Окислительное фосфорилирование
Окислительное фосфорилирование (Великобритания / ɒ k ˈ s ɪ d . ə . t ɪ v / , США / ˈ ɑː k . s ɪ ˌ d eɪ . t ɪ v / [1] ) или фосфорилирование, связанное с транспортом электронов , или терминальное окисление — это метаболический путь , в котором клетки используют ферменты для окисления питательных веществ , тем самым высвобождая химическую энергию для производства аденозинтрифосфата (АТФ). У эукариот это происходит внутри митохондрий . Почти все аэробные организмы осуществляют окислительное фосфорилирование. Этот путь настолько распространен, потому что он высвобождает больше энергии, чем альтернативные процессы ферментации , такие как анаэробный гликолиз .
Энергия, запасенная в химических связях глюкозы, высвобождается клеткой в цикле лимонной кислоты , производя углекислый газ и энергичных доноров электронов НАДН и ФАДН . Окислительное фосфорилирование использует эти молекулы и O 2 для производства АТФ , который используется в клетке всякий раз, когда необходима энергия. Во время окислительного фосфорилирования электроны передаются от доноров электронов к ряду акцепторов электронов в серии окислительно-восстановительных реакций, заканчивающихся кислородом, реакция которого высвобождает половину общей энергии. [2]
У эукариот эти окислительно-восстановительные реакции катализируются рядом белковых комплексов внутри внутренней мембраны митохондрий клетки, тогда как у прокариот эти белки расположены во внешней мембране клетки. Эти связанные наборы белков называются цепью переноса электронов . У эукариот задействованы пять основных белковых комплексов, тогда как у прокариот присутствует множество различных ферментов, использующих множество доноров и акцепторов электронов.
Энергия, передаваемая электронами, протекающими через эту цепь переноса электронов, используется для транспортировки протонов через внутреннюю мембрану митохондрий в процессе, называемом переносом электронов . Это генерирует потенциальную энергию в виде градиента pH и результирующего электрического потенциала на мембране. Этот запас энергии используется, когда протоны текут обратно через мембрану и вниз по градиенту потенциальной энергии через большой фермент, называемый АТФ-синтазой, в процессе, называемом хемиосмосом . АТФ-синтаза использует энергию для преобразования аденозиндифосфата (АДФ) в аденозинтрифосфат в реакции фосфорилирования . Реакция осуществляется потоком протонов, который заставляет вращаться часть фермента. АТФ-синтаза представляет собой вращательный механический двигатель.
Хотя окислительное фосфорилирование является жизненно важной частью метаболизма, оно производит активные формы кислорода , такие как супероксид и перекись водорода , которые приводят к распространению свободных радикалов , повреждая клетки и способствуя болезням и, возможно, старению и старению . Ферменты, осуществляющие этот метаболический путь, также являются мишенью многих лекарств и ядов, подавляющих их активность.
Хемиосмос [ править ]
Окислительное фосфорилирование основано на использовании химических реакций, выделяющих энергию , для запуска энергозатратных реакций. Говорят, что эти два набора реакций связаны . Это означает, что одно не может происходить без другого. Цепь окислительно-восстановительных реакций, управляющая потоком электронов через цепь переноса электронов от доноров электронов, таких как НАДН , к акцепторам электронов, таким как кислород и водород (протоны), представляет собой экзергонический процесс – он высвобождает энергию, тогда как синтез АТФ представляет собой эндергонический процесс, требующий затрат энергии. И цепь переноса электронов, и АТФ-синтаза встроены в мембрану, и энергия передается от цепи переноса электронов к АТФ-синтазе за счет движения протонов через эту мембрану в процессе, называемом хемиосмосом . [3] Ток протонов перемещается от отрицательной N-стороны мембраны к положительной P-стороне посредством ферментов, перекачивающих протоны цепи переноса электронов. Движение протонов создает электрохимический градиент на мембране, называемый протондвижущей силой . Он состоит из двух компонентов: разницы в концентрации протонов (H + градиент, Δ pH ) и разность электрических потенциалов , при этом N-сторона имеет отрицательный заряд. [4]
АТФ-синтаза высвобождает эту накопленную энергию, замыкая цепь и позволяя протонам течь по электрохимическому градиенту обратно на N-сторону мембраны. [5] Электрохимический градиент приводит в движение часть структуры фермента и связывает это движение с синтезом АТФ.
Две составляющие протондвижущей силы термодинамически эквивалентны: в митохондриях большая часть энергии обеспечивается потенциалом; у алкалофильных бактерий электрическая энергия вынуждена даже компенсировать противодействующую обратную разницу pH. И наоборот, хлоропласты работают в основном за счет ΔpH. Однако им также требуется небольшой мембранный потенциал для кинетики синтеза АТФ. В случае фузобактерии Propionigenium Modetum он вызывает встречное вращение субъединиц a и c мотора F O АТФ-синтазы. [4]
Количество энергии, выделяемой при окислительном фосфорилировании, велико по сравнению с количеством, вырабатываемым при анаэробном брожении . При гликолизе образуются только 2 молекулы АТФ, но где-то от 30 до 36 АТФ образуются в результате окислительного фосфорилирования 10 молекул НАДН и 2 молекул сукцината, образующихся в результате превращения одной молекулы глюкозы в углекислый газ и воду. [6] при этом каждый цикл бета-окисления дает жирной кислоты около 14 АТФ. Эти выходы АТФ представляют собой теоретические максимальные значения; на практике некоторые протоны просачиваются через мембрану, снижая выход АТФ. [7]
Молекулы-переносчики электронов и протонов [ править ]
Цепь переноса электронов переносит как протоны, так и электроны, передавая электроны от доноров к акцепторам и транспортируя протоны через мембрану. В этих процессах используются как растворимые, так и связанные с белками молекулы-переносчики. В митохондриях электроны передаются в межмембранном пространстве с помощью водорастворимого белка -переносчика электронов цитохрома с . [8] Он переносит только электроны, и они передаются за счет восстановления и окисления атома железа , который белок удерживает в гемовой группе в своей структуре. Цитохром С также обнаружен у некоторых бактерий, где он расположен в периплазматическом пространстве . [9]
Внутри внутренней мембраны митохондрий жирорастворимый переносчик электронов кофермент Q10 (Q) переносит как электроны, так и протоны посредством окислительно-восстановительного цикла. [10] Эта небольшая молекула бензохинона очень гидрофобна , поэтому она свободно диффундирует внутри мембраны. Когда Q принимает два электрона и два протона, он восстанавливается до формы убихинола (QH 2 ); когда QH 2 высвобождает два электрона и два протона, он снова окисляется до формы убихинона (Q). В результате, если два фермента расположены так, что Q восстанавливается на одной стороне мембраны, а QH 2 окисляется на другой, убихинон будет связывать эти реакции и переносить протоны через мембрану. [11] В некоторых бактериальных цепях переноса электронов используются различные хиноны, такие как менахинон . помимо убихинона [12]
Внутри белков электроны передаются между флавина . кофакторами [5] [13] железо-серные кластеры и цитохромы. Существует несколько типов железо-серного кластера. Самый простой тип цепи переноса электрона состоит из двух атомов железа, соединенных двумя атомами неорганической серы ; они называются кластерами [2Fe–2S]. Второй вид, названный [4Fe–4S], содержит куб из четырех атомов железа и четырех атомов серы. Каждый атом железа в этих кластерах координируется дополнительной аминокислотой , обычно атомом серы цистеина . Кофакторы ионов металлов подвергаются окислительно-восстановительным реакциям без связывания или высвобождения протонов, поэтому в цепи переноса электронов они служат исключительно для транспортировки электронов через белки. Электроны перемещаются через белки на довольно большие расстояния, прыгая по цепочкам этих кофакторов. [14] Это происходит за счет квантового туннелирования , которое происходит быстро на расстояниях менее 1,4 × 10 −9 м. [15]
Эукариотические цепи переноса электронов [ править ]
Многие катаболические биохимические процессы, такие как гликолиз , цикл лимонной кислоты и бета-окисление , приводят к образованию восстановленного кофермента НАДН . Этот кофермент содержит электроны, имеющие высокий потенциал переноса ; другими словами, они высвободят большое количество энергии при окислении. Однако клетка не выделяет всю эту энергию сразу, так как это будет неконтролируемая реакция. Вместо этого электроны отделяются от НАДН и передаются кислороду через ряд ферментов, каждый из которых выделяет небольшое количество энергии. Этот набор ферментов, состоящий из комплексов I–IV, называется цепью переноса электронов и находится во внутренней мембране митохондрии . Сукцинат также окисляется в цепи переноса электронов, но участвует в этом пути в другой точке.
У эукариот ферменты этой системы переноса электронов используют энергию, выделяемую из O 2 НАДН, для перекачки протонов через внутреннюю мембрану митохондрии. Это приводит к скоплению протонов в межмембранном пространстве и созданию электрохимического градиента на мембране. Энергия, запасенная в этом потенциале, затем используется АТФ-синтазой для производства АТФ. Окислительное фосфорилирование в митохондриях эукариот является наиболее изученным примером этого процесса. Митохондрии присутствуют почти у всех эукариот, за исключением анаэробных простейших, таких как Trichomonas vaginalis , которые вместо этого восстанавливают протоны до водорода в остатке митохондрии, называемом гидрогеносомой . [16]
| Дыхательный фермент | Редокс-пара | Потенциал средней точки (Вольты) |
|---|---|---|
| НАДН-дегидрогеназа | ОНИ + / НАДН | −0.32 [17] |
| Сукцинатдегидрогеназа | FMN или FAD / FMNH 2 или FADH 2 | −0.20 [17] |
| Цитохром bc 1 комплекс | Коэнзим Q10 бычий / Коэнзим Q10 красный | +0.06 [17] |
| Цитохром bc 1 комплекс | Цитохром бокс / Цитохром b красный | +0.12 [17] |
| Комплекс IV | Цитохром c ox / Цитохром c красный | +0.22 [17] |
| Комплекс IV | Цитохром быка / Цитохром красный | +0.29 [17] |
| Комплекс IV | О 2 / НО − | +0.82 [17] |
| Условия: pH = 7 [17] | ||
НАДН-коэнзим Q-оксидоредуктаза ( I комплекс )
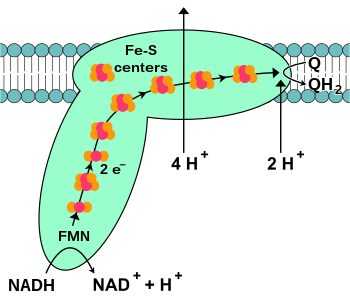
НАДН-коэнзим Q-оксидоредуктаза , также известная как НАДН-дегидрогеназа или комплекс I , является первым белком в цепи переноса электронов. [18] Комплекс I представляет собой гигантский фермент , комплекс I млекопитающих имеет 46 субъединиц и молекулярную массу около 1000 килодальтон (кДа). [19] Строение подробно известно только по бактериям; [20] [21] у большинства организмов комплекс напоминает ботинок с большим «шаром», торчащим из мембраны в митохондрии. [22] [23] Гены, кодирующие отдельные белки, содержатся как в ядре клетки , так и в митохондриальном геноме , как и в случае со многими ферментами, присутствующими в митохондриях.
Реакция, катализируемая этим ферментом, представляет собой двухэлектронное окисление НАДН коферментом Q10 или убихиноном (представленным как Q в уравнении ниже), жирорастворимым хиноном , который находится в мембране митохондрий:
| ( 1 ) |

Началом реакции, да и всей электронной цепи, является связывание молекулы НАДН с комплексом I и отдача двух электронов. Электроны входят в комплекс I через простетическую группу , присоединенную к комплексу, флавинмононуклеотид (FMN). Добавление электронов к FMN преобразует его в восстановленную форму FMNH 2 . Затем электроны передаются через серию железо-серных кластеров: второй вид простетической группы, присутствующей в комплексе. [20] В комплексе I присутствуют как железо-серные кластеры [2Fe–2S], так и [4Fe–4S].
При прохождении электронов через этот комплекс четыре протона перекачиваются из матрикса в межмембранное пространство. Как именно это происходит, неясно, но, по-видимому, это связано с конформационными изменениями в комплексе I, которые заставляют белок связывать протоны на N-стороне мембраны и высвобождать их на P-стороне мембраны. [24] Наконец, электроны передаются от цепочки железо-серных кластеров к молекуле убихинона в мембране. [18] Восстановление убихинона также способствует образованию протонного градиента, поскольку два протона захватываются из матрицы при восстановлении убихинола (QH 2 ).
-Q-оксидоредуктаза (комплекс Сукцинат ) II

Сукцинат-Q-оксидоредуктаза , также известная как комплекс II или сукцинатдегидрогеназа , является второй точкой входа в цепь переноса электронов. [25] Это необычно, поскольку это единственный фермент, который участвует как в цикле лимонной кислоты, так и в цепи переноса электронов. Комплекс II состоит из четырех белковых субъединиц и содержит связанный кофактор флавинадениндинуклеотид (FAD), железо-серные кластеры и гемовую группу, которая не участвует в переносе электрона к коферменту Q, но, как полагают, играет важную роль в снижении продукции реактивного виды кислорода. [26] [27] Он окисляет сукцинат до фумарата и восстанавливает убихинон. Поскольку в этой реакции выделяется меньше энергии, чем при окислении НАДН, комплекс II не переносит протоны через мембрану и не вносит вклада в протонный градиент.
| ( 2 ) |

У некоторых эукариот, таких как паразитический червь Ascaris suum , фермент, подобный комплексу II, фумаратредуктаза (менахинол:фумаратоксидоредуктаза, или QFR), действует наоборот, окисляя убихинол и восстанавливая фумарат. Это позволяет червю выживать в анаэробной среде толстого кишечника , осуществляя анаэробное окислительное фосфорилирование с фумаратом в качестве акцептора электронов. [28] Другая нетрадиционная функция комплекса II наблюдается у малярийного паразита Plasmodium falciparum . Здесь обратное действие комплекса II как оксидазы важно для регенерации убихинола, который паразит использует в необычной форме биосинтеза пиримидинов . [29]
Электронпереносящая флавопротеин оксидоредуктаза - Q
Флавопротеин-убихинон-оксидоредуктаза, переносящая электроны (ETF-Q-оксидоредуктаза), также известная как флавопротеин-переносящая электроны дегидрогеназа , является третьей точкой входа в цепь переноса электронов. Это фермент, который принимает электроны от переносящего электроны флавопротеина в митохондриальном матриксе и использует эти электроны для восстановления убихинона. [30] Этот фермент содержит флавин и кластер [4Fe–4S], но, в отличие от других дыхательных комплексов, прикрепляется к поверхности мембраны и не проникает через липидный бислой. [31]
| ( 3 ) |

У млекопитающих этот метаболический путь важен для бета-окисления и жирных кислот катаболизма аминокислот и холина , поскольку он принимает электроны от множества ацетил-КоА -дегидрогеназ. [32] [33] У растений оксидоредуктаза ETF-Q также играет важную роль в метаболических реакциях, которые позволяют выжить в длительные периоды темноты. [34]
(комплекс Q-цитохром с оксидоредуктаза ) III

Q-цитохром c-оксидоредуктаза также известна как цитохром c-редуктаза , цитохрома bc 1 комплекс или просто комплекс III . [35] [36] У млекопитающих этот фермент представляет собой димер , каждый комплекс субъединиц которого содержит 11 субъединиц белка, железо-серный кластер [2Fe-2S] и три цитохрома : один цитохром c 1 и два цитохрома b . [37] Цитохром — это разновидность белка-переносчика электронов, который содержит по крайней мере одну гемовую группу. Атомы железа внутри гемовых групп комплекса III чередуются между восстановленным трехвалентным (+2) и окисленным трехвалентным состоянием (+3), когда электроны передаются через белок.
Реакция, катализируемая комплексом III, представляет собой окисление одной молекулы убихинола и восстановление двух молекул цитохрома с — гемового белка, слабо связанного с митохондрией. В отличие от кофермента Q, который несет два электрона, цитохром С несет только один электрон.
| ( 4 ) |

Поскольку только один электрон может быть перенесен от донора QH 2 к акцептору цитохрома с за раз, механизм реакции комплекса III более сложен, чем у других дыхательных комплексов, и происходит в два этапа, называемых Q-циклом . [38] На первом этапе фермент связывает три субстрата, сначала QH 2 , который затем окисляется, при этом один электрон передается второму субстрату, цитохрому с. Два протона, высвободившиеся из QH 2, переходят в межмембранное пространство. Третий субстрат — Q, который принимает второй электрон от QH 2 и восстанавливается до Q .− , который является убисемихинона свободным радикалом . Первые два субстрата высвобождаются, но этот промежуточный убисемихинон остается связанным. На втором этапе вторая молекула QH 2 связывается и снова передает свой первый электрон акцептору цитохрома с. Второй электрон передается связанному убисемихинону, восстанавливая его до QH 2, поскольку он получает два протона из митохондриального матрикса. Этот QH 2 затем высвобождается из фермента. [39]
Поскольку коэнзим Q восстанавливается до убихинола на внутренней стороне мембраны и окисляется до убихинона на другой, происходит суммарный перенос протонов через мембрану, увеличивающий протонный градиент. [5] Довольно сложный двухэтапный механизм, с помощью которого это происходит, важен, поскольку повышает эффективность переноса протона. Если бы вместо цикла Q одна молекула QH 2 использовалась для прямого восстановления двух молекул цитохрома с, эффективность снизилась бы вдвое, при этом на каждый цитохром с был бы перенесен только один протон. [5]
Цитохром с оксидаза (комплекс IV) [ править ]

Цитохром С-оксидаза , также известная как комплекс IV , является конечным белковым комплексом в цепи переноса электронов. [40] Фермент млекопитающих имеет чрезвычайно сложную структуру и содержит 13 субъединиц, две гемовые группы, а также множество кофакторов ионов металлов – всего три атома меди , один магния и один цинка . [41]
Этот фермент опосредует конечную реакцию в цепи переноса электронов и переносит электроны на кислород и водород (протоны), одновременно перекачивая протоны через мембрану. [42] На этом этапе конечный акцептор электронов кислород восстанавливается до воды. В протонный градиент вносят вклад как прямая накачка протонов, так и расходование матричных протонов на восстановление кислорода. Катализируемой реакцией является окисление цитохрома с и восстановление кислорода:
| ( 5 ) |

и Альтернативные оксидазы редуктазы
Многие эукариотические организмы имеют цепи переноса электронов, которые отличаются от широко изученных ферментов млекопитающих, описанных выше. Например, у растений есть альтернативные НАДН-оксидазы, которые окисляют НАДН в цитозоле, а не в митохондриальном матриксе, и передают эти электроны в пул убихинонов. [43] Эти ферменты не переносят протоны и, следовательно, восстанавливают убихинон, не изменяя электрохимический градиент на внутренней мембране. [44]
Другим примером расходящейся цепи переноса электронов является альтернативная оксидаза , которая встречается у растений , а также у некоторых грибов , протистов и, возможно, у некоторых животных. [45] [46] Этот фермент переносит электроны непосредственно от убихинола к кислороду. [47]
Пути переноса электронов, продуцируемые этими альтернативными НАДН и убихиноноксидазами, имеют более низкие выходы АТФ , чем полный путь. Преимущества укороченного пути не совсем ясны. Однако альтернативная оксидаза вырабатывается в ответ на стрессы, такие как холод, активные формы кислорода и инфекция патогенами, а также другие факторы, которые ингибируют полную цепь транспорта электронов. [48] [49] Таким образом, альтернативные пути могут повысить устойчивость организма к повреждениям за счет снижения окислительного стресса . [50]
Организация комплексов [ править ]
Первоначальная модель организации комплексов дыхательной цепи заключалась в том, что они свободно и независимо диффундируют в митохондриальную мембрану. [51] Однако недавние данные позволяют предположить, что комплексы могут образовывать структуры более высокого порядка, называемые суперкомплексами или « респирасомами ». [52] В этой модели различные комплексы существуют как организованные наборы взаимодействующих ферментов. [53] Эти ассоциации могут позволить осуществлять каналирование субстратов между различными ферментными комплексами, увеличивая скорость и эффективность переноса электронов. [54] В таких суперкомплексах млекопитающих некоторые компоненты будут присутствовать в больших количествах, чем другие, при этом некоторые данные позволяют предположить, что соотношение между комплексами I/II/III/IV и АТФ-синтазой составляет примерно 1:1:3:7:4. [55] Однако дебаты по поводу этой сверхсложной гипотезы не решены полностью, поскольку некоторые данные, похоже, не соответствуют этой модели. [19] [56]
Прокариотические цепи переноса электронов [ править ]
В отличие от общего сходства строения и функций электрон-транспортных цепей у эукариот, бактерии и археи обладают большим разнообразием ферментов переноса электронов. В качестве субстратов они используют столь же широкий набор химических веществ. [57] Как и у эукариот, прокариотический транспорт электронов использует энергию, выделяемую при окислении субстрата, для перекачки ионов через мембрану и создания электрохимического градиента. У бактерий окислительное фосфорилирование у Escherichia coli , тогда как у архейных систем в настоящее время изучено мало. наиболее подробно изучено [58]
Основное различие между эукариотическим и прокариотическим окислительным фосфорилированием заключается в том, что бактерии и археи используют множество разных веществ для отдачи или принятия электронов. Это позволяет прокариотам расти в самых разных условиях окружающей среды. [59] Например, в E. coli окислительное фосфорилирование может управляться большим количеством пар восстановителей и окислителей, которые перечислены ниже. Потенциал средней точки химического вещества измеряет, сколько энергии высвобождается при его окислении или восстановлении, при этом восстановители имеют отрицательные потенциалы, а окислители - положительные потенциалы.
| Дыхательный фермент | Редокс-пара | Потенциал средней точки (Вольты) |
|---|---|---|
| Формиат дегидрогеназа | Бикарбонат / Формиат | −0.43 |
| Гидрогеназа | Протон / Водород | −0.42 |
| НАДН-дегидрогеназа | ОНИ + / НАДН | −0.32 |
| Глицерол-3-фосфатдегидрогеназа | ДГАП / Гли-3-П | −0.19 |
| Пируватоксидаза | Ацетат + углекислый газ / пируват | ? |
| Лактатдегидрогеназа | Пируват / лактат | −0.19 |
| Дегидрогеназа D -аминокислот | 2-оксокислота + аммиак / D -аминокислота | ? |
| Глюкозодегидрогеназа | Глюконат / Глюкоза | −0.14 |
| Сукцинатдегидрогеназа | Фумарат / Сукцинат | +0.03 |
| Убихинолоксидаза | Кислород / Вода | +0.82 |
| Нитратредуктаза | Нитрат / Нитрит | +0.42 |
| Нитритредуктаза | Нитриты / Аммиак | +0.36 |
| Диметилсульфоксидредуктаза | ДМСО / ДМС | +0.16 |
| Триметиламин- N -оксидредуктаза | ТМАО / ТМА | +0.13 |
| Фумаратредуктаза | Фумарат / Сукцинат | +0.03 |
Как показано выше, E. coli может расти с восстанавливающими агентами, такими как формиат, водород или лактат, в качестве доноров электронов, а также нитратом, ДМСО или кислородом в качестве акцепторов. [59] Чем больше разница потенциалов средней точки между окислителем и восстановителем, тем больше энергии выделяется при их реакции. Среди этих соединений необычной является пара сукцинат/фумарат, поскольку ее средний потенциал близок к нулю. Таким образом, сукцинат можно окислить до фумарата, если доступен сильный окислитель, такой как кислород, или фумарат можно восстановить до сукцината с использованием сильного восстановителя, такого как формиат. Эти альтернативные реакции катализируются сукцинатдегидрогеназой и фумаратредуктазой соответственно. [61]
Некоторые прокариоты используют окислительно-восстановительные пары, которые имеют лишь небольшую разницу в потенциале средней точки. Например, нитрифицирующие бактерии, такие как Nitrobacter, окисляют нитрит в нитрат, отдавая электроны кислороду. Небольшого количества энергии, выделяющейся в этой реакции, достаточно для перекачки протонов и генерации АТФ, но недостаточно для производства НАДН или НАДФН непосредственно для использования в анаболизме . [62] Эта проблема решается за счет использования нитритоксидоредуктазы для создания достаточной протондвижущей силы, чтобы запустить часть цепи переноса электронов в обратном направлении, заставляя комплекс I генерировать НАДН. [63] [64]
Прокариоты контролируют использование этих доноров и акцепторов электронов, изменяя, какие ферменты производятся в зависимости от условий окружающей среды. [65] Такая гибкость возможна, поскольку разные оксидазы и редуктазы используют один и тот же пул убихинонов. Это позволяет многим комбинациям ферментов функционировать вместе, связанных общим промежуточным продуктом убихинола. [60] Таким образом, эти дыхательные цепи имеют модульную конструкцию с легко заменяемыми наборами ферментных систем.
В дополнение к этому метаболическому разнообразию прокариоты также обладают рядом изоферментов – разных ферментов, катализирующих одну и ту же реакцию. Например, в E. coli существует два разных типа убихинолоксидазы, использующих кислород в качестве акцептора электронов. В высокоаэробных условиях клетка использует оксидазу с низким сродством к кислороду, которая может переносить два протона на электрон. Однако если уровень кислорода падает, они переключаются на оксидазу, которая переносит только один протон на электрон, но имеет высокое сродство к кислороду. [66]
АТФ-синтаза (комплекс V) [ править ]
АТФ-синтаза, также называемая комплексом V , является конечным ферментом пути окислительного фосфорилирования. Этот фермент встречается во всех формах жизни и функционирует одинаково как у прокариот, так и у эукариот. [67] Фермент использует энергию, запасенную в протонном градиенте через мембрану, для стимулирования синтеза АТФ из АДФ и фосфата (P i ). Оценки числа протонов, необходимых для синтеза одной АТФ, колеблются от трех до четырех. [68] [69] при этом некоторые предполагают, что клетки могут изменять это соотношение в зависимости от условий. [70]
| ( 6 ) |

Эта фосфорилирования реакция представляет собой равновесие , которое можно сдвинуть, изменяя протондвижущую силу. В отсутствие протондвижущей силы реакция АТФ-синтазы будет идти справа налево, гидролизуя АТФ и выкачивая протоны из матрикса через мембрану. Однако когда протондвижущая сила велика, реакция вынуждена идти в противоположном направлении; он идет слева направо, позволяя протонам течь по градиенту концентрации и превращать АДФ в АТФ. [67] Действительно, в близкородственных H+-АТФазах вакуолярного типа реакция гидролиза используется для подкисления клеточных компартментов путем перекачки протонов и гидролиза АТФ. [71]
АТФ-синтаза представляет собой массивный белковый комплекс грибовидной формы. Ферментный комплекс млекопитающих содержит 16 субъединиц и имеет массу около 600 килодальтон . [72] Часть, встроенная в мембрану, называется FO и содержит кольцо из субъединиц c и протонный канал. Стебель и шаровидная головка называются F 1 и являются местом синтеза АТФ. Шарообразный комплекс на конце участка F 1 содержит шесть белков двух разных видов (три субъединицы α и три субъединицы β), тогда как «стебель» состоит из одного белка: субъединицы γ с кончиком ножки. распространяющийся в клубок субъединиц α и β. [73] Обе субъединицы α и β связывают нуклеотиды, но только субъединицы β катализируют реакцию синтеза АТФ. Вдоль стороны части F 1 и обратно в мембрану проходит длинная стержнеобразная субъединица, которая закрепляет субъединицы α и β в основании фермента.
Когда протоны пересекают мембрану через канал в основании АТФ-синтазы, двигатель F O , управляемый протонами, вращается. [74] Вращение может быть вызвано изменениями в ионизации аминокислот в кольце субъединиц c, вызывающими электростатические взаимодействия, которые продвигают кольцо субъединиц c мимо протонного канала. [75] Это вращающееся кольцо, в свою очередь, приводит в движение центральную ось (стебель субъединицы γ) внутри субъединиц α и β. Субъединицы α и β не могут вращаться боковым рычагом, который действует как статор . Это движение кончика субъединицы γ внутри шара субъединиц α и β обеспечивает энергию для активных центров субъединиц β для прохождения цикла движений, который производит, а затем высвобождает АТФ. [76]

Эта реакция синтеза АТФ называется механизмом изменения связывания и включает в себя активный центр β-субъединицы, циклически переключающийся между тремя состояниями. [77] В «открытом» состоянии АДФ и фосфат попадают в активный центр (на схеме показан коричневым цветом). Затем белок замыкается вокруг молекул и свободно связывает их – «рыхлое» состояние (показано красным). Затем фермент снова меняет форму и сближает эти молекулы, в результате чего активный центр оказывается в «плотном» состоянии (показан розовым), связывая вновь полученную молекулу АТФ с очень высоким сродством . Наконец, активный сайт возвращается в открытое состояние, высвобождая АТФ и связывая больше АДФ и фосфата, готовясь к следующему циклу.
У некоторых бактерий и архей синтез АТФ обусловлен движением ионов натрия через клеточную мембрану, а не движением протонов. [78] [79] Археи, такие как Methanococcus, также содержат A 1 A o -синтазу, форму фермента, которая содержит дополнительные белки с небольшим сходством по последовательности с другими субъединицами бактериальной и эукариотической АТФ-синтазы. Возможно, что у некоторых видов форма A 1 A o фермента представляет собой специализированную АТФ-синтазу, управляемую натрием, [80] но это может быть не так во всех случаях. [79]
Окислительное фосфорилирование - энергетика [ править ]
Транспорт электронов от окислительно-восстановительной пары НАД + / НАДН к конечной окислительно-восстановительной паре 1/2 O 2 / H 2 O можно суммировать как
1/2 О 2 + НАДН + Н + → H 2 O + NAD +
Разность потенциалов между этими двумя окислительно-восстановительными парами составляет 1,14 вольт, что эквивалентно -52 ккал/моль или -2600 кДж на 6 моль O 2 .
При окислении одного НАДН по цепи переноса электронов образуются три АТФ, что эквивалентно 7,3 ккал/моль х 3 = 21,9 ккал/моль.
Сохранение энергии можно рассчитать по следующей формуле
Эффективность = (21,9 х 100%) / 52 = 42%
Таким образом, можно заключить, что при окислении НАДН около 42% энергии сохраняется в виде трех АТФ, а оставшаяся (58%) энергия теряется в виде тепла (если только химическая энергия АТФ в физиологических условиях не была недооценена).
кислорода Активные формы
Молекулярный кислород является хорошим акцептором концевых электронов , поскольку является сильным окислителем. Восстановление кислорода действительно связано с потенциально вредными промежуточными продуктами. [81] Хотя перенос четырех электронов и четырех протонов превращает кислород в воду, что безвредно, перенос одного или двух электронов приводит к образованию супероксидных или пероксидных анионов, которые являются опасно реакционноспособными.
| ( 7 ) |
![{\displaystyle {\ce {O2->[{\ce {e^{-}}}]{\underset {Супероксид}}{O2^{\underline {\bullet }}}}->[{\ce {e ^{-}}}]{\underset {Перекись}{O2^{2-}}}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/a3d9bf9d3a61736aa6207fa53b8ce0165b9eebb6)
Эти активные формы кислорода и продукты их реакции, такие как гидроксильный радикал, очень вредны для клеток, поскольку они окисляют белки и вызывают мутации в ДНК . Это клеточное повреждение может способствовать развитию заболеваний и считается одной из причин старения . [82] [83]
Комплекс цитохром-с-оксидазы очень эффективно восстанавливает кислород до воды и выделяет очень мало частично восстановленных промежуточных продуктов; однако небольшие количества супероксид-аниона и пероксида производятся цепью переноса электронов. [84] Особенно важным является восстановление кофермента Q в комплексе III, поскольку в качестве интермедиата в цикле Q образуется высокореакционноспособный свободный радикал убисемихинон. Этот нестабильный вид может привести к «утечке» электронов, когда электроны переходят непосредственно к кислороду, образуя супероксид. [85] Поскольку производство активных форм кислорода этими комплексами, перекачивающими протоны, является наибольшим при высоких мембранных потенциалах, было высказано предположение, что митохондрии регулируют свою активность для поддержания мембранного потенциала в узком диапазоне, который уравновешивает производство АТФ и образование оксидантов. [86] Например, оксиданты могут активировать разобщающие белки , которые снижают мембранный потенциал. [87]
Чтобы противодействовать этим активным формам кислорода, клетки содержат многочисленные антиоксидантные системы, в том числе витамины- антиоксиданты, такие как витамин С и витамин Е , а также антиоксидантные ферменты, такие как супероксиддисмутаза , каталаза и пероксидазы . [81] которые детоксицируют реактивные виды, ограничивая повреждение клетки.
Окислительное фосфорилирование в условиях гипоксии [ править ]
Поскольку кислород имеет основополагающее значение для окислительного фосфорилирования, дефицит уровня O2 , вероятно, изменяет скорость производства АТФ. Однако движущая сила протонов и выработка АТФ могут поддерживаться за счет внутриклеточного ацидоза. [88] Цитозольные протоны, накопившиеся в результате гидролиза АТФ и лактоацидоза, могут свободно диффундировать через внешнюю мембрану митохондрий и закислять межмембранное пространство, тем самым непосредственно внося вклад в движущую силу протонов и выработку АТФ.
Ингибиторы [ править ]
Существует несколько хорошо известных препаратов и токсинов , ингибирующих окислительное фосфорилирование. Хотя любой из этих токсинов ингибирует только один фермент в цепи переноса электронов, ингибирование любого этапа этого процесса остановит остальную часть процесса. Например, если олигомицин ингибирует АТФ-синтазу, протоны не могут вернуться обратно в митохондрии. [89] В результате протонные насосы не могут работать, поскольку градиент становится слишком сильным, чтобы они могли его преодолеть. НАДН больше не окисляется, и цикл лимонной кислоты прекращается, поскольку концентрация НАДН + падает ниже концентрации, которую могут использовать эти ферменты.
Многие сайт-специфические ингибиторы цепи переноса электронов внесли свой вклад в современные знания о митохондриальном дыхании. Синтез АТФ также зависит от цепи переноса электронов, поэтому все сайт-специфические ингибиторы также ингибируют образование АТФ. Рыбий яд ротенон , барбитурат амитал и антибиотик пиерицидин А ингибируют НАДН и коэнзим Q. [90]
Окись углерода, цианид, сероводород и азид эффективно ингибируют цитохромоксидазу. Окись углерода реагирует с восстановленной формой цитохрома, тогда как цианид и азид реагируют с окисленной формой. Антибиотик антимицин А и британский антилюизит , противоядие, используемое против химического оружия, являются двумя важными ингибиторами участка между цитохромом B и C1. [90]
| Соединения | Использовать | Место действия | Влияние на окислительное фосфорилирование |
|---|---|---|---|
| Цианид Окись углерода Азид Сероводород | Яды | Комплекс IV | Ингибируют цепь переноса электронов, связываясь сильнее, чем кислород, с центром Fe - Cu в цитохром-с-оксидазе, предотвращая восстановление кислорода. [91] |
| Олигомицин | Антибиотик | Комплекс V | Ингибирует АТФ-синтазу, блокируя поток протонов через субъединицу F o . [89] |
| CCCP 2,4-динитрофенол | Яды, потеря веса [Н 1] | Внутренняя мембрана | Ионофоры , которые нарушают протонный градиент, перенося протоны через мембрану. Этот ионофор отключает перекачку протонов от синтеза АТФ, поскольку он переносит протоны через внутреннюю митохондриальную мембрану. [92] |
| Ротенон | Пестицид | Комплекс I | Предотвращает перенос электронов от комплекса I к убихинону, блокируя сайт связывания убихинона. [93] |
| Малонат и оксалоацетат | Яды | Комплекс II | Конкурентные ингибиторы сукцинатдегидрогеназы (комплекс II). [94] |
| Антимицин А | Писцидицид | Комплекс III | Связывается с сайтом Qi редуктазы цитохрома с тем самым ингибируя окисление убихинола , . |
Не все ингибиторы окислительного фосфорилирования являются токсинами. В бурой жировой ткани регулируемые протонные каналы, называемые разобщающими белками, могут отсоединить дыхание от синтеза АТФ. [95] Это быстрое дыхание производит тепло и особенно важно как способ поддержания температуры тела у животных, находящихся в спячке , хотя эти белки могут также выполнять более общую функцию в ответах клеток на стресс. [96]
История [ править ]
Область окислительного фосфорилирования началась с сообщения Артура Хардена в 1906 году о жизненно важной роли фосфатов в клеточном брожении , но первоначально только фосфаты сахаров . было известно, что в них участвуют [97] Однако в начале 1940-х годов связь между окислением сахаров и образованием АТФ была твердо установлена Германом Калькаром , [98] подтверждая центральную роль АТФ в передаче энергии, предложенную Фрицем Альбертом Липманом в 1941 году. [99] Позже, в 1949 году, Моррис Фридкин и Альберт Л. Ленинджер доказали, что кофермент НАДН связан с такими метаболическими путями, как цикл лимонной кислоты и синтез АТФ. [100] Термин окислительное фосфорилирование был введен Владимиром Белицером в 1939 году. [101] [102]
Еще двадцать лет механизм образования АТФ оставался загадочным, и ученые искали неуловимый «высокоэнергетический промежуточный продукт», который мог бы связать реакции окисления и фосфорилирования. [103] Эту загадку решил Питер Д. Митчелл, опубликовав в 1961 году хемиосмотическую теорию . [104] Поначалу это предложение вызвало большие споры, но постепенно оно было принято, и в 1978 году Митчеллу была присуждена Нобелевская премия . [105] [106] Последующие исследования были сосредоточены на очистке и характеристике задействованных ферментов, при этом большой вклад внесли Дэвид Э. Грин в комплексы электрон-транспортной цепи, а также Эфраим Рэкер в АТФ-синтазу. [107] Критический шаг на пути к раскрытию механизма АТФ-синтазы был сделан Полом Д. Бойером , когда он разработал в 1973 году механизм «изменения связывания», за которым последовало его радикальное предложение ротационного катализа в 1982 году. [77] [108] Более поздние работы включали структурные исследования ферментов, участвующих в окислительном фосфорилировании, проведенные Джоном Э. Уокером , при этом Уокер и Бойер были удостоены Нобелевской премии в 1997 году. [109]
См. также [ править ]
Примечания [ править ]
- ^ DNP широко использовался в качестве лекарства от ожирения в 1930-х годах, но в конечном итоге был прекращен из-за его опасных побочных эффектов. Однако незаконное использование препарата с этой целью продолжается и сегодня. см. в разделе «2,4-динитрофенол # Диетическая помощь» . Дополнительную информацию
Ссылки [ править ]
- ^ «Значение окислительного в Кембриджском словаре английского языка» . словарь.cambridge.org . Архивировано из оригинала 24 января 2018 года . Проверено 28 апреля 2018 г.
- ^ Воэт, Д.; Воэт, JG (2004). «Биохимия», 3-е изд., с. 804, Wiley.ISBN 0-471-19350-X.
- ^ Митчелл П., Мойл Дж. (1967). «Хемиосмотическая гипотеза окислительного фосфорилирования». Природа . 213 (5072): 137–9. Бибкод : 1967Natur.213..137M . дои : 10.1038/213137a0 . ПМИД 4291593 . S2CID 4149605 .
- ↑ Перейти обратно: Перейти обратно: а б Димрот П., Каим Г., Мэтью У (1 января 2000 г.). «Решающая роль мембранного потенциала в синтезе АТФ с помощью F (1) F (о) АТФ-синтаз» . Дж. Эксп. Биол . 203 (Часть 1): 51–9. дои : 10.1242/jeb.203.1.51 . ПМИД 10600673 . Архивировано из оригинала 30 сентября 2007 года.
- ↑ Перейти обратно: Перейти обратно: а б с д Шульц Б.Е., Чан С.И. (2001). «Структуры и стратегии перекачки протонов митохондриальных дыхательных ферментов» (PDF) . Annu Rev Biophys Biomol Struct . 30 :23–65. doi : 10.1146/annurev.biophys.30.1.23 . ПМИД 11340051 .
- ^ Богатый пиар (2003). «Молекулярный механизм дыхательной цепи Кейлина». Биохим. Соц. Транс . 31 (Часть 6): 1095–105. дои : 10.1042/bst0311095 . ПМИД 14641005 .
- ^ Портер Р.К., Бренд, доктор медицинских наук (1995). «Митохондриальная протонная проводимость и соотношение H+/O не зависят от скорости транспорта электронов в изолированных гепатоцитах» . Биохим. Дж . 310 (Часть 2): 379–82. дои : 10.1042/bj3100379 . ПМЦ 1135905 . ПМИД 7654171 .
- ^ Мэтьюз Ф.С. (1985). «Строение, функции и эволюция цитохромов» . Прог. Биофиз. Мол. Биол . 45 (1): 1–56. дои : 10.1016/0079-6107(85)90004-5 . ПМИД 3881803 .
- ^ Вуд ПМ (1983). «Почему существуют цитохромы c-типа?» . ФЭБС Летт . 164 (2): 223–6. дои : 10.1016/0014-5793(83)80289-0 . ПМИД 6317447 . S2CID 7685958 .
- ^ Крейн, Флорида (1 декабря 2001 г.). «Биохимические функции коэнзима Q10». J Am Coll Nutr . 20 (6): 591–8. дои : 10.1080/07315724.2001.10719063 . ПМИД 11771674 . S2CID 28013583 .
- ^ Митчелл П. (1979). «Концепция дыхательной цепи Кейлина и ее хемиосмотические последствия». Наука . 206 (4423): 1148–59. Бибкод : 1979Sci...206.1148M . дои : 10.1126/science.388618 . ПМИД 388618 .
- ^ Сёбалле Б., Пул Р.К. (1999). «Микробные убихиноны: многочисленные роли в дыхании, регуляции генов и управлении окислительным стрессом» (PDF) . Микробиология . 145 (8): 1817–30. дои : 10.1099/13500872-145-8-1817 . ПМИД 10463148 . Архивировано (PDF) из оригинала 29 мая 2008 г.
- ^ Джонсон, округ Колумбия, Дин Д.Р., Смит А.Д., Джонсон М.К. (2005). «Структура, функции и образование биологических железо-серных кластеров». Анну. Преподобный Биохим . 74 : 247–81. doi : 10.1146/annurev.biochem.74.082803.133518 . ПМИД 15952888 .
- ^ Пейдж CC, Мозер CC, Чен X, Даттон PL (1999). «Природно-инженерные принципы туннелирования электронов при биологическом окислении-восстановлении». Природа . 402 (6757): 47–52. Бибкод : 1999Natur.402...47P . дои : 10.1038/46972 . ПМИД 10573417 . S2CID 4431405 .
- ^ Лейс Д., Скраттон Н.С. (2004). «Электрические схемы в биологии: новые принципы структуры белка». Курс. Мнение. Структура. Биол . 14 (6): 642–7. дои : 10.1016/j.sbi.2004.10.002 . ПМИД 15582386 .
- ^ Боксма Б, де Грааф Р.М., ван дер Стаай Г.В., ван Ален Т.А., Рикард Дж., Габальдон Т., ван Хук А.Х., Мун-ван дер Стаай С.Ю., Купман В.Дж., ван Хеллемонд Дж.Дж., Тиленс А.Г., Фридрих Т., Винхейс М., Хюйнен Массачусетс, Хакштейн Дж. Х. (2005). «Анаэробная митохондрия, производящая водород» (PDF) . Природа . 434 (7029): 74–9. Стартовый код : 2005Natur.434...74B . дои : 10.1038/nature03343 . ПМИД 15744302 . S2CID 4401178 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час Справочник по медицинской химии. Андерс Овергаард Педерсен и Хеннинг Нильсен. Орхусский университет. 2008 год
- ↑ Перейти обратно: Перейти обратно: а б Херст Дж (2005). «Передача энергии дыхательным комплексом I - оценка современных знаний». Биохим. Соц. Транс . 33 (Часть 3): 525–9. дои : 10.1042/BST0330525 . ПМИД 15916556 .
- ↑ Перейти обратно: Перейти обратно: а б Леназ Г., Фато Р., Дженова М.Л., Бергамини С., Бьянки С., Бионди А. (2006). «Митохондриальный комплекс I: структурные и функциональные аспекты» . Биохим. Биофиз. Акта . 1757 (9–10): 1406–20. дои : 10.1016/j.bbabio.2006.05.007 . ПМИД 16828051 .
- ↑ Перейти обратно: Перейти обратно: а б Сазанов Л.А., Хинчлифф П. (2006). «Структура гидрофильного домена дыхательного комплекса I Thermus thermophilus» . Наука . 311 (5766): 1430–6. Бибкод : 2006Sci...311.1430S . дои : 10.1126/science.1123809 . ПМИД 16469879 . S2CID 1892332 .
- ^ Ефремов Р.Г., Барадаран Р., Сазанов Л.А. (май 2010 г.). «Архитектура дыхательного комплекса I». Природа . 465 (7297): 441–5. Бибкод : 2010Natur.465..441E . дои : 10.1038/nature09066 . ПМИД 20505720 . S2CID 4372778 .
- ^ Баранова Е.А., Холт П.Дж., Сазанов Л.А. (2007). «Структура проекции мембранного домена дыхательного комплекса I Escherichia coli при разрешении 8 А». Дж. Мол. Биол . 366 (1): 140–54. дои : 10.1016/j.jmb.2006.11.026 . ПМИД 17157874 .
- ^ Фридрих Т., Бетчер Б. (2004). «Грубая структура дыхательного комплекса I: система Lego» . Биохим. Биофиз. Акта . 1608 (1): 1–9. дои : 10.1016/j.bbabio.2003.10.002 . ПМИД 14741580 .
- ^ Херст Дж. (январь 2010 г.). «К молекулярному механизму дыхательного комплекса I». Биохим. Дж . 425 (2): 327–39. дои : 10.1042/BJ20091382 . ПМИД 20025615 .
- ^ Чеккини Дж. (2003). «Функция и строение II комплекса дыхательной цепи». Анну. Преподобный Биохим . 72 : 77–109. doi : 10.1146/annurev.biochem.72.121801.161700 . ПМИД 14527321 .
- ^ Янковская В., Хорсфилд Р., Торнрот С., Луна-Чавес С., Миёши Х., Леже С., Бирн Б., Чеккини Г., Ивата С. и др. (2003). «Архитектура сукцинатдегидрогеназы и генерации активных форм кислорода». Наука . 299 (5607): 700–4. Бибкод : 2003Sci...299..700Y . дои : 10.1126/science.1079605 . ПМИД 12560550 . S2CID 29222766 .
- ^ Хорсфилд Р., Ивата С., Бирн Б. (2004). «Комплекс II со структурной точки зрения». Курс. Белковый пепт. Наука . 5 (2): 107–18. дои : 10.2174/1389203043486847 . ПМИД 15078221 .
- ^ Кита К., Хираваке Х., Миядера Х., Амино Х., Такео С. (2002). «Роль комплекса II в анаэробном дыхании митохондрий паразита Ascaris suum и Plasmodium falciparum» . Биохим. Биофиз. Акта . 1553 (1–2): 123–39. дои : 10.1016/S0005-2728(01)00237-7 . ПМИД 11803022 .
- ^ Художник Х.Дж., Морриси Дж.М., Мазер М.В., Вайдья А.Б. (2007). «Особая роль митохондриального транспорта электронов на стадии крови Plasmodium falciparum». Природа . 446 (7131): 88–91. Бибкод : 2007Natur.446...88P . дои : 10.1038/nature05572 . ПМИД 17330044 . S2CID 4421676 .
- ^ Рамзи Р.Р., Стинкамп диджей, Хусейн М. (1987). «Реакции электронпереносящего флавопротеина и электронпереносящего флавопротеина: убихинон оксидоредуктаза» . Биохим. Дж . 241 (3): 883–92. дои : 10.1042/bj2410883 . ПМЦ 1147643 . ПМИД 3593226 .
- ^ Чжан Дж., Фрерман Ф.Е., Ким Дж.Дж. (2006). «Структура электронного переноса флавопротеин-убихинон-оксидоредуктазы и перенос электрона в митохондриальный пул убихинона» . Учеб. Натл. акад. наук. США . 103 (44): 16212–7. Бибкод : 2006PNAS..10316212Z . дои : 10.1073/pnas.0604567103 . ПМЦ 1637562 . ПМИД 17050691 .
- ^ Икеда Ю., Дабровски С., Танака К. (25 января 1983 г.). «Выделение и свойства пяти различных ацил-КоА-дегидрогеназ из митохондрий печени крысы. Идентификация новой 2-метил-ацил-КоА-дегидрогеназы с разветвленной цепью» . Ж. Биол. Хим . 258 (2): 1066–76. дои : 10.1016/S0021-9258(18)33160-0 . ПМИД 6401712 . Архивировано из оригинала 29 сентября 2007 года.
- ^ Ружичка Ф.Дж., Бейнерт Х. (1977). «Новый железо-серный флавопротеин дыхательной цепи. Компонент пути бета-окисления жирных кислот» (PDF) . Ж. Биол. Хим . 252 (23): 8440–5. дои : 10.1016/S0021-9258(19)75238-7 . ПМИД 925004 . Архивировано (PDF) из оригинала 27 сентября 2007 г.
- ^ Ишизаки К., Ларсон Т.Р., Шауэр Н., Ферни А.Р., Грэм И.А., Ливер С.Дж. (2005). «Критическая роль флавопротеина переноса электронов арабидопсиса: убихиноноксидоредуктазы во время голодания, вызванного темнотой» . Растительная клетка . 17 (9): 2587–600. дои : 10.1105/tpc.105.035162 . ПМЦ 1197437 . ПМИД 16055629 .
- ^ Берри Э.А., Гергова-Курас М., Хуанг Л.С., Крофтс А.Р. (2000). «Структура и функция комплексов цитохрома bc» (PDF) . Анну. Преподобный Биохим . 69 : 1005–75. CiteSeerX 10.1.1.319.5709 . doi : 10.1146/annurev.biochem.69.1.1005 . ПМИД 10966481 . Архивировано (PDF) из оригинала 28 декабря 2015 г.
- ^ Крофтс А.Р. (2004). «Комплекс цитохрома bc1: функция в контексте структуры». Анну. Преподобный физиол . 66 : 689–733. doi : 10.1146/annurev.phyol.66.032102.150251 . ПМИД 14977419 .
- ^ Ивата С., Ли Дж.В., Окада К., Ли Дж.К., Ивата М., Расмуссен Б., Линк Т.А., Рамасвами С., Япончик Б.К. (1998). «Полная структура 11-субъединичного комплекса бычьего митохондриального цитохрома bc1». Наука . 281 (5373): 64–71. Бибкод : 1998Sci...281...64I . дои : 10.1126/science.281.5373.64 . ПМИД 9651245 .
- ^ Trumppower BL (1990). «Протонмотивный цикл Q. Трансдукция энергии путем сочетания транслокации протона с переносом электрона комплексом цитохрома bc1» (PDF) . Ж. Биол. Хим . 265 (20): 11409–12. дои : 10.1016/S0021-9258(19)38410-8 . ПМИД 2164001 . Архивировано (PDF) из оригинала 27 сентября 2007 г.
- ^ Хант С., Палсдоттир Х., Трампауэр Б.Л. (2003). «Протонмотивные пути и механизмы в комплексе цитохрома bc1» . ФЭБС Летт . 545 (1): 39–46. дои : 10.1016/S0014-5793(03)00391-0 . ПМИД 12788490 . S2CID 13942619 .
- ^ Кэлхун М.В., Томас Дж.В., Геннис Р.Б. (1994). «Суперсемейство цитохромоксидаз протонных насосов, управляемых окислительно-восстановительным процессом». Тенденции биохимии. Наука . 19 (8): 325–30. дои : 10.1016/0968-0004(94)90071-X . ПМИД 7940677 .
- ^ Цукихара Т., Аояма Х., Ямасита Э., Томидзаки Т., Ямагути Х., Синдзава-Ито К., Накашима Р., Яоно Р., Ёсикава С. (1996). «Вся структура 13-субъединичной окисленной цитохром с оксидазы при 2,8 А». Наука . 272 (5265): 1136–44. Бибкод : 1996Sci...272.1136T . дои : 10.1126/science.272.5265.1136 . ПМИД 8638158 . S2CID 20860573 .
- ^ Ёсикава С., Мурамото К., Синдзава-Ито К., Аояма Х., Цукихара Т., Симоката К., Катаяма Ю., Симада Х. (2006). «Механизм протонной накачки цитохром-с-оксидазы бычьего сердца» . Биохим. Биофиз. Акта . 1757 (9–10): 1110–6. дои : 10.1016/j.bbabio.2006.06.004 . ПМИД 16904626 .
- ^ Расмуссон А.Г., Сул К.Л., Элтон Т.Е. (2004). «Альтернативные НАД(Ф)Н-дегидрогеназы митохондрий растений». Annu Rev Plant Biol . 55 : 23–39. doi : 10.1146/annurev.arplant.55.031903.141720 . ПМИД 15725055 .
- ^ Менц Р.И., День ДА (1996). «Очистка и характеристика НАДН-дегидрогеназы 43 кДа, нечувствительной к ротенону, из митохондрий растений» . Ж. Биол. Хим . 271 (38): 23117–20. дои : 10.1074/jbc.271.38.23117 . ПМИД 8798503 . S2CID 893754 .
- ^ Макдональд А., Ванлерберг Г. (2004). «Разветвленный митохондриальный транспорт электронов у Animalia: наличие альтернативной оксидазы у нескольких типов животных» . ИУБМБ Жизнь . 56 (6): 333–41. дои : 10.1080/1521-6540400000876 . ПМИД 15370881 .
- ^ Слусе Ф.Е., Ярмушкевич В. (1998). «Альтернативная оксидаза в разветвленной дыхательной сети митохондрий: обзор структуры, функций, регуляции и роли» . Браз. Дж. Мед. Биол. Рез . 31 (6): 733–47. дои : 10.1590/S0100-879X1998000600003 . ПМИД 9698817 .
- ^ Мур А.Л., Сидов Дж.Н. (1991). «Регуляция и природа цианид-резистентной альтернативной оксидазы митохондрий растений». Биохим. Биофиз. Акта . 1059 (2): 121–40. дои : 10.1016/S0005-2728(05)80197-5 . ПМИД 1883834 .
- ^ Ванлерберге Г.К., Макинтош Л. (1997). «АЛЬТЕРНАТИВНАЯ ОКСИДАЗА: от гена к функции». Анну. Преподобный Физиол растений. Завод Мол. Биол . 48 : 703–734. doi : 10.1146/annurev.arplant.48.1.703 . ПМИД 15012279 .
- ^ Ито Ю, Сайшо Д, Накадзоно М, Цуцуми Н, Хираи А (1997). «Уровни транскриптов тандемно расположенных альтернативных генов оксидазы в рисе повышаются при низкой температуре». Джин . 203 (2): 121–9. дои : 10.1016/S0378-1119(97)00502-7 . ПМИД 9426242 .
- ^ Максвелл Д.П., Ван Ю., Макинтош Л. (1999). «Альтернативная оксидаза снижает выработку активного кислорода митохондриями в растительных клетках» . Учеб. Натл. акад. наук. США . 96 (14): 8271–6. Бибкод : 1999PNAS...96.8271M . дои : 10.1073/pnas.96.14.8271 . ПМК 22224 . ПМИД 10393984 .
- ^ Леназ Г (2001). «Критическая оценка пула митохондриального кофермента Q» . ФЭБС Летт . 509 (2): 151–5. дои : 10.1016/S0014-5793(01)03172-6 . ПМИД 11741580 . S2CID 46138989 .
- ^ Хайнемейер Дж., Браун Х.П., Букема Э.Дж., Курил Р. (2007). «Структурная модель суперкомплекса цитохром С редуктаза/оксидаза из митохондрий дрожжей» . Ж. Биол. Хим . 282 (16): 12240–8. дои : 10.1074/jbc.M610545200 . ПМИД 17322303 . S2CID 18123642 .
- ^ Шеггер Х., Пфайффер К. (2000). «Суперкомплексы в дыхательных цепях митохондрий дрожжей и млекопитающих» . ЭМБО Дж . 19 (8): 1777–83. дои : 10.1093/emboj/19.8.1777 . ПМК 302020 . ПМИД 10775262 .
- ^ Шеггер Х (2002). «Суперкомплексы дыхательной цепи митохондрий и бактерий» . Биохим. Биофиз. Акта . 1555 (1–3): 154–9. дои : 10.1016/S0005-2728(02)00271-2 . ПМИД 12206908 .
- ^ Шеггер Х., Пфайффер К. (2001). «Соотношение комплексов окислительного фосфорилирования IV в митохондриях сердца быка и состав суперкомплексов дыхательной цепи» . Ж. Биол. Хим . 276 (41): 37861–7. дои : 10.1074/jbc.M106474200 . ПМИД 11483615 . Архивировано из оригинала 29 сентября 2007 г.
- ^ Гупте С., Ву Э.С., Хоехли Л., Хоехли М., Джейкобсон К., Соуерс А.Э., Хакенброк Ч.Р. (1984). «Связь между латеральной диффузией, частотой столкновений и переносом электронов окислительно-восстановительных компонентов внутренней мембраны митохондрий» . Учеб. Натл. акад. наук. США . 81 (9): 2606–10. Бибкод : 1984PNAS...81.2606G . дои : 10.1073/pnas.81.9.2606 . ПМК 345118 . ПМИД 6326133 .
- ^ Нилсон К.Х. (1999). «Микробиология после викингов: новые подходы, новые данные, новые идеи». Ориг Лайф Эвол Биосф . 29 (1): 73–93. Бибкод : 1999ОЛЕВ...29...73Н . дои : 10.1023/А:1006515817767 . ПМИД 11536899 . S2CID 12289639 .
- ^ Шефер Г., Энгельхард М., Мюллер В. (1999). «Биоэнергетика архей» . Микробиол. Мол. Биол. Преподобный . 63 (3): 570–620. дои : 10.1128/MMBR.63.3.570-620.1999 . ПМЦ 103747 . ПМИД 10477309 .
- ↑ Перейти обратно: Перейти обратно: а б Инглдью В.Дж., Пул Р.К. (1984). «Дыхательные цепи кишечной палочки » . Микробиол. Преподобный . 48 (3): 222–71. дои : 10.1128/ммбр.48.3.222-271.1984 . ПМК 373010 . ПМИД 6387427 .
- ↑ Перейти обратно: Перейти обратно: а б Унден Дж., Бонгертс Дж. (1997). «Альтернативные дыхательные пути Escherichia coli : энергетика и регуляция транскрипции в ответ на акцепторы электронов» . Биохим. Биофиз. Акта . 1320 (3): 217–34. дои : 10.1016/S0005-2728(97)00034-0 . ПМИД 9230919 .
- ^ Чеккини Г., Шредер И., Гунсалус Р.П., Маклашина Е (2002). «Сукцинатдегидрогеназа и фумаратредуктаза Escherichia coli » . Биохим. Биофиз. Акта . 1553 (1–2): 140–57. дои : 10.1016/S0005-2728(01)00238-9 . ПМИД 11803023 .
- ^ Фрайтаг А, Бок Э (1990). «Энергосбережение в Nitrobacter» . Письма FEMS по микробиологии . 66 (1–3): 157–62. дои : 10.1111/j.1574-6968.1990.tb03989.x .
- ^ Старкенбург С.Р., Чейн П.С., Саяведра-Сото Л.А., Хаузер Л., Лэнд М.Л., Лаример Ф.В. и др. (март 2006 г.). «Последовательность генома хемолитоавтотрофной нитрит-окисляющей бактерии Nitrobacter winogradskyi Nb-255» . Прикладная и экологическая микробиология . 72 (3): 2050–63. Бибкод : 2006ApEnM..72.2050S . дои : 10.1128/АЕМ.72.3.2050-2063.2006 . ПМЦ 1393235 . ПМИД 16517654 .
- ^ Яманака Т., Фукумори Ю. (декабрь 1988 г.). «Нитритокисляющая система Nitrobacter winogradskyi» . Обзоры микробиологии FEMS . 54 (4): 259–70. дои : 10.1111/j.1574-6968.1988.tb02746.x . ПМИД 2856189 .
- ^ Ючи С., Лин ЕС (1993). «Адаптация Escherichia coli к окислительно-восстановительной среде путем экспрессии генов». Мол. Микробиол . 9 (1): 9–15. дои : 10.1111/j.1365-2958.1993.tb01664.x . ПМИД 8412675 . S2CID 39165641 .
- ^ Кэлхун М.В., Оден К.Л., Геннис Р.Б., де Маттос М.Дж., Нейссел О.М. (1993). «Энергетическая эффективность Escherichia coli : эффекты мутаций в компонентах аэробной дыхательной цепи» (PDF) . Дж. Бактериол . 175 (10): 3020–5. дои : 10.1128/jb.175.10.3020-3025.1993 . ПМК 204621 . ПМИД 8491720 . Архивировано (PDF) из оригинала 27 сентября 2007 г.
- ↑ Перейти обратно: Перейти обратно: а б Бойер П.Д. (1997). «АТФ-синтаза — великолепная молекулярная машина». Анну. Преподобный Биохим . 66 : 717–49. doi : 10.1146/annurev.biochem.66.1.717 . ПМИД 9242922 .
- ^ Ван Вальравен Х.С., Стротманн Х., Шварц О., Румберг Б. (1996). «Коэффициент связывания H + / АТФ АТФ-синтазы из тиол-модулированных хлоропластов и двух штаммов цианобактерий равен четырем». ФЭБС Летт . 379 (3): 309–13. дои : 10.1016/0014-5793(95)01536-1 . ПМИД 8603713 . S2CID 35989618 .
- ^ Ёсида М., Мунеюки Э., Хисабори Т. (2001). «АТФ-синтаза — чудесный роторный двигатель клетки». Нат. Преподобный мол. Клеточная Биол . 2 (9): 669–77. дои : 10.1038/35089509 . ПМИД 11533724 . S2CID 3926411 .
- ^ Шемидт Р.А., Цюй Дж., Уильямс-младший, Брусилов В.С. (1998). «Влияние источника углерода на экспрессию генов F0 и на стехиометрию субъединицы c в АТФазе F1F0 Escherichia coli » . Дж. Бактериол . 180 (12): 3205–8. дои : 10.1128/jb.180.12.3205-3208.1998 . ПМК 107823 . ПМИД 9620972 .
- ^ Нельсон Н., Перзов Н., Коэн А., Хагай К., Падлер В., Нельсон Х. (1 января 2000 г.). «Клеточная биология генерации протондвижущей силы V-АТФазами» . Дж. Эксп. Биол . 203 (Часть 1): 89–95. дои : 10.1242/jeb.203.1.89 . ПМИД 10600677 . Архивировано из оригинала 30 сентября 2007 года.
- ^ Рубинштейн Дж.Л., Уокер Дж.Э., Хендерсон Р. (2003). «Структура митохондриальной АТФ-синтазы по данным электронной криомикроскопии» . ЭМБО Дж . 22 (23): 6182–92. дои : 10.1093/emboj/cdg608 . ПМК 291849 . ПМИД 14633978 .
- ^ Лесли А.Г., Уокер Дж.Э. (2000). «Структурная модель F1-АТФазы и значение ротационного катализа» . Филос. Пер. Р. Сок. Лонд. Б Биол. Наука . 355 (1396): 465–71. дои : 10.1098/rstb.2000.0588 . ПМК 1692760 . ПМИД 10836500 .
- ^ Ноджи Х., Ёсида М. (2001). «Вращающаяся машина в клетке, АТФ-синтаза» . Ж. Биол. Хим . 276 (3): 1665–8. дои : 10.1074/jbc.R000021200 . ПМИД 11080505 . S2CID 30953216 .
- ^ Капальди Р.А., Аггелер Р. (2002). «Механизм АТФ-синтазы F (1) F (0)-типа, биологического вращательного двигателя». Тенденции биохимии. Наука . 27 (3): 154–60. дои : 10.1016/S0968-0004(01)02051-5 . ПМИД 11893513 .
- ^ Димрот П., фон Баллмос С., Мейер Т. (2006). «Каталитические и механические циклы в синтезе F-АТФ. Четвертый в серии обзоров циклов» . Представитель ЭМБО . 7 (3): 276–82. дои : 10.1038/sj.embor.7400646 . ПМЦ 1456893 . ПМИД 16607397 .
- ↑ Перейти обратно: Перейти обратно: а б Грессер М.Дж., Майерс Дж.А., Бойер П.Д. (25 октября 1982 г.). «Кооперативность каталитических участков митохондриальной аденозинтрифосфатазы F1 говяжьего сердца. Корреляции начальной скорости, связанных промежуточных продуктов и измерений кислородного обмена с чередующейся трехсайтовой моделью» . Ж. Биол. Хим . 257 (20): 12030–8. дои : 10.1016/S0021-9258(18)33672-X . ПМИД 6214554 . Архивировано из оригинала 29 сентября 2007 года.
- ^ Димрот П. (1994). «Бактериальная ионно-связанная энергетика натрия». Антони ван Левенгук . 65 (4): 381–95. дои : 10.1007/BF00872221 . ПМИД 7832594 . S2CID 23763996 .
- ↑ Перейти обратно: Перейти обратно: а б Бехер Б, Мюллер В (1994). «Дельта мю Na+ управляет синтезом АТФ посредством дельта мю Na(+)-транслокирующей F1F0-АТФ-синтазы в мембранных везикулах археи Methanosarcina mazei Gö1» . Дж. Бактериол . 176 (9): 2543–50. дои : 10.1128/jb.176.9.2543-2550.1994 . ПМК 205391 . ПМИД 8169202 .
- ^ Мюллер В. (2004). «Исключительная изменчивость мотора АТФаз архаила A1A0: от мультимерных до мономерных роторов, содержащих 6-13 сайтов связывания ионов». Дж. Биоэнергетика. Биомембр . 36 (1): 115–25. дои : 10.1023/B:JOBB.0000019603.68282.04 . ПМИД 15168615 . S2CID 24887884 .
- ↑ Перейти обратно: Перейти обратно: а б Дэвис К.Дж. (1995). «Окислительный стресс: парадокс аэробной жизни». Биохим. Соц. Симп . 61 : 1–31. дои : 10.1042/bss0610001 . ПМИД 8660387 .
- ^ Ротанг СИ (2006). «Теории биологического старения: гены, белки и свободные радикалы» (PDF) . Свободный Радик. Рез . 40 (12): 1230–8. CiteSeerX 10.1.1.476.9259 . дои : 10.1080/10715760600911303 . ПМИД 17090411 . S2CID 11125090 . Архивировано из оригинала (PDF) 14 июня 2014 г. Проверено 27 октября 2017 г.
- ^ Валко М., Лейбфриц Д., Монкол Дж., Кронин М.Т., Мазур М., Тельсер Дж. (2007). «Свободные радикалы и антиоксиданты в нормальных физиологических функциях и заболеваниях человека». Межд. Дж. Биохим. Клеточная Биол . 39 (1): 44–84. doi : 10.1016/j.biocel.2006.07.001 . ПМИД 16978905 .
- ^ Раха С., Робинсон Б.Х. (2000). «Митохондрии, свободные радикалы кислорода, болезни и старение». Тенденции биохимии. Наука . 25 (10): 502–8. дои : 10.1016/S0968-0004(00)01674-1 . ПМИД 11050436 .
- ^ Финкель Т., Холбрук, штат Нью-Джерси (2000). «Окислители, окислительный стресс и биология старения». Природа . 408 (6809): 239–47. Бибкод : 2000Natur.408..239F . дои : 10.1038/35041687 . ПМИД 11089981 . S2CID 2502238 .
- ^ Каденбах Б., Рамзан Р., Вэнь Л., Фогт С. (март 2010 г.). «Новое расширение теории Митчелла окислительного фосфорилирования в митохондриях живых организмов». Биохим. Биофиз. Акта . 1800 (3): 205–12. дои : 10.1016/j.bbagen.2009.04.019 . ПМИД 19409964 .
- ^ Эхтай К.С., Руссель Д., Сен-Пьер Дж., Джекабсонс М.Б., Каденас С., Стюарт Дж.А., Харпер Дж.А., Робак С.Дж., Моррисон А., Пикеринг С., Клэпхэм Дж.К., Брэнд, доктор медицинских наук (январь 2002 г.). «Супероксид активирует митохондриальные разобщающие белки». Природа . 415 (6867): 96–9. Бибкод : 2002Natur.415...96E . дои : 10.1038/415096а . ПМИД 11780125 . S2CID 4349744 .
- ^ Дево Дж.Б., Хеджес С.П., Берч Н., Герберт Н., Реншоу ГМ, Хики А.Дж. (январь 2019 г.). «Ацидоз поддерживает функцию митохондрий мозга у толерантных к гипоксии трехплефиновых рыб: стратегия выживания при остром гипоксическом воздействии?» . Границы в физиологии . 9 : 1941. doi : 10.3389/fphys.2018.01941 . ПМК 6346031 . ПМИД 30713504 .
- ↑ Перейти обратно: Перейти обратно: а б Джоши С., Хуан Ю.Г. (1991). «Комплекс АТФ-синтазы из митохондрий бычьего сердца: белок, придающий чувствительность к олигомицину, необходим для АТФазы, чувствительной к дициклогексилкарбодиимиду». Биохим. Биофиз. Акта . 1067 (2): 255–8. дои : 10.1016/0005-2736(91)90051-9 . ПМИД 1831660 .
- ↑ Перейти обратно: Перейти обратно: а б Сатьянараяна У (2002). Биохимия (2-е изд.). Калькутта, Индия: Книги и сопутствующие товары. ISBN 8187134801 . OCLC 71209231 .
- ^ Цубаки М (1993). «Инфракрасное исследование с Фурье-преобразованием связывания цианида с биядерным сайтом Fea3-CuB цитохром с-оксидазы бычьего сердца: влияние окислительно-восстановительных конформационных изменений в биядерном сайте». Биохимия . 32 (1): 164–73. дои : 10.1021/bi00052a022 . ПМИД 8380331 .
- ^ Гейтлер П.Г. (1979). «Разобщители окислительного фосфорилирования». У Сидни Флейшера, Лестер Пакера (ред.). Биомембраны. Часть F: Биоэнергетика: окислительное фосфорилирование . Методы энзимологии. Том. 55. стр. 462–472. дои : 10.1016/0076-6879(79)55060-5 . ISBN 978-0-12-181955-2 . ПМИД 156853 .
- ^ Ламберт А.Дж., Бренд, доктор медицинских наук (2004). «Ингибиторы хинон-связывающего сайта позволяют быстро производить супероксид из митохондриальной НАДН:убихиноноксидоредуктазы (комплекс I)» . Ж. Биол. Хим . 279 (38): 39414–20. дои : 10.1074/jbc.M406576200 . ПМИД 15262965 . S2CID 26620903 .
- ^ Дервартанян Д.В., Вигер С. (ноябрь 1964 г.). «Исследование сукцинатдегидрогеназы: I. Спектральные свойства очищенного фермента и образование фермент-конкурентных ингибиторных комплексов». Биохим. Биофиз. Акта . 92 (2): 233–47. дои : 10.1016/0926-6569(64)90182-8 . ПМИД 14249115 .
- ^ Рикье Д., Буйо Ф. (2000). «Гомологи разобщающих белков: UCP1, UCP2, UCP3, StUCP и AtUCP» . Биохим. Дж . 345 (2): 161–79. дои : 10.1042/0264-6021:3450161 . ПМК 1220743 . ПМИД 10620491 .
- ^ Борецкий Дж., Верчези А.Е. (2005). «Растительный разобщающий митохондриальный белок и альтернативная оксидаза: энергетический метаболизм и стресс» . Биология. Представитель . 25 (3–4): 271–86. дои : 10.1007/s10540-005-2889-2 . ПМИД 16283557 . S2CID 18598358 .
- ^ Харден А., Янг У.Дж. (1906). «Алкогольная закваска дрожжевого сока» . Труды Королевского общества . Б (77): 405–20. дои : 10.1098/rspb.1906.0029 .
- ^ Калькар Х.М. (1974). «Истоки понятия окислительного фосфорилирования». Мол. Клетка. Биохим . 5 (1–2): 55–63. дои : 10.1007/BF01874172 . ПМИД 4279328 . S2CID 26999163 .
- ^ Липманн Ф (1941). «Метаболическое образование и использование энергии фосфатных связей». Адв Энзимол . 1 : 99–162. doi : 10.4159/harvard.9780674366701.c141 . ISBN 9780674366701 .
- ^ Фридкин М., Ленингер А.Л. (1 апреля 1949 г.). «Этерификация неорганического фосфата, связанная с транспортом электронов между дигидродифосфопиридиновым нуклеотидом и кислородом» . Ж. Биол. Хим . 178 (2): 611–44. дои : 10.1016/S0021-9258(18)56879-4 . ПМИД 18116985 . Архивировано из оригинала 16 декабря 2008 года.
- ^ Калькар Х.М. (1991). «50 лет биологических исследований — от окислительного фосфорилирования до регулирования транспорта энергии» . Ежегодный обзор биохимии . 60 : 1–37. дои : 10.1146/annurev.bi.60.070191.000245 . ПМИД 1883194 .
- ^ Белицер В.А., Цибакова Е.Т. (1939). «О механизме фосфорилирования, связанном с дыханием». Биохимия . 4 : 516–534.
- ^ Слейтер ЕС (1953). «Механизм фосфорилирования в дыхательной цепи». Природа . 172 (4387): 975–8. Бибкод : 1953Natur.172..975S . дои : 10.1038/172975a0 . ПМИД 13111237 . S2CID 4153659 .
- ^ Митчелл П. (1961). «Сочетание фосфорилирования с переносом электронов и водорода по механизму хемиосмотического типа». Природа . 191 (4784): 144–8. Бибкод : 1961Natur.191..144M . дои : 10.1038/191144a0 . ПМИД 13771349 . S2CID 1784050 .
- ^ Сайер-младший М.Х. Питер Митчелл и жизненная сила . OCLC 55202414 .
- ^ Митчелл П. (1978). «Концепция дыхательной цепи Дэвида Кейлина и ее хемиосмотические последствия» (PDF) . Нобелевская лекция . Нобелевский фонд. Архивировано (PDF) из оригинала 27 сентября 2007 г. Проверено 21 июля 2007 г.
- ^ Пуллман М.Э., Пенефски Х.С., Датта А., Рэкер Э. (1 ноября 1960 г.). «Частичное разрешение ферментов, катализирующих окислительное фосфорилирование. I. Очистка и свойства растворимой динитрофенол-стимулированной аденозинтрифосфатазы» . Ж. Биол. Хим . 235 (11): 3322–9. дои : 10.1016/S0021-9258(20)81361-1 . ПМИД 13738472 . Архивировано из оригинала 29 сентября 2007 года.
- ^ Бойер П.Д., Кросс Р.Л., Момсен В. (1973). «Новая концепция энергетического взаимодействия при окислительном фосфорилировании, основанная на молекулярном объяснении реакций обмена кислорода» . Учеб. Натл. акад. наук. США . 70 (10): 2837–9. Бибкод : 1973PNAS...70.2837B . дои : 10.1073/pnas.70.10.2837 . ПМК 427120 . ПМИД 4517936 .
- ^ «Нобелевская премия по химии 1997 года» . Нобелевский фонд. Архивировано из оригинала 25 марта 2017 г. Проверено 21 июля 2007 г.
Дальнейшее чтение [ править ]
Введение [ править ]
- Нельсон Д.Л., Кокс М.М. (2004). Ленингерские принципы биохимии (4-е изд.). У. Х. Фриман. ISBN 0-7167-4339-6 .
- Шнайдер Э.Д., Саган Д. (2006). В прохладу: поток энергии, термодинамика и жизнь (1-е изд.). Издательство Чикагского университета. ISBN 0-226-73937-6 .
- Лейн Н (2006). Власть, секс, самоубийство: митохондрии и смысл жизни (1-е изд.). Издательство Оксфордского университета, США. ISBN 0-19-920564-7 .
Расширенный [ править ]
- Николлс Д.Г., Фергюсон С.Дж. (2002). Биоэнергетика 3 (1-е изд.). Академическая пресса. ISBN 0-12-518121-3 .
- Хейни Д. (2001). Биологическая термодинамика (1-е изд.). Издательство Кембриджского университета. ISBN 0-521-79549-4 .
- Раджан СС (2003). Введение в биоэнергетику (1-е изд.). Анмол. ISBN 81-261-1364-2 .
- Викстром М, изд. (2005). Биофизические и структурные аспекты биоэнергетики (1-е изд.). Королевское химическое общество. ISBN 0-85404-346-2 .
Общие ресурсы [ править ]
- Анимированные диаграммы, иллюстрирующие окислительное фосфорилирование. Концепции Wiley and Co в биохимии.
- Онлайн-лекции по биофизике Энтони Крофтс, Университет Иллинойса в Урбане-Шампейне
- АТФ-синтаза Грэм Джонсон
Структурные ресурсы [ править ]
- PDB Молекула месяца :
- АТФ-синтаза. Архивировано 24 июля 2020 г. в Wayback Machine.
- Цитохром c. Архивировано 24 июля 2020 г. в Wayback Machine.
- Цитохром с-оксидаза. Архивировано 24 июля 2020 г. в Wayback Machine.
- Интерактивные молекулярные модели в Университете Фернандо Пессоа :
метаболизма Карта |
|---|
