактериологии. [164] и Руководство Берджи по систематической бактериологии. [165] Международный комитет по систематической бактериологии (ICSB) поддерживает международные правила наименования бактерий и таксономических категорий, а также их ранжирования в Международном кодексе номенклатуры бактерий . [166]
Исторически бактерии считались частью Plantae , царства растений, и назывались «шизомицетами» (грибами деления). [167] По этой причине коллективные бактерии и другие микроорганизмы хозяина часто называют «флорой». [168] Термин «бактерии» традиционно применялся ко всем микроскопическим одноклеточным прокариотам. Однако молекулярная систематика показала, что прокариотическая жизнь состоит из двух отдельных доменов , первоначально называвшихся Eubacteria и Archaebacteria, но теперь называемых Bacteria и Archaea, которые развились независимо от древнего общего предка. [4] Археи и эукариоты более тесно связаны друг с другом, чем с бактериями. Эти два домена вместе с эукарией составляют основу трехдоменной системы , которая в настоящее время является наиболее широко используемой классификационной системой в микробиологии. [169] Однако из-за относительно недавнего внедрения молекулярной систематики и быстрого увеличения числа доступных последовательностей генома классификация бактерий остается меняющейся и расширяющейся областью. [170] [171] Например, Кавальер-Смит утверждал, что археи и эукариоты произошли от грамположительных бактерий. [172]
Идентификация бактерий в лаборатории особенно актуальна в медицине , где правильное лечение определяется видом бактерий, вызвавшим инфекцию. Следовательно, необходимость идентификации патогенов человека стала основным стимулом для разработки методов идентификации бактерий. [173]
Окраска по Граму , разработанная в 1884 году Гансом Христианом Грамом , характеризует бактерии на основе структурных характеристик их клеточных стенок. [174] [74] Толстые слои пептидогликана в стенке «грамположительных» клеток окрашиваются в фиолетовый цвет, а тонкие стенки «грамотрицательных» клеток кажутся розовыми. [174] Объединив морфологию и окраску по Граму, большинство бактерий можно отнести к одной из четырех групп (грамположительные кокки, грамположительные бациллы, грамотрицательные кокки и грамотрицательные бациллы). Некоторые организмы лучше всего идентифицируются с помощью красителей, отличных от окраски по Граму, особенно микобактерий или Nocardia , которые проявляют кислотоустойчивость при окраске по Цилю-Нильсену или подобных окрасках. [175] Другие организмы, возможно, придется идентифицировать по их росту в специальной среде или с помощью других методов, таких как серология . [176]
Методы культивирования предназначены для стимулирования роста и идентификации конкретных бактерий, одновременно ограничивая рост других бактерий в образце. [177] Часто эти методы предназначены для конкретных образцов; например, образец мокроты будет обработан для выявления микроорганизмов, вызывающих пневмонию , а образцы стула будут культивироваться на селективной среде для выявления организмов, вызывающих диарею, при этом предотвращая рост непатогенных бактерий. Обычно стерильные образцы, такие как кровь , моча или спинномозговая жидкость , культивируются в условиях, предназначенных для выращивания всех возможных организмов. [116] [178] После выделения патогенного организма его можно дополнительно охарактеризовать по морфологии, характеру роста (например, аэробный или анаэробный рост), характеру гемолиза и окрашиванию. [179]
Как и в случае с бактериальной классификацией, для идентификации бактерий все чаще используют молекулярные методы. [180] и масс-спектроскопия . [181] Большинство бактерий не охарактеризовано, и существует множество видов, которые невозможно выращивать в лаборатории. [182] Диагностика с использованием инструментов на основе ДНК, таких как полимеразная цепная реакция , становится все более популярной благодаря своей специфичности и скорости по сравнению с методами, основанными на культуре. [183] Эти методы также позволяют обнаруживать и идентифицировать « жизнеспособные, но некультивируемые » клетки, метаболически активные, но неделящиеся. [184] Однако даже при использовании этих усовершенствованных методов общее количество видов бактерий неизвестно и даже не может быть оценено с какой-либо уверенностью. Согласно современной классификации, известно немногим менее 9300 видов прокариот, включая бактерии и археи; [185] но попытки оценить истинное количество бактериального разнообразия варьировались от 10 7 до 10 9 общее количество видов — и даже эти различные оценки могут отличаться на многие порядки. [186] [187]
Фила
Следующие типы были официально опубликованы в соответствии с Бактериологическим кодексом : [188]
- Ацидобактериота
- Актиномицетота
- Аквификота
- Арматимонадота
- Атрибактериота
- Бациллота
- Бактероидота
- Бальнеолота
- Бделловибрионота
- Кальдисерикота
- Калдитрихота
- Кампилобактерия
- Хламидиота
- Хлоробиота
- Хлорофлексота
- Хризиогенота
- Копротермобактерия
- Деферрибактериота
- Дейнококкота
- Диктиогломота
- Элюзимикробиота
- Фибробактерита
- Фузобактериота
- Одаренный драгоценными камнями
- Игнавибактериота
- Лентисфаэрота
- Микоплазматота
- Миксококкота
- Нитроспинота
- Нитроспирота
- Планктомицетота
- Псевдомонадота
- Родотермота
- Спирохетата
- синергист
- Термодесульфобактерии
- Термомикробиота
- Термотогота
- Веррукомикробиота
Взаимодействие с другими организмами
Несмотря на кажущуюся простоту, бактерии могут образовывать сложные ассоциации с другими организмами. Эти симбиотические ассоциации можно разделить на паразитизм , мутуализм и комменсализм . [190]
Комменсалы
Слово « комменсализм » происходит от слова «commensal», что означает «есть за одним столом». [191] и все растения и животные колонизированы комменсальными бактериями. У людей и других животных миллионы из них обитают на коже, дыхательных путях, кишечнике и других отверстиях. [192] [193] Называется «нормальной флорой». [194] или «комменсалы», [195] эти бактерии обычно не причиняют вреда, но иногда могут проникать в другие участки тела и вызывать инфекцию. Escherichia coli является комменсалом в кишечнике человека, но может вызывать инфекции мочевыводящих путей. [196] Точно так же стрептококки, являющиеся частью нормальной флоры полости рта человека, могут вызывать болезни сердца . [197]
Хищники
Некоторые виды бактерий убивают, а затем поедают другие микроорганизмы; эти виды называются хищными бактериями . [198] К ним относятся такие организмы, как Myxococcus xanthus , который образует рои клеток , которые убивают и переваривают любые бактерии, с которыми они сталкиваются. [199] Другие бактериальные хищники либо прикрепляются к своей добыче, чтобы переварить ее и поглотить питательные вещества, либо проникают в другую клетку и размножаются внутри цитозоля. [200] Считается, что эти хищные бактерии произошли от сапрофагов , потреблявших мертвые микроорганизмы, путем адаптации, позволившей им захватывать и убивать другие организмы. [201]
Мутуалисты
Некоторые бактерии образуют тесные пространственные ассоциации, необходимые для их выживания. Одна из таких мутуалистических ассоциаций, называемая межвидовым переносом водорода, возникает между кластерами анаэробных бактерий , которые потребляют органические кислоты , такие как масляная или пропионовая кислота , и производят водород , и метаногенными архей, которые потребляют водород. [202] Бактерии в этой ассоциации не могут потреблять органические кислоты, поскольку в результате этой реакции образуется водород, который накапливается в их среде. Только тесная связь с археями, потребляющими водород, поддерживает концентрацию водорода на достаточно низком уровне, чтобы позволить бактериям расти. [203]
В почве микроорганизмы, обитающие в ризосфере (зоне, включающей поверхность корня и почву, которая прилипает к корню после легкого встряхивания), осуществляют фиксацию азота , превращая газообразный азот в азотистые соединения. [204] Это обеспечивает легкоусвояемую форму азота для многих растений, которые не могут самостоятельно фиксировать азот. Многие другие бактерии встречаются в качестве симбионтов у человека и других организмов. Например, присутствие более 1000 видов бактерий в нормальной кишечной флоре человека способствовать кишечному может иммунитету, синтезу витаминов , таких как фолиевая кислота , витамин К и биотин , превращению сахаров в молочную кислоту (см. Lactobacillus ), а также как ферментирующие сложные неперевариваемые углеводы . [205] [206] [207] Присутствие этой кишечной флоры также подавляет рост потенциально патогенных бактерий (обычно посредством конкурентного исключения ), и эти полезные бактерии впоследствии продаются в качестве пробиотических пищевых добавок . [208]
Выживание почти всех животных зависит от бактерий, поскольку только бактерии и некоторые археи обладают генами и ферментами, необходимыми для синтеза витамина B12 , также известного как кобаламин , и обеспечения его через пищевую цепь. Витамин B 12 – водорастворимый витамин , который участвует в обмене веществ каждой клетки человеческого организма. Он является кофактором синтеза ДНК , а также метаболизма жирных кислот и аминокислот . Он особенно важен для нормального функционирования нервной системы благодаря своей роли в синтезе миелина . [209]
Патогены

Организм постоянно подвергается воздействию многих видов бактерий, в том числе полезных комменсалов, которые растут на коже и слизистых оболочках , и сапрофитов , которые растут главным образом в почве и разлагающихся веществах. Кровь и тканевые жидкости содержат питательные вещества, достаточные для поддержания роста многих бактерий. Организм имеет защитные механизмы, которые позволяют ему противостоять микробному вторжению в его ткани и придают ему естественный иммунитет или врожденную устойчивость против многих микроорганизмов . [210] В отличие от некоторых вирусов , бактерии развиваются относительно медленно, поэтому многие бактериальные заболевания встречаются и у других животных. [211]
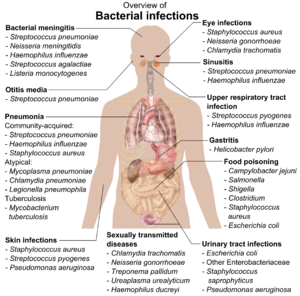
Если бактерии образуют паразитарную ассоциацию с другими организмами, их относят к патогенам. [212] Патогенные бактерии являются основной причиной смертности и заболеваний человека и вызывают такие инфекции, как столбняк (вызванный Clostridium tetani ), брюшной тиф , дифтерия , сифилис , холера , болезни пищевого происхождения , проказа (вызванная Mycobacterium leprae ) и туберкулез (вызванный Mycobacterium Tuberculosis ). ). [213] Патогенная причина известного заболевания может быть обнаружена только много лет спустя, как это было в случае с Helicobacter pylori и язвенной болезнью . [214] Бактериальные заболевания также играют важную роль в сельском хозяйстве : бактерии вызывают пятнистость листьев , ожоги и увядание растений, а также болезнь Ионе , мастит , сальмонеллез и сибирскую язву у сельскохозяйственных животных. [215]

Каждый вид возбудителя имеет характерный спектр взаимодействия с человеком- хозяином . Некоторые микроорганизмы, такие как стафилококк или стрептококк , могут вызывать кожные инфекции, пневмонию , менингит и сепсис , системную воспалительную реакцию, вызывающую шок , массивную вазодилатацию и смерть. [216] Тем не менее, эти организмы также являются частью нормальной флоры человека и обычно существуют на коже или в носу, не вызывая никаких заболеваний. Другие организмы неизменно вызывают заболевания у людей, например, Rickettsia , которые являются облигатными внутриклеточными паразитами, способными расти и размножаться только внутри клеток других организмов. Один вид риккетсий вызывает сыпной тиф , а другой — пятнистую лихорадку Скалистых гор . Хламидии , еще один тип облигатных внутриклеточных паразитов, содержат виды, которые могут вызывать пневмонию или инфекцию мочевыводящих путей , а также могут быть вовлечены в ишемическую болезнь сердца . [217] Некоторые виды, такие как Pseudomonas aeruginosa , Burkholderia cenocepacia и Mycobacterium avium , являются условно-патогенными микроорганизмами и вызывают заболевания преимущественно у людей с ослабленным иммунитетом или муковисцидозом . [218] [219] Некоторые бактерии производят токсины , вызывающие заболевания. [220] Это эндотоксины , которые образуются из разрушенных бактериальных клеток, и экзотоксины , которые производятся бактериями и выбрасываются в окружающую среду. [221] бактерия Clostridium botulinum Например, вырабатывает мощный экзотоксин, вызывающий паралич дыхания, а сальмонеллы производят эндотоксин, вызывающий гастроэнтерит. [221] Некоторые экзотоксины могут быть преобразованы в анатоксины , которые используются в качестве вакцин для предотвращения заболевания. [222]
Бактериальные инфекции можно лечить антибиотиками , которые классифицируются как бактерицидные, если они убивают бактерии, или бактериостатические, если они просто предотвращают рост бактерий. Существует много типов антибиотиков, и каждый класс ингибирует процесс, который у возбудителя отличается от процесса, обнаруженного у хозяина. Примером того, как антибиотики вызывают избирательную токсичность, являются хлорамфеникол и пуромицин , которые ингибируют бактериальную рибосому , но не структурно отличающуюся эукариотическую рибосому. [223] Антибиотики используются как для лечения заболеваний человека, так и в интенсивном сельском хозяйстве для стимулирования роста животных, где они могут способствовать быстрому развитию устойчивости к антибиотикам в популяциях бактерий. [224] Инфекции можно предотвратить с помощью антисептических мер, таких как стерилизация кожи перед прокалыванием ее иглой шприца, а также путем надлежащего ухода за постоянными катетерами. Хирургические и стоматологические инструменты также стерилизуются для предотвращения заражения бактериями. Дезинфицирующие средства, такие как отбеливатель, используются для уничтожения бактерий или других болезнетворных микроорганизмов на поверхностях, чтобы предотвратить загрязнение и еще больше снизить риск заражения. [225]
Значение в технологиях и промышленности
Бактерии, часто молочнокислые бактерии , такие как виды Lactobacillus и виды Lactococcus , в сочетании с дрожжами и плесенью , использовались в течение тысяч лет при приготовлении ферментированных продуктов, таких как сыр , соленые огурцы , соевый соус , квашеная капуста , уксус , вино. и йогурт . [226] [227]
Способность бактерий разлагать различные органические соединения замечательна и используется при переработке отходов и биоремедиации . способные переваривать углеводороды нефти Бактерии , , часто используются для очистки разливов нефти . [228] Удобрения были добавлены на некоторые пляжи в проливе Принца Уильяма в попытке способствовать росту этих естественных бактерий после Exxon Valdez разлива нефти в 1989 году . Эти усилия были эффективны на пляжах, которые не были слишком густо покрыты нефтью. Бактерии также используются для биоремедиации промышленных токсичных отходов . [229] В химической промышленности бактерии играют наиболее важную роль в производстве энантиомерно чистых химикатов для использования в фармацевтических и агрохимических препаратах . [230]
Бактерии также можно использовать вместо пестицидов при биологической борьбе с вредителями . Обычно это касается Bacillus thuringiensis (также называемой BT), грамположительной почвенной бактерии. Подвиды этих бактерий используются в качестве для чешуекрылых, , специфичных инсектицидов под торговыми названиями, такими как Dipel и Thuricide. [231] Из-за своей специфичности эти пестициды считаются экологически чистыми и практически не оказывают никакого воздействия на людей, дикую природу , опылителей и большинство других полезных насекомых . [232] [233]
Благодаря своей способности быстро расти и относительной легкости манипулирования ими, бактерии являются «рабочими лошадками» в области молекулярной биологии , генетики и биохимии . Выполняя мутации в бактериальной ДНК и исследуя полученные фенотипы, ученые могут определить функцию генов, ферментов и метаболических путей бактерий, а затем применить эти знания к более сложным организмам. [234] Эта цель понимания биохимии клетки достигает своего наиболее сложного выражения в синтезе огромного количества данных о кинетике ферментов и экспрессии генов в математические модели целых организмов. Это достижимо для некоторых хорошо изученных бактерий: Escherichia coli . в настоящее время создаются и тестируются модели метаболизма [235] [236] Такое понимание бактериального метаболизма и генетики позволяет использовать биотехнологии для биоинженерии бактерий для производства терапевтических белков, таких как инсулин , факторы роста или антитела . [237] [238]
Из-за их важности для исследований в целом образцы бактериальных штаммов изолируются и сохраняются в центрах биологических ресурсов . Это обеспечивает доступность штамма для ученых во всем мире. [239]
История бактериологии

Бактерии впервые наблюдал голландский микроскопист Антони ван Левенгук в 1676 году с помощью однолинзового микроскопа собственной конструкции. Затем он опубликовал свои наблюдения в серии писем Лондонскому королевскому обществу . [240] Бактерии были самым замечательным микроскопическим открытием Левенгука. Их размер был на пределе того, что могли разрешить его простые линзы, и в один из самых поразительных перерывов в истории науки никто больше не видел их больше столетия. [241] Его наблюдения также включали простейших, которых он назвал анимакулами , и его открытия были снова рассмотрены в свете более поздних открытий клеточной теории . [242]
Кристиан Готфрид Эренберг ввел слово «бактерия» в 1828 году. [243] Фактически, его бактерия представляла собой род, который содержал неспорообразующие палочковидные бактерии. [244] в отличие от Bacillus , рода спорообразующих палочковидных бактерий, определенного Эренбергом в 1835 году. [245]
Луи Пастер продемонстрировал в 1859 году, что рост микроорганизмов вызывает процесс брожения и что этот рост не происходит из-за самопроизвольного образования ( дрожжи и плесени , обычно связанные с брожением, являются не бактериями, а скорее грибами ). Вместе со своим современником Робертом Кохом Пастер был одним из первых сторонников микробной теории болезней . [246] До них Игнац Земмельвейс и Джозеф Листер осознали важность дезинфекции рук в медицинской работе. Земмельвейс, который в 1840-х годах сформулировал свои правила мытья рук в больнице, еще до появления микробной теории, объяснял болезнь «разложением органических веществ животных». Его идеи были отвергнуты, а его книга на эту тему осуждена медицинским сообществом. Однако после Листера в 1870-х годах врачи начали дезинфицировать руки. [247]
Роберт Кох, пионер медицинской микробиологии, работал над холерой , сибирской язвой и туберкулезом . В своих исследованиях туберкулеза Кох наконец доказал микробную теорию, за что получил Нобелевскую премию в 1905 году. [248] В постулатах Коха он изложил критерии проверки того, является ли организм причиной заболевания , и эти постулаты используются до сих пор. [249]
Фердинанд Кон считается основателем бактериологии , изучающим бактерии с 1870 года. Кон был первым, кто классифицировал бактерии на основе их морфологии. [250] [251]
Хотя в девятнадцатом веке было известно, что бактерии являются причиной многих заболеваний, эффективных антибактериальных методов лечения не существовало. [252] В 1910 году Пауль Эрлих разработал первый антибиотик, заменив красители, избирательно окрашивающие бледную трепонему — спирохету , вызывающую сифилис , на соединения, избирательно убивающие возбудитель. [253] Эрлих, который был удостоен Нобелевской премии 1908 года за свою работу в области иммунологии , был пионером в использовании красителей для обнаружения и идентификации бактерий; его работа легла в основу окрашивания по Граму и окрашивания по Цилю-Нильсену . [254]
Важный шаг вперед в изучении бактерий был сделан в 1977 году, когда Карл Везе признал, что археи имеют отдельную от бактерий линию эволюционного происхождения. [255] Эта новая филогенетическая таксономия зависела от секвенирования и 16S рибосомальной РНК разделяла прокариотов на два эволюционных домена как часть трехдоменной системы . [4]
См. также
Ссылки
- ^ «31. Древняя жизнь: микроокаменелости вершинного кремня» . www.lpi.usra.edu . Проверено 12 марта 2022 г.
- ^ Отправитель Р., Фукс С., Майло Р. (19 августа 2016 г.). «Пересмотренные оценки количества клеток человека и бактерий в организме» . ПЛОС Биология . 14 (8): e1002533. дои : 10.1371/journal.pbio.1002533 . ISSN 1545-7885 . ПМЦ 4991899 . ПМИД 27541692 .
- ^ Перейти обратно: а б Маккатчеон Дж.П. (октябрь 2021 г.). «Геномика и клеточная биология внутриклеточных инфекций, полезных для хозяина» . Ежегодный обзор клеточной биологии и биологии развития . 37 (1): 115–142. doi : 10.1146/annurev-cellbio-120219-024122 . ПМИД 34242059 . S2CID 235786110 .
- ^ Перейти обратно: а б с Холл 2008 , с. 145.
- ^ βακτήριον . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей» .
- ^ бактерия Лидделла и Скотта .
- ^ Харпер Д. «Бактерии» . Интернет-словарь этимологии .
- ^ Перейти обратно: а б Краснер 2014 , с. 74.
- ^ Вёзе Ч.Р., Кандлер О., Уилис М.Л. (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–79. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Холл 2008 , с. 84.
- ^ Годой-Виторино Ф (июль 2019 г.). «Микробная экология человека и развивающаяся новая медицина» . Анналы трансляционной медицины . 7 (14): 342. doi : 10.21037/атм.2019.06.56 . ПМК 6694241 . ПМИД 31475212 .
- ^ Шопф JW (июль 1994 г.). «Разные темпы, разные судьбы: темп и способ эволюции менялись от докембрия к фанерозою» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (15): 6735–42. Бибкод : 1994PNAS...91.6735S . дои : 10.1073/pnas.91.15.6735 . ПМЦ 44277 . ПМИД 8041691 .
- ^ Делонг Э.Ф., Пейс Н.Р. (август 2001 г.). «Экологическое разнообразие бактерий и архей». Систематическая биология . 50 (4): 470–78. CiteSeerX 10.1.1.321.8828 . дои : 10.1080/106351501750435040 . ПМИД 12116647 .
- ^ Браун-младший, Дулиттл В.Ф. (декабрь 1997 г.). «Археи и переход от прокариот к эукариотам» . Обзоры микробиологии и молекулярной биологии . 61 (4): 456–502. doi : 10.1128/mmbr.61.4.456-502.1997 . ПМК 232621 . ПМИД 9409149 .
- ^ Даум Б, Золото V (июнь 2018 г.). «Дергайся или плыви: к пониманию движения прокариот на основе схемы пилуса IV типа». Биологическая химия . 399 (7): 799–808. дои : 10.1515/hsz-2018-0157 . hdl : 10871/33366 . ПМИД 29894297 . S2CID 48352675 .
- ^ Ди Джулио М. (декабрь 2003 г.). «Универсальным предком и предком бактерий были гипертермофилы». Журнал молекулярной эволюции . 57 (6): 721–30. Бибкод : 2003JMolE..57..721D . дои : 10.1007/s00239-003-2522-6 . ПМИД 14745541 . S2CID 7041325 .
- ^ Баттистуцци Ф.У., Фейджао А., Хеджес С.Б. (ноябрь 2004 г.). «Геномная временная шкала эволюции прокариот: понимание происхождения метаногенеза, фототрофии и колонизации суши» . Эволюционная биология BMC . 4:44 . дои : 10.1186/1471-2148-4-44 . ПМЦ 533871 . ПМИД 15535883 .
- ^ Хоманн М., Сансьофре П., Ван Зуилен М., Хойбек С., Гонг Дж., Киллингсворт Б. и др. (23 июля 2018 г.). «Микробная жизнь и биогеохимический круговорот на суше 3220 миллионов лет назад» (PDF) . Природа Геонауки . 11 (9): 665–671. Бибкод : 2018NatGe..11..665H . дои : 10.1038/s41561-018-0190-9 . S2CID 134935568 .
- ^ Габальдон Т. (октябрь 2021 г.). «Происхождение и ранняя эволюция эукариотической клетки» . Ежегодный обзор микробиологии . 75 (1): 631–647. doi : 10.1146/annurev-micro-090817-062213 . ПМИД 34343017 . S2CID 236916203 . Архивировано из оригинала 19 августа 2022 года . Проверено 19 августа 2022 г.
- ^ Каллиер V (8 июня 2022 г.). «Митохондрии и происхождение эукариот» . Знающий журнал . doi : 10.1146/knowable-060822-2 . Проверено 19 августа 2022 г.
- ^ Пул А.М., Пенни Д. (январь 2007 г.). «Оценка гипотез происхождения эукариот». Биоэссе . 29 (1): 74–84. doi : 10.1002/bies.20516 . ПМИД 17187354 .
- ^ Дьялл С.Д., Браун М.Т., Джонсон П.Дж. (апрель 2004 г.). «Древние вторжения: от эндосимбионтов к органеллам». Наука . 304 (5668): 253–257. Бибкод : 2004Sci...304..253D . дои : 10.1126/science.1094884 . ПМИД 15073369 . S2CID 19424594 .
- ^ Стивенс Т.Г., Габр А., Калатрава В., Гроссман А.Р., Бхаттачарья Д. (сентябрь 2021 г.). «Почему первичный эндосимбиоз так редок?» . Новый фитолог . 231 (5): 1693–1699. дои : 10.1111/nph.17478 . ПМЦ 8711089 . ПМИД 34018613 .
- ^ Перейти обратно: а б Бейкер-Остин С., Допсон М. (апрель 2007 г.). «Жизнь в кислоте: гомеостаз pH у ацидофилов». Тенденции в микробиологии . 15 (4): 165–171. дои : 10.1016/j.tim.2007.02.005 . ПМИД 17331729 .
- ^ Чон С.В., Чхве Ю.Дж. (октябрь 2020 г.). «Экстремофильные микроорганизмы для очистки от токсичных загрязнителей окружающей среды» . Молекулы . 25 (21): 4916. doi : 10,3390/molecules25214916 . ПМК 7660605 . ПМИД 33114255 .
- ^ Флемминг ХК, Вюрц С (апрель 2019 г.). «Бактерии и археи на Земле и их численность в биопленках». Обзоры природы. Микробиология . 17 (4): 247–260. дои : 10.1038/s41579-019-0158-9 . ПМИД 30760902 . S2CID 61155774 .
- ^ Бар-Он Ю.М., Филлипс Р., Майло Р. (июнь 2018 г.). «Распределение биомассы на Земле» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (25): 6506–6511. Бибкод : 2018PNAS..115.6506B . дои : 10.1073/pnas.1711842115 . ПМК 6016768 . ПМИД 29784790 .
- ^ Уилис 2008 , с. 362.
- ^ Кушкевич И., Прохазка Ю., Гайдач М., Риттманн С.К., Витезова М. (июнь 2021 г.). «Молекулярная физиология анаэробных фототрофных пурпурных и зеленых серных бактерий» . Международный журнал молекулярных наук . 22 (12): 6398. doi : 10.3390/ijms22126398 . ПМЦ 8232776 . ПМИД 34203823 .
- ^ Уилис 2008 , с. 6.
- ^ Поммервилль 2014 , с. 3–6.
- ^ Перейти обратно: а б с Краснер 2014 , с. 38.
- ^ Перейти обратно: а б с д и ж г Поммервиль 2014 , с. 134.
- ^ Шульц Х.Н., Йоргенсен Б.Б. (2001). «Большие бактерии». Ежегодный обзор микробиологии . 55 : 105–137. дои : 10.1146/аннурев.микро.55.1.105 . ПМИД 11544351 . S2CID 18168018 .
- ^ Уильямс С (2011). «Кого ты называешь простым?». Новый учёный . 211 (2821): 38–41. дои : 10.1016/S0262-4079(11)61709-0 .
- ^ Волланд Дж.М., Гонсалес-Риццо С., Грос О., Тымл Т., Иванова Н., Шульц Ф., Гудо Д., Элизабет Н.Х., Нат Н., Удвари Д., Мальмстрем Р.Р. (18 февраля 2022 г.). «Бактерия длиной в сантиметр, ДНК которой разделена на мембраносвязанные органеллы» . bioRxiv (препринт). дои : 10.1101/2022.02.16.480423 . S2CID 246975579 .
- ^ Сандерсон К. (июнь 2022 г.). «Самая большая бактерия, когда-либо найденная, удивительно сложна». Природа . дои : 10.1038/d41586-022-01757-1 . ПМИД 35750919 . S2CID 250022076 .
- ^ Робертсон Дж., Гомерсолл М., Гилл П. (ноябрь 1975 г.). «Mycoplasma hominis: рост, размножение и выделение мелких жизнеспособных клеток» . Журнал бактериологии . 124 (2): 1007–1018. дои : 10.1128/JB.124.2.1007-1018.1975 . ПМК 235991 . ПМИД 1102522 .
- ^ Велимиров Б (2001). «Нанобактерии, ультрамикробактерии и голодающие формы: поиск мельчайших метаболизирующих бактерий» . Микробы и окружающая среда . 16 (2): 67–77. дои : 10.1264/jsme2.2001.67 .
- ^ Дюзенбери Д.Б. (2009). Жизнь в микромасштабе . Кембридж, Массачусетс: Издательство Гарвардского университета . стр. 20–25. ISBN 978-0-674-03116-6 .
- ^ Ян, округ Колумбия, Блэр К.М., Салама Н.Р. (март 2016 г.). «Оставаться в форме: влияние формы клеток на выживание бактерий в различных средах» . Обзоры микробиологии и молекулярной биологии . 80 (1): 187–203. дои : 10.1128/MMBR.00031-15 . ПМЦ 4771367 . ПМИД 26864431 .
- ^ Кабин М.Т., Джейкобс-Вагнер С. (август 2005 г.). «Форма бактериальной клетки». Обзоры природы. Микробиология . 3 (8): 601–10. дои : 10.1038/nrmicro1205 . ПМИД 16012516 . S2CID 23938989 .
- ^ Молодой К.Д. (сентябрь 2006 г.). «Селективное значение формы бактерий» . Обзоры микробиологии и молекулярной биологии . 70 (3): 660–703. дои : 10.1128/MMBR.00001-06 . ПМЦ 1594593 . ПМИД 16959965 .
- ^ Кроуфорд 2007 , с. xi.
- ^ Классен Д., Розен Д.Е., Койперс О.П., Согаард-Андерсен Л., ван Везель Г.П. (февраль 2014 г.). «Бактериальные решения проблемы многоклеточности: рассказ о биопленках, нитях и плодовых телах» . Обзоры природы. Микробиология . 12 (2): 115–24. дои : 10.1038/nrmicro3178 . hdl : 11370/0db66a9c-72ef-4e11-a75d-9d1e5827573d . ПМИД 24384602 . S2CID 20154495 .
- ^ Шимкец ЖЖ (1999). «Межклеточная передача сигналов во время развития плодовых тел Myxococcus xanthus». Ежегодный обзор микробиологии . 53 : 525–49. дои : 10.1146/аннурев.микро.53.1.525 . ПМИД 10547700 .
- ^ Перейти обратно: а б Кайзер Д. (2004). «Сигнализация в миксобактериях». Ежегодный обзор микробиологии . 58 : 75–98. дои : 10.1146/annurev.micro.58.030603.123620 . ПМИД 15487930 .
- ^ Уилис 2008 , с. 75.
- ^ Мандал А., Дутта А., Дас Р., Мукерджи Дж. (июнь 2021 г.). «Роль приливных микробных сообществ в секвестрации углекислого газа и удалении загрязняющих веществ: обзор». Бюллетень о загрязнении морской среды . 170 : 112626. Бибкод : 2021MarPB.17012626M . doi : 10.1016/j.marpolbul.2021.112626 . ПМИД 34153859 .
- ^ Донлан Р.М. (сентябрь 2002 г.). «Биопленки: микробная жизнь на поверхностях» . Новые инфекционные заболевания . 8 (9): 881–90. дои : 10.3201/eid0809.020063 . ПМЦ 2732559 . ПМИД 12194761 .
- ^ Бранда С.С., Вик С., Фридман Л., Колтер Р. (январь 2005 г.). «Биопленки: новый взгляд на матрицу». Тенденции в микробиологии . 13 (1): 20–26. дои : 10.1016/j.tim.2004.11.006 . ПМИД 15639628 .
- ^ Перейти обратно: а б Дэйви М.Э., О'Тул Дж.А. (декабрь 2000 г.). «Микробные биопленки: от экологии к молекулярной генетике» . Обзоры микробиологии и молекулярной биологии . 64 (4): 847–67. дои : 10.1128/MMBR.64.4.847-867.2000 . ПМК 99016 . ПМИД 11104821 .
- ^ Донлан Р.М., Костертон Дж.В. (апрель 2002 г.). «Биопленки: механизмы выживания клинически значимых микроорганизмов» . Обзоры клинической микробиологии . 15 (2): 167–93. doi : 10.1128/CMR.15.2.167-193.2002 . ПМК 118068 . ПМИД 11932229 .
- ^ Слончевски Дж.Л., Фостер Дж.В. (2013). Микробиология: развивающаяся наука (Третье изд.). Нью-Йорк: WW Нортон. п. 82. ИСБН 978-0-393-12367-8 .
- ^ Фейхоо-Сиота Л., Рама Х.Л., Санчес-Перес А., Вилла Т.Г. (июль 2017 г.). «Соображения о бактериальных нуклеоидах». Прикладная микробиология и биотехнология . 101 (14): 5591–602. дои : 10.1007/s00253-017-8381-7 . ПМИД 28664324 . S2CID 10173266 .
- ^ Бобик Т.А. (май 2006 г.). «Многогранные органеллы, разделяющие бактериальные метаболические процессы». Прикладная микробиология и биотехнология . 70 (5): 517–25. дои : 10.1007/s00253-005-0295-0 . ПМИД 16525780 . S2CID 8202321 .
- ^ Йейтс Т.О., Керфельд К.А., Хайнхорст С., Кэннон Г.К., Шайвли Дж.М. (сентябрь 2008 г.). «Белковые органеллы бактерий: карбоксисомы и родственные микрокомпарты». Обзоры природы. Микробиология . 6 (9): 681–91. дои : 10.1038/nrmicro1913 . ПМИД 18679172 . S2CID 22666203 .
- ^ Керфельд К.А., Савая М.Р., Танака С., Нгуен К.В., Филлипс М., Биби М., Йейтс Т.О. (август 2005 г.). «Белковые структуры, образующие оболочку примитивных бактериальных органелл». Наука . 309 (5736): 936–38. Бибкод : 2005Sci...309..936K . CiteSeerX 10.1.1.1026.896 . дои : 10.1126/science.1113397 . ПМИД 16081736 . S2CID 24561197 .
- ^ Гитай З. (март 2005 г.). «Новая биология бактериальных клеток: движущиеся части и субклеточная архитектура» . Клетка . 120 (5): 577–86. дои : 10.1016/j.cell.2005.02.026 . ПМИД 15766522 . S2CID 8894304 .
- ^ Ши Ю.Л., Ротфилд Л. (сентябрь 2006 г.). «Бактериальный цитоскелет» . Обзоры микробиологии и молекулярной биологии . 70 (3): 729–54. дои : 10.1128/MMBR.00017-06 . ПМЦ 1594594 . ПМИД 16959967 .
- ^ Норрис В., ден Блаувен Т., Кабин-Фламан А., Дой Р.Х., Харши Р., Яньер Л., Хименес-Санчес А., Джин Д.Д., Левин П.А., Милейковская Е., Мински А., Сайер М., Скарстад К. (март 2007 г.). «Функциональная систематика бактериальных гиперструктур» . Обзоры микробиологии и молекулярной биологии . 71 (1): 230–53. дои : 10.1128/MMBR.00035-06 . ПМЦ 1847379 . ПМИД 17347523 .
- ^ Поммервилль, 2014 , стр. 120–121.
- ^ Брайант Д.А., Фригаард Н.У. (ноябрь 2006 г.). «Освещение фотосинтеза и фототрофии прокариот». Тенденции в микробиологии . 14 (11): 488–96. дои : 10.1016/j.tim.2006.09.001 . ПМИД 16997562 .
- ^ Псенчик Дж., Иконен Т.П., Лауринмяки П., Меркель М.С., Батчер С.Дж., Серимаа Р.Э., Тума Р. (август 2004 г.). «Ламеллярная организация пигментов в хлоросомах — светособирающих комплексах зеленых фотосинтезирующих бактерий» . Биофизический журнал . 87 (2): 1165–72. Бибкод : 2004BpJ....87.1165P . дои : 10.1529/biophysj.104.040956 . ПМК 1304455 . ПМИД 15298919 .
- ^ Танбихлер М., Ван С.К., Шапиро Л. (октябрь 2005 г.). «Бактериальный нуклеоид: высокоорганизованная и динамичная структура» . Журнал клеточной биохимии . 96 (3): 506–21. дои : 10.1002/jcb.20519 . ПМИД 15988757 . S2CID 25355087 .
- ^ Поелсгаард Дж., Даутвейт С. (ноябрь 2005 г.). «Бактериальная рибосома как мишень для антибиотиков». Обзоры природы. Микробиология . 3 (11): 870–81. дои : 10.1038/nrmicro1265 . ПМИД 16261170 . S2CID 7521924 .
- ^ Йео М., Чейтер К. (март 2005 г.). «Взаимодействие метаболизма и дифференцировки гликогена дает представление о биологии развития Streptomyces coelicolor» . Микробиология . 151 (Часть 3): 855–61. дои : 10.1099/mic.0.27428-0 . ПМИД 15758231 . Архивировано из оригинала 29 сентября 2007 года.
- ^ Шиба Т., Цуцуми К., Исигэ К., Ногучи Т. (март 2000 г.). «Неорганический полифосфат и полифосфаткиназа: их новые биологические функции и применение» . Биохимия. Биохимия . 65 (3): 315–23. ПМИД 10739474 . Архивировано из оригинала 25 сентября 2006 года.
- ^ Брюн, округ Колумбия (июнь 1995 г.). «Выделение и характеристика белков глобул серы из Chromatium vinosum и Thiocapsaroseopersicina». Архив микробиологии . 163 (6): 391–99. Бибкод : 1995ArMic.163..391B . дои : 10.1007/BF00272127 . ПМИД 7575095 . S2CID 22279133 .
- ^ Кадури Д., Юркевич Е., Окон Ю., Кастро-Совински С. (2005). «Экологическое и сельскохозяйственное значение бактериальных полигидроксиалканоатов». Критические обзоры по микробиологии . 31 (2): 55–67. дои : 10.1080/10408410590899228 . ПМИД 15986831 . S2CID 4098268 .
- ^ Уолсби А.Е. (март 1994 г.). «Газовые пузырьки» . Микробиологические обзоры . 58 (1): 94–144. дои : 10.1128/ММБР.58.1.94-144.1994 . ПМК 372955 . ПМИД 8177173 .
- ^ ван Хейеноорт Дж. (март 2001 г.). «Образование гликановых цепей при синтезе бактериального пептидогликана» . Гликобиология . 11 (3): 25П–36П. дои : 10.1093/гликоб/11.3.25R . ПМИД 11320055 . S2CID 46066256 .
- ^ Перейти обратно: а б Кох А.Л. (октябрь 2003 г.). «Бактериальная стена как мишень для нападения: исследования прошлого, настоящего и будущего» . Обзоры клинической микробиологии . 16 (4): 673–87. doi : 10.1128/CMR.16.4.673-687.2003 . ПМК 207114 . ПМИД 14557293 .
- ^ Перейти обратно: а б Грамм ХК (1884 г.). «Об изолированном окрашивании шизомицетов в срезанных и сухих препаратах». Прогресс Мед . 2 : 185-89.
- ^ Гугенгольц П. (2002). «Изучение прокариотического разнообразия в эпоху генома» . Геномная биология . 3 (2): ОБЗОРЫ0003. doi : 10.1186/gb-2002-3-2-reviews0003 . ПМК 139013 . ПМИД 11864374 .
- ^ Уолш FM, Эмис С.Г. (октябрь 2004 г.). «Микробиология и механизмы лекарственной устойчивости полностью устойчивых патогенов» (PDF) . Современное мнение в микробиологии . 7 (5): 439–44. дои : 10.1016/j.mib.2004.08.007 . ПМИД 15451497 .
- ^ Олдервик Л.Дж., Харрисон Дж., Ллойд Г.С., Берч Х.Л. (март 2015 г.). «Клеточная стенка микобактерий – пептидогликан и арабиногалактан» . Перспективы Колд-Спринг-Харбора в медицине . 5 (8): а021113. doi : 10.1101/cshperspect.a021113 . ПМЦ 4526729 . ПМИД 25818664 .
- ^ Фэган Р.П., Фэйрвезер Н.Ф. (март 2014 г.). «Биогенез и функции бактериальных S-слоев» (PDF) . Обзоры природы. Микробиология . 12 (3): 211–22. дои : 10.1038/nrmicro3213 . ПМИД 24509785 . S2CID 24112697 .
- ^ Томпсон С.А. (декабрь 2002 г.). «Поверхностные слои Campylobacter (S-слои) и уклонение от иммунитета» . Анналы пародонтологии . 7 (1): 43–53. дои : 10.1902/анналы.2002.7.1.43 . ПМЦ 2763180 . ПМИД 16013216 .
- ^ Беверидж Т.Дж., Пауэлс П.Х., Сара М., Котиранта А., Лунатмаа К., Кари К., Керосуо Е., Хаапасало М., Эгельсир Э.М., Шохер И., Слейтр У.Б., Морелли Л., Каллегари М.Л., Номеллини Дж.Ф., Бингл В.Х., Смит Дж., Лейбовиц Е. , Лемэр М., Мирас И., Саламиту С., Беген П., Охайон Х., Гунон П., Матушек М., Коваль С.Ф. (июнь 1997 г.). «Функции S-слоев». Обзоры микробиологии FEMS . 20 (1–2): 99–149. дои : 10.1111/j.1574-6976.1997.tb00305.x . ПМИД 9276929 .
- ^ Кодзима С., Блер Д.Ф. (2004). Бактериальный жгутиковый мотор: структура и функция сложной молекулярной машины . Международный обзор цитологии. Том. 233. С. 93–134. дои : 10.1016/S0074-7696(04)33003-2 . ISBN 978-0-12-364637-8 . ПМИД 15037363 .
- ^ Уилис 2008 , с. 76.
- ^ Ченг Р.А., Видманн М. (2020). «Последние достижения в нашем понимании разнообразия и роли фимбрий шаперона-Ашера в содействии тропизму хозяина сальмонеллы и тканям» . Границы клеточной и инфекционной микробиологии . 10 : 628043. дои : 10.3389/fcimb.2020.628043 . ПМЦ 7886704 . ПМИД 33614531 .
- ^ Сильверман ПМ (февраль 1997 г.). «К структурной биологии бактериальной конъюгации» . Молекулярная микробиология . 23 (3): 423–29. дои : 10.1046/j.1365-2958.1997.2411604.x . ПМИД 9044277 . S2CID 24126399 .
- ^ Коста Т.Р., Фелисберто-Родригес К., Меир А., Превост М.С., Реджей А., Троктер М., Ваксман Г. (июнь 2015 г.). «Системы секреции грамотрицательных бактерий: структурные и механические сведения». Обзоры природы. Микробиология . 13 (6): 343–59. дои : 10.1038/nrmicro3456 . ПМИД 25978706 . S2CID 8664247 .
- ^ Луонг П., Дубе Д.Х. (июль 2021 г.). «Демонтаж бактериального гликокаликса: химические инструменты для исследования, возмущения и изображения бактериальных гликанов» . Биоорганическая и медицинская химия . 42 : 116268. doi : 10.1016/j.bmc.2021.116268 . ISSN 0968-0896 . ПМЦ 8276522 . ПМИД 34130219 .
- ^ Стоукс Р.В., Норрис-Джонс Р., Брукс Д.Е., Беверидж Т.Дж., Докси Д., Торсон Л.М. (октябрь 2004 г.). «Богатый гликанами внешний слой клеточной стенки Mycobacterium Tuberculosis действует как антифагоцитарная капсула, ограничивающая ассоциацию бактерии с макрофагами» . Инфекция и иммунитет . 72 (10): 5676–86. дои : 10.1128/IAI.72.10.5676-5686.2004 . ПМК 517526 . ПМИД 15385466 .
- ^ Перейти обратно: а б Калшойер Р., Паласиос А., Ансо И., Сифуэнте Дж., Ангита Дж., Джейкобс В.Р., Герен М.Е., Прадос-Росалес Р. (июль 2019 г.). «Капсула микобактерии туберкулеза: клеточная структура, имеющая ключевое значение в патогенезе» . Биохимический журнал . 476 (14): 1995–2016. дои : 10.1042/BCJ20190324 . ПМК 6698057 . ПМИД 31320388 .
- ^ Джерниган Дж.А., Стивенс Д.С., Эшфорд Д.А., Оменака К., Топиэль М.С., Гэлбрейт М., Таппер М., Фиск Т.Л., Заки С., Попович Т., Мейер Р.Ф., Куинн К.П., Харпер С.А., Фридкин С.К., Сейвар Дж.Дж., Шепард К.В., МакКоннелл М. , Гуарнер Дж., Ши В.Дж., Малецки Дж.М., Гербердинг Дж.Л., Хьюз Дж.М., Перкинс Б.А. (2001). «Ингаляционная форма сибирской язвы, связанная с биотерроризмом: первые 10 случаев зарегистрированы в США» . Новые инфекционные заболевания . 7 (6): 933–44. дои : 10.3201/eid0706.010604 . ПМК 2631903 . ПМИД 11747719 .
- ^ Николсон В.Л., Мунаката Н., Хорнек Г., Мелош Х.Дж., Сетлоу П. (сентябрь 2000 г.). «Устойчивость эндоспор Bacillus к экстремальным земным и внеземным средам» . Обзоры микробиологии и молекулярной биологии . 64 (3): 548–72. дои : 10.1128/MMBR.64.3.548-572.2000 . ПМК 99004 . ПМИД 10974126 .
- ^ Перейти обратно: а б Маккенни П.Т., Дрикс А., Айхенбергер П. (январь 2013 г.). «Эндоспора Bacillus subtilis: сборка и функции многослойной оболочки» . Обзоры природы. Микробиология . 11 (1): 33–44. дои : 10.1038/nrmicro2921 . ПМК 9910062 . ПМИД 23202530 . S2CID 205498395 .
- ^ Николсон В.Л., Фахардо-Кавасос П., Ребейл Р., Слиман Т.А., Ризенман П.Дж., Лоу Дж.Ф., Сюэ Ю (август 2002 г.). «Бактериальные эндоспоры и их значение в устойчивости к стрессу». Антони ван Левенгук . 81 (1–4): 27–32. дои : 10.1023/A:1020561122764 . ПМИД 12448702 . S2CID 30639022 .
- ^ Вриланд Р.Х., Розенцвейг В.Д., Пауэрс Д.В. (октябрь 2000 г.). «Выделение галотолерантной бактерии возрастом 250 миллионов лет из первичного кристалла соли». Природа . 407 (6806): 897–900. Бибкод : 2000Natur.407..897V . дои : 10.1038/35038060 . ПМИД 11057666 . S2CID 9879073 .
- ^ Кано Р.Дж., Боруки М.К. (май 1995 г.). «Возрождение и идентификация бактериальных спор в доминиканском янтаре возрастом от 25 до 40 миллионов лет». Наука . 268 (5213): 1060–64. Бибкод : 1995Sci...268.1060C . дои : 10.1126/science.7538699 . ПМИД 7538699 .
- ^ «Ссора над древними бактериями» . Новости Би-би-си . 7 июня 2001 года . Проверено 26 апреля 2020 г. .
- ^ Николсон В.Л., Шуергер А.С., Сетлоу П. (апрель 2005 г.). «Солнечная УФ-среда и устойчивость бактериальных спор к УФ-излучению: соображения по транспортировке с Земли на Марс посредством естественных процессов и полета человека в космос». Мутационные исследования . 571 (1–2): 249–64. дои : 10.1016/j.mrfmmm.2004.10.012 . ПМИД 15748651 .
- ^ «Колонизировать галактику сложно. Почему бы вместо этого не послать бактерии?» . Экономист . 12 апреля 2018 г. ISSN 0013-0613 . Проверено 26 апреля 2020 г. .
- ^ Revitt-Mills SA, Видор С.Дж., Уоттс Т.Д., Лирас Д., Руд Дж.И., Адамс В. (май 2019 г.). «Плазмиды вирулентности патогенных клостридий» . Микробиологический спектр . 7 (3). doi : 10.1128/microbiolspec.GPP3-0034-2018 . ПМИД 31111816 . S2CID 160013108 .
- ^ Рейгадас Э., ван Прен Дж., Фальконе М., Фицпатрик Ф., Верешильд М.Дж., Куйпер Э.Дж., Буза Э. (июль 2021 г.). «Как: профилактические мероприятия для предотвращения инфекции Clostridioides difficile» . Клиническая микробиология и инфекции . 27 (12): 1777–1783. дои : 10.1016/j.cmi.2021.06.037 . hdl : 1887/3249077 . ПМИД 34245901 .
- ^ Нилсон К.Х. (январь 1999 г.). «Микробиология после викингов: новые подходы, новые данные, новые идеи». Происхождение жизни и эволюция биосферы . 29 (1): 73–93. Бибкод : 1999ОЛЕВ...29...73Н . дои : 10.1023/А:1006515817767 . ПМИД 11536899 . S2CID 12289639 .
- ^ Сюй Дж (июнь 2006 г.). «Микробная экология в эпоху геномики и метагеномики: концепции, инструменты и последние достижения» . Молекулярная экология . 15 (7): 1713–31. дои : 10.1111/j.1365-294X.2006.02882.x . ПМИД 16689892 . S2CID 16374800 .
- ^ Зиллиг В. (декабрь 1991 г.). «Сравнительная биохимия архей и бактерий». Текущее мнение в области генетики и развития . 1 (4): 544–51. дои : 10.1016/S0959-437X(05)80206-0 . ПМИД 1822288 .
- ^ Перейти обратно: а б с Слончевски Дж.Л., Фостер Дж.В. Микробиология: развивающаяся наука (3-е изд.). WW Нортон и компания. стр. 491–44.
- ^ Хеллингверф К.Дж., Крилаард В., Хофф В.Д., Маттейс Х.К., Мур Л.Р., ван Роттердам Б.Дж. (1994). «Фотобиология бактерий» . Антони ван Левенгук (Представлена рукопись). 65 (4): 331–47. дои : 10.1007/BF00872217 . ПМИД 7832590 . S2CID 23438926 .
- ^ Далтон Х (июнь 2005 г.). «Лекция Левенгука 2000 года о естественной и неестественной истории метанокисляющих бактерий» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 360 (1458): 1207–22. дои : 10.1098/rstb.2005.1657 . ПМК 1569495 . ПМИД 16147517 .
- ^ Имран А., Хаким С., Тарик М., Наваз М.С., Лараиб И., Гульзар У., Ханиф М.К., Сиддик М.Дж., Хаят М., Фраз А., Ахмад М. (2021). «Диазотрофы для снижения кризисов, вызванных азотным загрязнением: взгляд глубоко в корни» . Границы микробиологии . 12 : 637815. doi : 10.3389/fmicb.2021.637815 . ПМК 8180554 . ПМИД 34108945 .
- ^ Зер Дж.П., Дженкинс Б.Д., Шорт С.М., Стюард Г.Ф. (июль 2003 г.). «Разнообразие генов нитрогеназы и структура микробного сообщества: межсистемное сравнение» . Экологическая микробиология . 5 (7): 539–54. Бибкод : 2003EnvMi...5..539Z . дои : 10.1046/j.1462-2920.2003.00451.x . ПМИД 12823187 .
- ^ Косуги Й., Мацуура Н., Лян К., Ямамото-Икемото Р. (октябрь 2020 г.). «Очистка сточных вод с использованием процесса сульфатредукции, денитрификации Анаммокс и частичной нитрификации (SRDAPN)». Хемосфера . 256 : 127092. Бибкод : 2020Chmsp.25627092K . doi : 10.1016/j.chemSphere.2020.127092 . ПМИД 32559887 . S2CID 219476361 .
- ^ Морель Ф.М., Крапил А.М., Амиот М. (1998). «Химический круговорот и биоаккумуляция ртути». Ежегодный обзор экологии и систематики . 29 : 543–66. doi : 10.1146/annurev.ecolsys.29.1.543 . S2CID 86336987 .
- ^ Слесак И, Кула М, Слесак Х, Мишальски З, Стшалка К (август 2019 г.). «Как определить обязательный анаэробиоз? Эволюционный взгляд на систему антиоксидантного ответа и ранние этапы эволюции жизни на Земле» . Свободно-радикальная биология и медицина . 140 : 61–73. doi : 10.1016/j.freeradbiomed.2019.03.004 . ПМИД 30862543 .
- ^ Стюарт Э.Дж., Мэдден Р., Пол Дж., Таддей Ф. (февраль 2005 г.). «Старение и смерть организма, размножающегося морфологически симметричным делением» . ПЛОС Биология . 3 (2): е45. doi : 10.1371/journal.pbio.0030045 . ПМК 546039 . ПМИД 15685293 .
- ^ Кох А.Л. (2002). «Контроль бактериального клеточного цикла путем роста цитоплазмы». Критические обзоры по микробиологии . 28 (1): 61–77. дои : 10.1080/1040-840291046696 . ПМИД 12003041 . S2CID 11624182 .
- ^ Поммервилль 2014 , с. 138.
- ^ Поммервилль 2014 , с. 557.
- ^ Перейти обратно: а б Уилис 2008 , с. 42.
- ^ Перейти обратно: а б с Томсон Р.Б., Бертрам Х. (декабрь 2001 г.). «Лабораторная диагностика инфекций центральной нервной системы». Клиники инфекционных заболеваний Северной Америки . 15 (4): 1047–71. дои : 10.1016/S0891-5520(05)70186-0 . ПМИД 11780267 .
- ^ Паерл Х.В., Фултон Р.С., Мойсандер П.Х., Дайбл Дж. (апрель 2001 г.). «Вредное цветение пресноводных водорослей с акцентом на цианобактерии» . Научный мировой журнал . 1 : 76–113. дои : 10.1100/tsw.2001.16 . ПМК 6083932 . ПМИД 12805693 .
- ^ Чаллис Г.Л., Хопвуд Д.А. (ноябрь 2003 г.). «Синергия и непредвиденные обстоятельства как движущие силы эволюции производства множественных вторичных метаболитов видами Streptomyces» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (Приложение 2): 14555–61. Бибкод : 2003PNAS..10014555C . дои : 10.1073/pnas.1934677100 . ПМК 304118 . ПМИД 12970466 .
- ^ Койман С.А., Оже П., Поджиале Дж.К., Кои Б.В. (август 2003 г.). «Количественные этапы симбиогенеза и эволюция гомеостаза». Биологические обзоры Кембриджского философского общества . 78 (3): 435–63. дои : 10.1017/S1464793102006127 . ПМИД 14558592 . S2CID 41072709 .
- ^ Бертран РЛ (2019). «Лаг-фаза — это динамичный, организованный, адаптивный и развивающийся период, который подготавливает бактерии к делению клеток» . Журнал бактериологии . 201 (7): e00697-18. дои : 10.1128/JB.00697-18 . ПМК 6416914 . ПМИД 30642990 .
- ^ Пратс К., Лопес Д., Жиро А., Феррер Дж., Валлс Дж. (август 2006 г.). «Индивидуальное моделирование бактериальных культур для изучения микроскопических причин лаг-фазы». Журнал теоретической биологии . 241 (4): 939–53. Бибкод : 2006JThBi.241..939P . дои : 10.1016/j.jtbi.2006.01.029 . ПМИД 16524598 .
- ^ Хекер М., Фёлькер Ю (2001). Общая реакция на стресс Bacillus subtilis и других бактерий . Достижения микробной физиологии. Том. 44. С. 35–91. дои : 10.1016/S0065-2911(01)44011-2 . ISBN 978-0-12-027744-5 . ПМИД 11407115 .
- ^ Слончевски Дж.Л., Фостер Дж.В. Микробиология: развивающаяся наука (3-е изд.). WW Нортон и компания. п. 143.
- ^ Леппянен М., Сундберг Л.Р., Лаанто Э., де Фрейтас Алмейда ГМ, Паппонен П., Маасилта И.Ю. (август 2017 г.). «Визуализация бактериальных колоний и взаимодействия фага и бактерии с субнанометровым разрешением с использованием гелий-ионной микроскопии» . Продвинутые биосистемы . 1 (8): e1700070. дои : 10.1002/adbi.201700070 . ПМИД 32646179 . S2CID 90960276 .
- ^ Накабати А., Ямасита А., Тох Х., Исикава Х., Данбар Х.Э., Моран Н.А., Хаттори М. (октябрь 2006 г.). «Геном бактериального эндосимбионта Carsonella длиной 160 тысяч оснований». Наука . 314 (5797): 267. doi : 10.1126/science.1134196 . ПМИД 17038615 . S2CID 44570539 .
- ^ Праделла С., Ханс А., Шпроер С., Райхенбах Х., Герт К., Бейер С. (декабрь 2002 г.). «Характеристика, размер генома и генетические манипуляции миксобактерии Sorangium cellulosum So ce56». Архив микробиологии . 178 (6): 484–92. Бибкод : 2002ArMic.178..484P . дои : 10.1007/s00203-002-0479-2 . ПМИД 12420170 . S2CID 21023021 .
- ^ Хиннебуш Дж., Тилли К. (декабрь 1993 г.). «Линейные плазмиды и хромосомы бактерий» . Молекулярная микробиология . 10 (5): 917–22. дои : 10.1111/j.1365-2958.1993.tb00963.x . ПМИД 7934868 . S2CID 23852021 .
- ^ Лин Ю.С., Кизер Х.М., Хопвуд Д.А., Чен К.В. (декабрь 1993 г.). «Хромосомная ДНК Streptomyces lividans 66 является линейной». Молекулярная микробиология . 10 (5): 923–33. дои : 10.1111/j.1365-2958.1993.tb00964.x . ПМИД 7934869 . S2CID 8536066 .
- ^ Вал М.Е., Солер-Бистю А., Бланд М.Дж., Мазель Д. (декабрь 2014 г.). «Управление многочастными геномами: модель холерного вибриона» . Современное мнение в микробиологии . 22 : 120–26. дои : 10.1016/j.mib.2014.10.003 . ПМИД 25460805 . S2CID 215743285 .
- ^ Кадо CI (октябрь 2014 г.). «Исторические события, породившие область плазмидной биологии» . Микробиологический спектр . 2 (5): 3. doi : 10.1128/microbiolspec.PLAS-0019-2013 . ПМИД 26104369 .
- ^ Белфорт М. , Рибан М.Э., Кутзи Т., Далгаард Дж.З. (июль 1995 г.). «Прокариотические интроны и интеины: множество форм и функций» . Журнал бактериологии . 177 (14): 3897–903. дои : 10.1128/jb.177.14.3897-3903.1995 . ПМК 177115 . ПМИД 7608058 .
- ^ Денамур Э., Матич I (май 2006 г.). «Эволюция частоты мутаций у бактерий» . Молекулярная микробиология . 60 (4): 820–27. дои : 10.1111/j.1365-2958.2006.05150.x . ПМИД 16677295 . S2CID 20713095 .
- ^ Райт Б.Е. (май 2004 г.). «Адаптивные мутации и эволюция, направленные на стресс» . Молекулярная микробиология . 52 (3): 643–50. дои : 10.1111/j.1365-2958.2004.04012.x . ПМИД 15101972 . S2CID 1071308 .
- ^ Чен И, Дубнау Д (март 2004 г.). «Поглощение ДНК во время бактериальной трансформации». Обзоры природы. Микробиология . 2 (3): 241–49. дои : 10.1038/nrmicro844 . ПМИД 15083159 . S2CID 205499369 .
- ^ Джонсборг О., Эльдхольм В., Ховарштайн Л.С. (декабрь 2007 г.). «Естественная генетическая трансформация: распространенность, механизмы и функции» . Исследования в области микробиологии . 158 (10): 767–78. дои : 10.1016/j.resmic.2007.09.004 . ПМИД 17997281 .
- ^ Бернштейн Х, Бернштейн С, Мишод Р.Э. (2012). «Репарация ДНК как основная адаптивная функция пола у бактерий и эукариот». Глава 1: стр. 1–49 в: Восстановление ДНК: новые исследования , Сакура Кимура и Сора Симидзу (ред.). Новая наука. Издательство, Хоппож, Нью-Йорк ISBN 978-1-62100-808-8 .
- ^ Брюссов Х., Канчая С., Хардт В.Д. (сентябрь 2004 г.). «Фаги и эволюция бактериальных патогенов: от геномных перестроек к лизогенной конверсии» . Обзоры микробиологии и молекулярной биологии . 68 (3): 560–602, оглавление. дои : 10.1128/MMBR.68.3.560-602.2004 . ПМК 515249 . ПМИД 15353570 .
- ^ Бикл Т.А., Крюгер Д.Х. (июнь 1993 г.). «Биология рестрикции ДНК» . Микробиологические обзоры . 57 (2): 434–50. дои : 10.1128/MMBR.57.2.434-450.1993 . ПМК 372918 . ПМИД 8336674 .
- ^ Баррангу Р., Фремо С., Дево Х., Ричардс М., Бояваль П., Муано С., Ромеро Д.А., Хорват П. (март 2007 г.). «CRISPR обеспечивает приобретенную устойчивость прокариот к вирусам». Наука . 315 (5819): 1709–12. Бибкод : 2007Sci...315.1709B . дои : 10.1126/science.1138140 . hdl : 20.500.11794/38902 . ПМИД 17379808 . S2CID 3888761 .
- ^ Браунс С.Дж., Джор М.М., Лундгрен М., Вестра Э.Р., Слейкхейс Р.Дж., Снейдерс А.П., Дикман М.Дж., Макарова К.С., Кунин Е.В., ван дер Оост Дж. (август 2008 г.). «Маленькие РНК CRISPR обеспечивают противовирусную защиту у прокариот» . Наука . 321 (5891): 960–64. Стартовый код : 2008Sci...321..960B . дои : 10.1126/science.1159689 . ПМЦ 5898235 . ПМИД 18703739 .
- ^ Кабесон Э, Риполь-Розада Х, Пенья А, де ла Крус Ф, Аречага I (январь 2015 г.). «На пути к интегрированной модели бактериальной конъюгации» . Обзоры микробиологии FEMS . 39 (1): 81–95. дои : 10.1111/1574-6976.12085 . ПМИД 25154632 . S2CID 34245150 .
- ^ Мишод Р.Э., Бернштейн Х., Недельку А.М. (май 2008 г.). «Адаптивное значение пола у микробных патогенов» (PDF) . Инфекция, генетика и эволюция . 8 (3): 267–85. дои : 10.1016/j.meegid.2008.01.002 . ПМИД 18295550 . Архивировано (PDF) из оригинала 30 декабря 2016 года.
- ^ Гастингс П.Дж., Розенберг С.М., Slack A (сентябрь 2004 г.). «Боковая передача устойчивости к антибиотикам, вызванная антибиотиками». Тенденции в микробиологии . 12 (9): 401–14. дои : 10.1016/j.tim.2004.07.003 . ПМИД 15337159 .
- ^ Дэвисон Дж. (сентябрь 1999 г.). «Генетический обмен между бактериями в окружающей среде». Плазмида . 42 (2): 73–91. дои : 10.1006/plas.1999.1421 . ПМИД 10489325 .
- ^ Перейти обратно: а б с Ким К.В. (декабрь 2017 г.). «Электронно-микроскопические наблюдения поверхностных придатков прокариот». Журнал микробиологии (Сеул, Корея) . 55 (12): 919–26. дои : 10.1007/s12275-017-7369-4 . ПМИД 29214488 . S2CID 89872403 .
- ^ Макнаб РМ (декабрь 1999 г.). «Бактериальный жгутик: реверсивный вращающийся пропеллер и экспортный аппарат типа III» . Журнал бактериологии . 181 (23): 7149–53. дои : 10.1128/JB.181.23.7149-7153.1999 . ПМЦ 103673 . ПМИД 10572114 .
- ^ Ву М., Робертс Дж.В., Ким С., Кох Д.Л., ДеЛиза MP (июль 2006 г.). «Коллективная бактериальная динамика выявлена с помощью трехмерного метода отслеживания дефокусированных частиц в масштабе популяции» . Прикладная и экологическая микробиология . 72 (7): 4987–94. Бибкод : 2006ApEnM..72.4987W . дои : 10.1128/АЕМ.00158-06 . ПМЦ 1489374 . ПМИД 16820497 .
- ^ Мэттик Дж.С. (2002). «Пили IV типа и подергивания моторики». Ежегодный обзор микробиологии . 56 : 289–314. дои : 10.1146/annurev.micro.56.012302.160938 . ПМИД 12142488 .
- ^ Мерц А.Дж., Со М., член парламента Шитца (сентябрь 2000 г.). «Втягивание пилуса приводит к подергивающейся подвижности бактерий». Природа . 407 (6800): 98–102. Бибкод : 2000Natur.407...98M . дои : 10.1038/35024105 . ПМИД 10993081 . S2CID 4425775 .
- ^ Люкс Р., Ши В (июль 2004 г.). «Движения бактерий, управляемые хемотаксисом» . Критические обзоры по оральной биологии и медицине . 15 (4): 207–20. дои : 10.1177/154411130401500404 . ПМИД 15284186 .
- ^ Швайницер Т., Йозенханс К. (июль 2010 г.). «Бактериальные энергетические такси: глобальная стратегия?» . Архив микробиологии . 192 (7): 507–20. Бибкод : 2010ArMic.192..507S . дои : 10.1007/s00203-010-0575-7 . ПМЦ 2886117 . ПМИД 20411245 .
- ^ Франкель Р.Б., Базылински Д.А., Джонсон М.С., Тейлор Б.Л. (август 1997 г.). «Магнитоаэротаксис морских кокковидных бактерий» . Биофизический журнал . 73 (2): 994–1000. Бибкод : 1997BpJ....73..994F . дои : 10.1016/S0006-3495(97)78132-3 . ПМК 1180996 . ПМИД 9251816 .
- ^ Нан Б., Зусман Д.Р. (2011). «Раскрытие тайны скользящей подвижности миксобактерий» . Ежегодный обзор генетики . 45 : 21–39. doi : 10.1146/annurev-genet-110410-132547 . ПМЦ 3397683 . ПМИД 21910630 .
- ^ Гольдберг М.Б. (декабрь 2001 г.). «Актиновая подвижность внутриклеточных микробных патогенов» . Обзоры микробиологии и молекулярной биологии . 65 (4): 595–626, оглавление. дои : 10.1128/MMBR.65.4.595-626.2001 . ПМК 99042 . ПМИД 11729265 .
- ^ Кальканьиле М., Тредичи С.М., Тала А., Алифано П. (декабрь 2019 г.). «Бактериальные семиохимические вещества и взаимодействие трансцарства с насекомыми и растениями» . Насекомые . 10 (12): 441. doi : 10.3390/insects10120441 . ПМК 6955855 . ПМИД 31817999 .
- ^ Перейти обратно: а б Шапиро Дж. А. (1998). «Думая о бактериальных популяциях как о многоклеточных организмах» (PDF) . Ежегодный обзор микробиологии . 52 : 81–104. дои : 10.1146/аннурев.микро.52.1.81 . ПМИД 9891794 . Архивировано из оригинала (PDF) 17 июля 2011 года.
- ^ Перейти обратно: а б Костертон Дж.В., Левандовски З., Колдуэлл Д.Е., Корбер Д.Р., Лаппин-Скотт Х.М. (1995). «Микробные биопленки». Ежегодный обзор микробиологии . 49 : 711–45. дои : 10.1146/annurev.mi.49.100195.003431 . ПМИД 8561477 .
- ^ Миллер Д.П., Ламонт Р.Дж. (2019). «Сигнальные системы в бактериях полости рта». Иммунитет слизистой оболочки полости рта и микробиом . Достижения экспериментальной медицины и биологии. Том. 1197. стр. 27–43. дои : 10.1007/978-3-030-28524-1_3 . ISBN 978-3-030-28523-4 . ПМИД 31732932 . S2CID 208063186 .
- ^ Абисадо Р.Г., Беномар С., Клаус-младший, Дандекар А.А., Чендлер-младший (май 2018 г.). «Ощущение бактериального кворума и взаимодействие микробного сообщества» . мБио . 9 (3). дои : 10.1128/mBio.02331-17 . ПМЦ 5964356 . ПМИД 29789364 .
- ^ Миллер М.Б., Басслер Б.Л. (2001). «Кворум-чувство у бактерий». Ежегодный обзор микробиологии . 55 : 165–99. дои : 10.1146/annurev.micro.55.1.165 . ПМИД 11544353 . S2CID 1099089 .
- ^ Чжу К., Май У., Пфайффер В., Янссен С., Асникар Ф., Сандерс Дж.Г. и др. (декабрь 2019 г.). «Филогеномика 10 575 геномов выявила эволюционную близость между доменами бактерий и архей» . Природные коммуникации . 10 (1): 5477. Бибкод : 2019NatCo..10.5477Z . дои : 10.1038/s41467-019-13443-4 . ПМК 6889312 . ПМИД 31792218 .
- ^ Баучер И., Дуади С.Дж., Папке Р.Т., Уолш Д.А., Будро М.Е., Несбё К.Л., Кейс Р.Дж., Дулиттл В.Ф. (2003). «Боковой перенос генов и происхождение прокариотических групп». Ежегодный обзор генетики . 37 : 283–328. дои : 10.1146/annurev.genet.37.050503.084247 . ПМИД 14616063 .
- ^ Олсен Г.Дж., Вёзе Ч.Р., Овербик Р. (январь 1994 г.). «Ветры (эволюционных) перемен: вдохнуть новую жизнь в микробиологию» . Журнал бактериологии . 176 (1): 1–6. дои : 10.2172/205047 . ПМК 205007 . ПМИД 8282683 .
- ^ «Дом ИЖСЭМ» . Ijs.sgmjournals.org. 28 октября 2011 года. Архивировано из оригинала 19 октября 2011 года . Проверено 4 ноября 2011 г.
- ^ «Руководство Берджи» . Bergeys.org. Архивировано из оригинала 7 ноября 2011 года . Проверено 4 ноября 2011 г.
- ^ Хедлунд Б.П., Додсворт Дж.А., Стейли Дж.Т. (июнь 2015 г.). «Изменение ландшафта исследования микробного биоразнообразия и его значение для систематики». Систематическая и прикладная микробиология . 38 (4): 231–36. дои : 10.1016/j.syapm.2015.03.003 . ПМИД 25921438 .
- ^ « Шизомицеты ». Медицинский словарь Мерриам-Вебстера. По состоянию на 3 августа 2021 г.
- ^ Браун М.М., Хорсвилл, Арканзас (ноябрь 2020 г.). «Staphylococcus epidermidis – друг или враг кожи?» . ПЛОС Патогены . 16 (11): e1009026. дои : 10.1371/journal.ppat.1009026 . ПМЦ 7660545 . ПМИД 33180890 .
- ^ Гупта РС (2000). «Естественные эволюционные отношения между прокариотами». Критические обзоры по микробиологии . 26 (2): 111–31. CiteSeerX 10.1.1.496.1356 . дои : 10.1080/10408410091154219 . ПМИД 10890353 . S2CID 30541897 .
- ^ Раппе М.С., Джованнони С.Дж. (2003). «Некультивируемое микробное большинство». Ежегодный обзор микробиологии . 57 : 369–94. дои : 10.1146/annurev.micro.57.030502.090759 . ПМИД 14527284 . S2CID 10781051 .
- ^ Дулиттл РФ (июнь 2005 г.). «Эволюционные аспекты полногеномной биологии». Современное мнение в области структурной биологии . 15 (3): 248–53. дои : 10.1016/j.sbi.2005.04.001 . ПМИД 15963888 .
- ^ Кавалер-Смит Т. (январь 2002 г.). «Неомуранское происхождение архебактерий, негибактериальный корень универсального дерева и бактериальная мегаклассификация» . Международный журнал систематической и эволюционной микробиологии . 52 (Часть 1): 7–76. дои : 10.1099/00207713-52-1-7 . ПМИД 11837318 .
- ^ Поммервилль 2014 , с. 15−31.
- ^ Перейти обратно: а б Краснер 2014 , с. 77.
- ^ Вудс Г.Л., Уокер Д.Х. (июль 1996 г.). «Обнаружение инфекции или инфекционных агентов с помощью цитологического и гистологического окрашивания» . Обзоры клинической микробиологии . 9 (3): 382–404. дои : 10.1128/CMR.9.3.382 . ПМК 172900 . ПМИД 8809467 .
- ^ Тан С., Орси Р.Х., Луо Х., Ге С., Чжан Г., Бейкер Р.К., Стивенсон А., Видманн М. (2019). «Оценка и сравнение методов молекулярного подтипирования и характеристики сальмонеллы» . Границы микробиологии . 10 : 1591. дои : 10.3389/fmicb.2019.01591 . ПМК 6639432 . ПМИД 31354679 .
- ^ Краснер 2014 , с. 87–89.
- ^ Член парламента Вайнштейна (март 1994 г.). «Клиническое значение культур крови». Клиники лабораторной медицины . 14 (1): 9–16. дои : 10.1016/S0272-2712(18)30390-1 . ПМИД 8181237 .
- ^ Райли Л.В. (ноябрь 2018 г.). «Лабораторные методы молекулярной эпидемиологии: бактериальные инфекции». Микробиологический спектр . 6 (6). doi : 10.1128/microbiolspec.AME-0004-2018 . ПМИД 30387415 . S2CID 54234977 .
- ^ Ленковски М., Нияковски К., Качмарек М., Сурдачка А. (март 2021 г.). «Техника петлевой изотермической амплификации в пародонтальной диагностике: систематический обзор» . Журнал клинической медицины . 10 (6): 1189. doi : 10.3390/jcm10061189 . ПМК 8000232 . ПМИД 33809163 .
- ^ Ализаде М., Юсефи Л., Пакдел Ф., Готаслоу Р., Резаи М.А., Ходадади Э., Оскуэй М.А., Соруш Бархаги М.Х., Кафил Х.С. (2021). «Применение масс-спектроскопии MALDI-TOF в клинической микробиологии» . Достижения фармакологических и фармацевтических наук . 2021 : 9928238. doi : 10.1155/2021/9928238 . ПМЦ 8121603 . ПМИД 34041492 .
- ^ Дудек Н.К., Сан С.Л., Бурштейн Д. (2017). «Новое микробное разнообразие и функциональный потенциал микробиома полости рта морских млекопитающих» (PDF) . Современная биология . 27 (24): 3752–62. Бибкод : 2017CBio...27E3752D . дои : 10.1016/j.cub.2017.10.040 . ПМИД 29153320 . S2CID 43864355 .
- ^ Луи М., Луи Л., Симор А.Е. (август 2000 г.). «Роль технологии амплификации ДНК в диагностике инфекционных заболеваний» . CMAJ . 163 (3): 301–09. дои : 10.1016/s1381-1169(00)00220-x . ПМК 80298 . ПМИД 10951731 . Архивировано из оригинала 14 июня 2006 года.
- ^ Оливер Джей Ди (февраль 2005 г.). «Жизнеспособное, но некультивируемое состояние бактерий» . Журнал микробиологии . 43 Спецификация: 93–100. ПМИД 15765062 . Архивировано из оригинала 28 сентября 2007 года.
- ^ Эзеби JP (8 декабря 2011 г.). «Количество опубликованных имен» . Список названий прокариот, стоящих в номенклатуре . Архивировано из оригинала 19 января 2012 года . Проверено 10 декабря 2011 г.
- ^ Кертис Т.П., Слоан В.Т., Сканнелл Дж.В. (август 2002 г.). «Оценка прокариотического разнообразия и его пределы» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (16): 10494–99. Бибкод : 2002PNAS...9910494C . дои : 10.1073/pnas.142680199 . ПМК 124953 . ПМИД 12097644 .
- ^ Шлосс П.Д., Хандельсман Дж. (декабрь 2004 г.). «Состояние микробной переписи» . Обзоры микробиологии и молекулярной биологии . 68 (4): 686–91. дои : 10.1128/MMBR.68.4.686-691.2004 . ПМК 539005 . ПМИД 15590780 .
- ^ Орен А., генеральный менеджер Гаррити (2021 г.). «Действительная публикация названий сорока двух типов прокариот» . Int J Syst Evol Microbiol . 71 (10): 5056. doi : 10.1099/ijsem.0.005056 . ПМИД 34694987 . S2CID 239887308 .
- ^ Фишер Б., Харви Р.П., Чамп ПК (2007). «Глава 33». Иллюстрированные обзоры Липпинкотта: микробиология (серия иллюстрированных обзоров Липпинкотта) . Хагерствон, доктор медицины: Липпинкотт Уильямс и Уилкинс. стр. 367–92. ISBN 978-0-7817-8215-9 .
- ^ Мушегян А.А., Эберт Д. (январь 2016 г.). «Переосмысление «мутуализма» в различных сообществах хозяев-симбионтов». Биоэссе . 38 (1): 100–8. doi : 10.1002/bies.201500074 . ПМИД 26568407 . S2CID 31661712 .
- ^ Харпер Д. «Комменсализм» . Интернет-словарь этимологии .
- ^ Sears CL (октябрь 2005 г.). «Динамическое партнерство: прославление нашей кишечной флоры». Анаэроб . 11 (5): 247–51. дои : 10.1016/j.anaerobe.2005.05.001 . ПМИД 16701579 .
- ^ Хан Р., Петерсен ФК, Шекхар С. (2019). «Комменсальные бактерии: новый игрок в защите от респираторных патогенов» . Границы в иммунологии . 10 : 1203. дои : 10.3389/fimmu.2019.01203 . ПМК 6554327 . ПМИД 31214175 .
- ^ Роско Д.Л., Чоу А.В. (март 1988 г.). «Нормальная флора и иммунитет слизистых оболочек головы и шеи». Клиники инфекционных заболеваний Северной Америки . 2 (1): 1–19. дои : 10.1016/S0891-5520(20)30163-X . ПМИД 3074102 .
- ^ Шиао С.Л., Кершоу К.М., Лимон Дж.Дж., Ю С., Юн Дж., Ко Е.Ю., Гуарнерио Дж., Потдар А.А., Макговерн Д.П., Бозе С., Дар ТБ, Ноэ П., Ли Дж., Кубота Ю., Майми В.И., Дэвис М.Дж., Хенсон Р.М. , Чой Р.Ю., Ян В., Тан Дж., Гаргус М., Принс А.Д., Зумстег З.С., Андерхилл Д.М. (июль 2021 г.). «Комменсальные бактерии и грибы по-разному регулируют реакцию опухоли на лучевую терапию» . Раковая клетка . 39 (9): 1202–1213.e6. doi : 10.1016/j.ccell.2021.07.002 . ПМЦ 8830498 . ПМИД 34329585 .
- ^ Ляпичев К.А., Ивашкевич Ю., Чернов Ю., Чиненов Д., Шпот Е., Бессонов А.А., Дабая Б.С., Коноплев С. (2021). «MALT-лимфома мочевого пузыря демонстрирует резкое преобладание женщин, неравномерное географическое распространение и возможную инфекционную этиологию» . Исследования и отчеты в области урологии . 13 : 49–62. дои : 10.2147/RRU.S283366 . ПМЦ 7873029 . ПМИД 33575225 .
- ^ Элеян Л., Хан А.А., Мусоллари Г., Чандирамани А.С., Шейх С., Салха А., Тармахомед А., Харки А. (апрель 2021 г.). «Инфекционный эндокардит в детской популяции». Европейский журнал педиатрии . 180 (10): 3089–3100. дои : 10.1007/s00431-021-04062-7 . ПМИД 33852085 . S2CID 233225250 .
- ^ Мартин М.О. (сентябрь 2002 г.). «Хищные прокариоты: новые возможности для исследований». Журнал молекулярной микробиологии и биотехнологии . 4 (5): 467–77. ПМИД 12432957 .
- ^ Велисер Г.Дж., Стредвик К.Л. (август 2002 г.). «Экспериментальная социальная эволюция с Myxococcus xanthus». Антони ван Левенгук . 81 (1–4): 155–64. дои : 10.1023/А:1020546130033 . ПМИД 12448714 . S2CID 20018104 .
- ^ Бауэр А., Форххаммер К. (май 2021 г.). «Бактериальное хищничество цианобактерий» . Микробная физиология . 31 (2): 99–108. дои : 10.1159/000516427 . ISSN 2673-1665 . ПМИД 34010833 .
- ^ Велисер Г.Дж., Мендес-Соареш Х. (январь 2009 г.). «Бактериальные хищники» . Современная биология . 19 (2): R55–56. Бибкод : 2009CBio...19..R55V . дои : 10.1016/j.cub.2008.10.043 . ПМИД 19174136 . S2CID 5432036 .
- ^ Стамс А.Дж., де Бок Ф.А., Плугге СМ, ван Экерт М.Х., Дольфинг Дж., Шраа Г. (март 2006 г.). «Экзоклеточный перенос электронов в анаэробных микробных сообществах». Экологическая микробиология . 8 (3): 371–82. Бибкод : 2006EnvMi...8..371S . дои : 10.1111/j.1462-2920.2006.00989.x . ПМИД 16478444 .
- ^ Зинзер Э.Р. (август 2018 г.). «Перекрестная защита от перекиси водорода с помощью микробов-помощников: воздействие на цианобактерию Prochromococcus и других бенефициаров морских сообществ» . Отчеты по экологической микробиологии . 10 (4): 399–411. Бибкод : 2018EnvMR..10..399Z . дои : 10.1111/1758-2229.12625 . ПМИД 29411546 .
- ^ Бареа Х.М., Посо М.Дж., Азкон Р., Азкон-Агилар К. (июль 2005 г.). «Микробное сотрудничество в ризосфере» . Журнал экспериментальной ботаники . 56 (417): 1761–78. дои : 10.1093/jxb/eri197 . ПМИД 15911555 .
- ^ О'Хара А.М., Шанахан Ф. (июль 2006 г.). «Кишечная флора как забытый орган» . Отчеты ЭМБО . 7 (7): 688–93. дои : 10.1038/sj.embor.7400731 . ПМК 1500832 . ПМИД 16819463 .
- ^ Зотендал Э.Г., Воан Э.Э., де Вос В.М. (март 2006 г.). «Микробный мир внутри нас» . Молекулярная микробиология . 59 (6): 1639–50. дои : 10.1111/j.1365-2958.2006.05056.x . ПМИД 16553872 . S2CID 37602619 .
- ^ Горбач С.Л. (февраль 1990 г.). «Молочнокислые бактерии и здоровье человека». Анналы медицины . 22 (1): 37–41. дои : 10.3109/07853899009147239 . ПМИД 2109988 .
- ^ Салминен С.Дж., Геймонд М., Исолаури Э. (май 2005 г.). «Пробиотики, которые изменяют риск заболеваний» . Журнал питания . 135 (5): 1294–98. дои : 10.1093/jn/135.5.1294 . ПМИД 15867327 .
- ^ Ватанабэ Ф., Бито Т. (январь 2018 г.). «Источники витамина B12 и микробное взаимодействие» . Экспериментальная биология и медицина . 243 (2): 148–58. дои : 10.1177/1535370217746612 . ПМК 5788147 . ПМИД 29216732 .
- ^ Поммервилль, 2014 , стр. 16–21.
- ^ Кларк 2010 , с. 215.
- ^ Уилис 2008 , с. 44.
- ^ Кларк 2010 , стр. 30, 195, 233, 236.
- ^ Миллер А.К., Уильямс С.М. (июль 2021 г.). «Инфекция Helicobacter pylori оказывает как защитное, так и вредное воздействие на здоровье и болезни человека» . Гены и иммунитет . 22 (4): 218–226. дои : 10.1038/s41435-021-00146-4 . ПМЦ 8390445 . ПМИД 34244666 .
- ^ Шварц С., Энне VI, ван Дуйкерен Э. (октябрь 2016 г.). «40 лет ветеринарных работ в JAC – чему мы научились?» . Журнал антимикробной химиотерапии . 71 (10): 2681–90. дои : 10.1093/jac/dkw363 . ПМИД 27660260 .
- ^ Фиш Д.Н. (февраль 2002 г.). «Оптимальная противомикробная терапия сепсиса» . Американский журнал аптеки системы здравоохранения . 59 (Приложение 1): С13–19. дои : 10.1093/ajhp/59.suppl_1.S13 . ПМИД 11885408 .
- ^ Белланд Р.Дж., Уэллетт С.П., Гифферс Дж., Бирн Дж.И. (февраль 2004 г.). «Хламидия пневмония и атеросклероз» . Клеточная микробиология . 6 (2): 117–27. дои : 10.1046/j.1462-5822.2003.00352.x . ПМИД 14706098 . S2CID 45218449 .
- ^ Хейзе Э.Р. (февраль 1982 г.). «Заболевания, связанные с иммуносупрессией» . Перспективы гигиены окружающей среды . 43 : 9–19. дои : 10.2307/3429162 . JSTOR 3429162 . ПМЦ 1568899 . ПМИД 7037390 .
- ^ Сайман Л (2004). «Микробиология ранних заболеваний легких МВ». Обзоры детских респираторных заболеваний . 5 (Приложение А): S367–69. дои : 10.1016/S1526-0542(04)90065-6 . ПМИД 14980298 .
- ^ Поммервилль 2014 , с. 118.
- ^ Перейти обратно: а б Поммервилль, 2014 г. , стр. 646–47.
- ^ Краснер 2014 , стр. 165, 369.
- ^ Йонат А., Башан А. (2004). «Рибосомальная кристаллография: инициация, образование пептидных связей и полимеризация аминокислот затрудняются антибиотиками» . Ежегодный обзор микробиологии . 58 : 233–51. дои : 10.1146/annurev.micro.58.030603.123822 . ПМИД 15487937 .
- ^ Хачатурян Г.Г. (ноябрь 1998 г.). «Использование антибиотиков в сельском хозяйстве, эволюция и перенос бактерий, устойчивых к антибиотикам» . CMAJ . 159 (9): 1129–36. ПМЦ 1229782 . ПМИД 9835883 .
- ^ Куо Дж (октябрь 2017 г.). «Процессы дезинфекции» . Исследования водной среды . 89 (10): 1206–44. Бибкод : 2017WaEnR..89.1206K . дои : 10.2175/106143017X15023776270278 . ПМИД 28954657 .
- ^ Джонсон М.Э., Люси Дж.А. (апрель 2006 г.). «Основные технологические достижения и тенденции в области сыра» . Журнал молочной науки . 89 (4): 1174–78. doi : 10.3168/jds.S0022-0302(06)72186-5 . ПМИД 16537950 .
- ^ Краснер 2014 , стр. 25–26.
- ^ Коэн Ю. (декабрь 2002 г.). «Биоремедиация нефти морскими микробными матами» . Международная микробиология . 5 (4): 189–93. дои : 10.1007/s10123-002-0089-5 . ПМИД 12497184 . S2CID 26039323 .
- ^ Невес Л.К., Миямура Т.Т., Мораес Д.А., Пенна Т.К., Конверти А. (2006). «Методы биофильтрации для удаления фенольных остатков». Прикладная биохимия и биотехнология . 129–132 (1–3): 130–52. дои : 10.1385/ABAB:129:1:130 . ПМИД 16915636 . S2CID 189905816 .
- ^ Лизе А., Фильо М.В. (декабрь 1999 г.). «Производство продуктов тонкой химии с использованием биокатализа». Современное мнение в области биотехнологии . 10 (6): 595–603. дои : 10.1016/S0958-1669(99)00040-3 . ПМИД 10600695 .
- ^ Аронсон А.И., Шай Ю. (февраль 2001 г.). «Почему инсектицидные токсины Bacillus thuringiensis так эффективны: уникальные особенности их действия» . Письма FEMS по микробиологии . 195 (1): 1–8. дои : 10.1111/j.1574-6968.2001.tb10489.x . ПМИД 11166987 .
- ^ Божик А (июль 2006 г.). «Восприимчивость взрослых Coccinella septempunctata (Coleoptera: Coccinellidae) к инсектицидам с различным механизмом действия». Наука борьбы с вредителями . 62 (7): 651–54. дои : 10.1002/ps.1221 . ПМИД 16649191 .
- ^ Чаттопадхьяй А., Бхатнагар Н.Б., Бхатнагар Р. (2004). «Бактериальные инсектицидные токсины». Критические обзоры по микробиологии . 30 (1): 33–54. дои : 10.1080/10408410490270712 . ПМИД 15116762 . S2CID 1580984 .
- ^ Серрес М.Х., Гопал С., Нахум Л.А., Лян П., Гаастерланд Т., Райли М. (2001). «Функциональное обновление генома Escherichia coli K-12» . Геномная биология . 2 (9): ИССЛЕДОВАНИЕ0035. doi : 10.1186/gb-2001-2-9-research0035 . ПМК 56896 . ПМИД 11574054 .
- ^ Алмаас Э., Ковач Б., Вичек Т., Олваи З.Н., Барабаши А.Л. (февраль 2004 г.). «Глобальная организация метаболических потоков у бактерии Escherichia coli». Природа . 427 (6977): 839–43. arXiv : q-bio/0403001 . Бибкод : 2004Natur.427..839A . дои : 10.1038/nature02289 . ПМИД 14985762 . S2CID 715721 .
- ^ Рид Дж.Л., Во Т.Д., Шиллинг Ч., Палссон Б.О. (2003). «Модель расширенного генома Escherichia coli K-12 (iJR904 GSM/GPR)» . Геномная биология . 4 (9): Р54. дои : 10.1186/gb-2003-4-9-r54 . ЧВК 193654 . ПМИД 12952533 .
- ^ Уолш Дж. (апрель 2005 г.). «Терапевтические инсулины и их крупнотоннажное производство». Прикладная микробиология и биотехнология . 67 (2): 151–59. дои : 10.1007/s00253-004-1809-x . ПМИД 15580495 . S2CID 5986035 .
- ^ Грауманн К., Премсталлер А (февраль 2006 г.). «Производство рекомбинантных терапевтических белков в микробных системах». Биотехнологический журнал . 1 (2): 164–86. дои : 10.1002/biot.200500051 . ПМИД 16892246 . S2CID 24702839 .
- ^ Рабш В., Хельм Р.А., Эйзенштарк А. (февраль 2004 г.). «Разнообразие типов фагов среди архивных культур Демерецкой коллекции штаммов серовара Salmonella enterica Typhimurium» . Прикладная и экологическая микробиология . 70 (2): 664–69. Бибкод : 2004ApEnM..70..664R . doi : 10.1128/aem.70.2.664-669.2004 . ПМЦ 348941 . ПМИД 14766539 .
- ^ Уилис 2008 .
- ^ Азимов I (1982). Биографическая энциклопедия науки и техники Азимова (2-е изд.). Гарден-Сити, Нью-Йорк: Doubleday and Company. п. 143.
- ^ Поммервилль 2014 , с. 7.
- ^ Эренберг К.Г. (1828). Символы Физио. Животные вечнопозвоночные . Берлин: Декас прима.
- ^ Порода RS, Conn HJ (май 1936 г.). «Статус общего термина Bacterium Ehrenberg 1828» . Журнал бактериологии . 31 (5): 517–18. дои : 10.1128/jb.31.5.517-518.1936 . ПМЦ 543738 . ПМИД 16559906 .
- ^ Эренберг К.Г. (1835). вклад в знание великой организации в направлении наименьшего пространства ( Третий на немецком языке). Берлин: Физические трактаты Королевской академии наук. стр. 143–336.
- ^ «Документы Пастера по теории микробов» . Юридический сайт юридического центра LSU в области медицины и общественного здравоохранения, исторические статьи об общественном здравоохранении. Архивировано из оригинала 18 декабря 2006 года . Проверено 23 ноября 2006 г.
- ^ « Раньше «мыть руки» было спорным медицинским советом» . История . 6 марта 2020 г. Архивировано из оригинала 11 августа 2023 г.
- ^ «Нобелевская премия по физиологии и медицине 1905 года» . Нобелевская премия.org. Архивировано из оригинала 10 декабря 2006 года . Проверено 22 ноября 2006 г.
- ^ О'Брайен С.Дж., Годерт Дж.Дж. (октябрь 1996 г.). «ВИЧ вызывает СПИД: постулаты Коха сбылись» . Современное мнение в иммунологии . 8 (5): 613–18. дои : 10.1016/S0952-7915(96)80075-6 . ПМИД 8902385 .
- ^ Чунг КТ. «Фердинанд Юлиус Кон (1828–1898): пионер бактериологии» (PDF) . Кафедра микробиологии и молекулярно-клеточных наук Университета Мемфиса. Архивировано (PDF) из оригинала 27 июля 2011 года.
- ^ Дрюс, Герхарт (1999). «Фердинанд Кон, основатель современной микробиологии» (PDF) . Новости АСМ . 65 (8): 547–52. Архивировано из оригинала (PDF) 13 июля 2017 года.
- ^ Терстон Эй Джей (декабрь 2000 г.). «О крови, воспалении и огнестрельных ранениях: история борьбы с сепсисом» . Австралийский и новозеландский журнал хирургии . 70 (12): 855–61. дои : 10.1046/j.1440-1622.2000.01983.x . ПМИД 11167573 .
- ^ Шварц Р.С. (март 2004 г.). «Волшебные пули Пауля Эрлиха». Медицинский журнал Новой Англии . 350 (11): 1079–80. дои : 10.1056/NEJMp048021 . ПМИД 15014180 .
- ^ «Жизнеописание Пауля Эрлиха» . Нобелевская премия.org. Архивировано из оригинала 28 ноября 2006 года . Проверено 26 ноября 2006 г.
- ^ Woese CR, Fox GE (ноябрь 1977 г.). «Филогенетическая структура прокариотического домена: первичные царства» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (11): 5088–90. Бибкод : 1977PNAS...74.5088W . дои : 10.1073/pnas.74.11.5088 . ПМК 432104 . ПМИД 270744 .
Библиография
- Кларк Д. (2010). Микробы, гены и цивилизация: как эпидемии сформировали то, кем мы являемся сегодня . Река Аппер-Седл, Нью-Джерси: FT Press. ISBN 978-0-13-701996-0 . OCLC 473120711 .
- Кроуфорд Д. (2007). Смертельные спутники: как микробы сформировали нашу историю . Оксфорд, Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-956144-5 . OCLC 183198723 .
- Зал Б (2008 г.). Эволюция Стрикбергера: интеграция генов, организмов и популяций . Садбери, Массачусетс: Джонс и Бартлетт. ISBN 978-0-7637-0066-9 . OCLC 85814089 .
- Краснер Р. (2014). Микробная проблема: взгляд на общественное здравоохранение . Берлингтон, Массачусетс: Jones & Bartlett Learning. ISBN 978-1-4496-7375-8 . OCLC 794228026 .
- Поммервиль Дж. К. (2014). Основы микробиологии (10-е изд.). Бостон: Джонс и Бартлетт. ISBN 978-1-284-03968-9 .
- Уилис М (2008). Основы современной микробиологии . Садбери, Массачусетс: Издательство Jones and Bartlett. ISBN 978-0-7637-1075-0 . OCLC 67392796 .
