Облигатные анаэробы
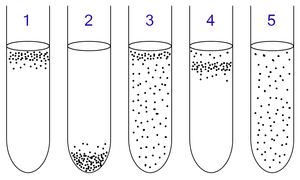
1: Облигатным аэробам необходим кислород, поскольку они не могут ферментировать или дышать анаэробно. Они собираются в верхней части трубки, где концентрация кислорода самая высокая.
2: Облигатные анаэробы отравлены кислородом, поэтому они собираются на дне пробирки, где концентрация кислорода наименьшая.
3. Факультативные анаэробы могут расти как с кислородом, так и без него, поскольку они могут метаболизировать энергию аэробно или анаэробно. Они собираются в основном вверху, потому что аэробное дыхание генерирует больше АТФ, чем ферментация или анаэробное дыхание.
4. Микроаэрофилам необходим кислород, поскольку они не могут ферментировать или дышать анаэробно. Однако они отравлены высокими концентрациями кислорода. Они собираются в верхней части пробирки, а не на самом верху.
5. Аэротолерантные организмы не нуждаются в кислороде и не могут его использовать, даже если он присутствует; они метаболизируют энергию анаэробно. Однако в отличие от облигатных анаэробов они не отравляются кислородом. Их можно найти равномерно распределенными по всей пробирке.
И факультативные анаэробы, и аэротолерантные организмы подвергаются ферментации в отсутствие кислорода, но факультативные анаэробы переключаются на аэробный метаболизм в присутствии кислорода (феномен, известный как эффект Пастера ). Эффект Пастера иногда используется в лаборатории для различения факультативных анаэробов и аэротолерантных организмов.
Облигатные анаэробы – это микроорганизмы , погибающие при нормальной атмосферной концентрации кислорода (20,95% O 2 ). [1] [2] Толерантность к кислороду варьируется у разных видов: некоторые виды способны выживать при содержании кислорода до 8%, тогда как другие теряют жизнеспособность в среде с концентрацией кислорода более 0,5%. [3]
Чувствительность к кислороду
[ редактировать ]Чувствительность к кислороду облигатных анаэробов объясняется сочетанием факторов, включая окислительный стресс и выработку ферментов. Кислород также может повреждать облигатные анаэробы способами, не связанными с окислительным стрессом. [ нужна ссылка ]
Поскольку молекулярный кислород содержит два неспаренных электрона на высшей занятой молекулярной орбитали , он легко восстанавливается до супероксида ( O −
2 ) и перекись водорода ( H
22О
2 ) внутри клеток. [1] Реакция между этими двумя продуктами приводит к образованию свободного гидроксильного радикала ( OH . ). [4] Супероксид, перекись водорода и гидроксильные радикалы представляют собой класс соединений, известных как активные формы кислорода (АФК), высокореактивные продукты, которые повреждают микробы, включая облигатные анаэробы. [4] Аэробные организмы производят супероксиддисмутазу и каталазу для детоксикации этих продуктов, но облигатные анаэробы производят эти ферменты в очень небольших количествах или не производят вообще. [1] [2] [3] [5] Считается , что вариабельность толерантности к кислороду облигатных анаэробов (от <0,5 до 8% O 2 ) отражает количество вырабатываемых супероксиддисмутазы и каталазы. [2] [3]
В 1986 году Карлиоз и Туати провели эксперименты, которые подтвердили идею о том, что активные формы кислорода могут быть токсичными для анаэробов. E. coli , факультативный анаэроб, была мутирована путем делеции генов супероксиддисмутазы. В присутствии кислорода эта мутация приводила к неспособности правильно синтезировать определенные аминокислоты или использовать обычные источники углерода в качестве субстратов во время метаболизма. [6] В отсутствие кислорода мутировавшие образцы росли нормально. [6] В 2018 году Лу и др. обнаружили, что у Bacteroides thetaiotaomicron , облигатного анаэроба, обнаруженного в пищеварительном тракте млекопитающих, воздействие кислорода приводит к повышению уровня супероксида, который инактивирует важные метаболические ферменты. [6]
Растворенный кислород увеличивает окислительно-восстановительный потенциал раствора, а высокий окислительно-восстановительный потенциал подавляет рост некоторых облигатных анаэробов. [3] [5] [7] Например, метаногены растут при окислительно-восстановительном потенциале ниже -0,3 В. [7] Сульфид является важным компонентом некоторых ферментов, а молекулярный кислород окисляет его с образованием дисульфида , тем самым инактивируя определенные ферменты (например, нитрогеназу ). Организмы могут оказаться неспособными расти, если эти важные ферменты деактивированы. [1] [5] [7] Рост также может быть ингибирован из-за отсутствия восстанавливающих эквивалентов для биосинтеза, поскольку электроны истощаются при восстановлении кислорода. [7]
Энергетический обмен
[ редактировать ]Облигатные анаэробы преобразуют питательные вещества в энергию посредством анаэробного дыхания или ферментации . При аэробном дыхании пируват, образующийся в результате гликолиза, превращается в ацетил-КоА . Затем он расщепляется через цикл ТСА и цепь переноса электронов . Анаэробное дыхание отличается от аэробного дыхания тем, что электронов при нем используется не кислород, а акцептор в цепи переноса электронов. Примеры альтернативных акцепторов электронов включают сульфат , нитрат , железо , марганец , ртуть и окись углерода . [8]
Ферментация отличается от анаэробного дыхания тем, что пируват, образующийся в результате гликолиза, расщепляется без участия цепи переноса электронов (т. е. отсутствует окислительное фосфорилирование ). Существуют многочисленные пути ферментации, такие как молочнокислое брожение , смешаннокислое брожение , 2-3-бутандиоловое брожение , при котором органические соединения восстанавливаются до органических кислот и спирта. [8] [4]
Энергетический выход анаэробного дыхания и ферментации (т.е. количество образующихся молекул АТФ ) меньше, чем при аэробном дыхании. [8] Вот почему факультативные анаэробы , которые могут метаболизировать энергию как аэробно, так и анаэробно, предпочитают метаболизировать энергию аэробно. Это наблюдается при культивировании факультативных анаэробов в тиогликолятном бульоне . [1]
Экология и примеры
[ редактировать ]Облигатные анаэробы встречаются в бескислородных средах, таких как кишечный тракт животных, глубины океана, стоячие воды, свалки, в глубоких отложениях почвы. [9] Примеры облигатно анаэробных бактерий родов включают Actinomyces , Bacteroides , Clostridium , Fusobacterium , Peptostreptococcus , Porphyromonas , Prevotella , Propionibacterium и Veillonella . Clostridium Виды представляют собой бактерии, образующие эндоспоры , и в этой спящей форме могут выживать при атмосферных концентрациях кислорода. Остальные перечисленные бактерии не образуют эндоспор. [5]
Несколько видов родов Mycobacterium , Streptomyces и Rhodococcus являются примерами облигатных анаэробов, обнаруженных в почве. [10] Облигатные анаэробы обнаруживаются также в пищеварительном тракте человека и других животных, а также в первом желудке жвачных животных . [11]
Примеры облигатно анаэробных родов грибов включают рубцовые грибы Neocallimastix , Piromonas и Sphaeromonas . [12]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и Прескотт, Лансинг М.; Харли, Джон П.; Кляйн, Дэвид А. (1996). Микробиология (3-е изд.). Паб Уильяма Си Брауна. стр. 130–131. ISBN 0-697-29390-4 .
- ^ Jump up to: а б с Брукс, Гео Ф.; Кэрролл, Карен С.; Бутель, Джанет С; Морс, Стивен А. (2007). Медицинская микробиология Джаветца, Мельника и Адельберга (24-е изд.). МакГроу Хилл. стр. 307–312. ISBN 978-0-07-128735-7 .
- ^ Jump up to: а б с д Райан, Кеннет Дж.; Рэй, К. Джордж, ред. (2004). Медицинская микробиология Шерриса (4-е изд.). МакГроу Хилл. стр. 309–326, 378–384. ISBN 0-8385-8529-9 .
- ^ Jump up to: а б с Хентгес, Дэвид Дж. (1996), Бэрон, Сэмюэл (редактор), «Анаэробы: общие характеристики» , Медицинская микробиология (4-е изд.), Галвестон (Техас): Медицинское отделение Техасского университета в Галвестоне, ISBN 978-0-9631172-1-2 , PMID 21413255 , получено 26 апреля 2021 г.
- ^ Jump up to: а б с д Левинсон, В. (2010). Обзор медицинской микробиологии и иммунологии (11-е изд.). МакГроу-Хилл. стр. 91–178. ISBN 978-0-07-174268-9 .
- ^ Jump up to: а б с Лу, Чжэн; Сету, Рамакришнан; Имлей, Джеймс А. (3 апреля 2018 г.). «Эндогенный супероксид является ключевым эффектором чувствительности к кислороду модельного облигатного анаэроба» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (14): Е3266–Е3275. дои : 10.1073/pnas.1800120115 . ISSN 0027-8424 . ПМЦ 5889672 . ПМИД 29559534 .
- ^ Jump up to: а б с д Ким, Бён Хон; Гэдд, Джеффри Майкл (2008). Бактериальная физиология и метаболизм . Издательство Кембриджского университета. дои : 10.1017/CBO9780511790461 . ISBN 9780511790461 .
- ^ Jump up to: а б с Хогг, Стюарт (2005). Основная микробиология (1-е изд.). Уайли. стр. 99–100, 118–148. ISBN 0-471-49754-1 .
- ^ «Потребность в кислороде для роста микробов | Микробиология» . Courses.lumenlearning.com . Проверено 8 мая 2021 г.
- ^ Берни, Майкл; Грининг, Крис; Конрад, Ральф; Джейкобс, Уильям Р.; Кук, Грегори М. (2014). «Облигатно аэробная почвенная бактерия активирует ферментативное производство водорода, чтобы пережить восстановительный стресс во время гипоксии» . Труды Национальной академии наук . 111 (31): 11479–11484. дои : 10.1073/pnas.1407034111 . ISSN 0027-8424 . ПМК 4128101 . ПМИД 25049411 .
- ^ Уэки, Ацуко; Каку, Нобуо; Уэки, Кацудзи (01 августа 2018 г.). «Роль анаэробных бактерий в биологической дезинсекции почвы для уничтожения почвенных патогенов растений в сельском хозяйстве». Прикладная микробиология и биотехнология . 102 (15): 6309–6318. дои : 10.1007/s00253-018-9119-x . ISSN 1432-0614 . ПМИД 29858952 . S2CID 44123873 .
- ^ Карлайл, Майкл Дж.; Уоткинсон, Сара К. (1994). Грибы . Академическая пресса. стр. 33–34. ISBN 0-12-159960-4 .
| Медицинский микробиология | |||||||
|---|---|---|---|---|---|---|---|
| Биохимия и экология |
| ||||||
| Форма | |||||||
| Структура |
| ||||||
| Таксономия и эволюция | |||||||