Клеточное дыхание
В этой статье есть несколько проблем. Пожалуйста, помогите улучшить его или обсудите эти проблемы на странице обсуждения . ( Узнайте, как и когда удалять эти шаблонные сообщения )
|
Клеточное дыхание — это процесс, посредством которого биологическое топливо окисляется в присутствии неорганического акцептора электронов , такого как кислород , чтобы стимулировать массовое производство аденозинтрифосфата (АТФ) , который содержит энергию. Клеточное дыхание можно описать как совокупность метаболических которые происходят в клетках организмов реакций и процессов , с целью преобразования химической энергии в питательных веществ АТФ и последующего высвобождения отходов . [1]
Клеточное дыхание — жизненно важный процесс, происходящий в клетках всех живых организмов . [2] [ нужен лучший источник ] Дыхание может быть аэробным, требующим кислорода, или анаэробным; некоторые организмы могут переключаться между аэробным и анаэробным дыханием. [3] [ нужен лучший источник ]
Реакции, участвующие в дыхании, являются катаболическими реакциями , которые расщепляют большие молекулы на более мелкие, производя большое количество энергии (АТФ). Дыхание — один из ключевых способов высвобождения клеткой химической энергии для поддержания клеточной активности. Общая реакция происходит в ряд биохимических стадий, некоторые из которых являются окислительно-восстановительными реакциями. Хотя клеточное дыхание технически является реакцией горения , оно необычно из-за медленного, контролируемого высвобождения энергии в результате серии реакций.
Питательные вещества, которые обычно используются клетками животных и растений при дыхании, включают сахара , аминокислоты и жирные кислоты , а наиболее распространенным окислителем является молекулярный кислород (О 2 ). Химическая энергия, запасенная в АТФ (связь ее третьей фосфатной группы с остальной частью молекулы может быть разорвана, позволяя образовываться более стабильным продуктам, тем самым высвобождая энергию для использования клеткой), может затем использоваться для запуска процессов, требующих энергии, в том числе биосинтез , передвижение или транспортировка молекул через клеточные мембраны .
Аэробное дыхание
[ редактировать ]Аэробное дыхание требует кислорода (O 2 ) для создания АТФ . Хотя углеводы , жиры и белки потребляются в качестве реагентов , аэробное дыхание является предпочтительным методом производства пирувата при гликолизе и требует поступления пирувата в митохондрии для полного окисления в цикле лимонной кислоты . Продуктами этого процесса являются углекислый газ и вода, а передаваемая энергия используется для образования связей между АДФ и третьей фосфатной группой с образованием АТФ ( аденозинтрифосфата ) путем фосфорилирования на уровне субстрата , НАДН и ФАДН 2 . [ нужна ссылка ]
| Массовый баланс глобальной реакции: | C 6 H 12 O 6 (т) + 6 O 2 (г) → 6 CO 2 (г) + 6 H 2 O (л) + энергия |
| ΔG = −2880 кДж на моль C 6 H 12 O 6 |
Отрицательное значение ΔG указывает на то, что реакция экзотермическая ( экзергоническая ) и может протекать самопроизвольно. [4]
Потенциал НАДН и ФАДН 2 преобразуется в большее количество АТФ через цепь переноса электронов с кислородом и протонами (водородом) в качестве « концевых акцепторов электронов ». Большая часть АТФ, вырабатываемая аэробным клеточным дыханием, производится путем окислительного фосфорилирования . Высвобождаемая энергия используется для создания хемиосмотического потенциала путем перекачки протонов через мембрану. Этот потенциал затем используется для управления АТФ-синтазой и производства АТФ из АДФ и фосфатной группы. В учебниках по биологии часто указывается, что на каждую окисленную молекулу глюкозы в ходе клеточного дыхания может быть получено 38 молекул АТФ (2 — в результате гликолиза, 2 — в цикле Кребса и около 34 — в системе транспорта электронов). [5] Однако этот максимальный выход никогда полностью не достигается из-за потерь из-за негерметичности мембран , а также из-за стоимости перемещения пирувата и АДФ в митохондриальный матрикс, и текущие оценки колеблются от 29 до 30 АТФ на глюкозу. [5]
Аэробный метаболизм до 15 раз эффективнее анаэробного метаболизма (при котором на 1 молекулу глюкозы образуется 2 молекулы АТФ). Однако некоторые анаэробные организмы, такие как метаногены, способны продолжать анаэробное дыхание , производя больше АТФ, используя неорганические молекулы, отличные от кислорода, в качестве конечных акцепторов электронов в цепи переноса электронов. Они разделяют начальный путь гликолиза , но аэробный метаболизм продолжается с помощью цикла Кребса и окислительного фосфорилирования. Постгликолитические реакции протекают в митохондриях эукариотических клеток и в цитоплазме прокариотических клеток . [ нужна ссылка ]
Хотя растения являются чистыми потребителями углекислого газа и производителями кислорода посредством фотосинтеза , на дыхание растений приходится около половины CO 2 , ежегодно вырабатываемого наземными экосистемами . [6] [7] : 87
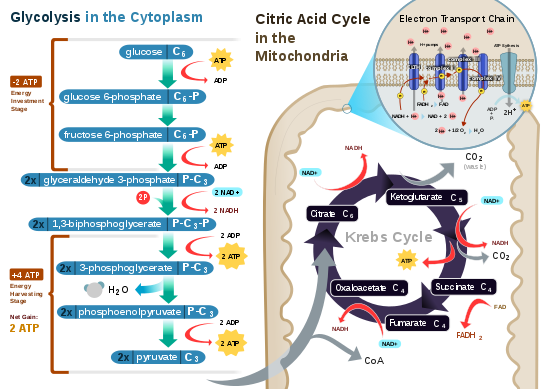
Гликолиз
[ редактировать ]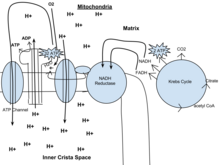
Гликолиз — это метаболический путь , который происходит в цитозоле клеток всех живых организмов. Гликолиз можно дословно перевести как «расщепление сахара». [8] и происходит независимо от присутствия или отсутствия кислорода. В аэробных условиях этот процесс превращает одну молекулу глюкозы в две молекулы пирувата (пировиноградной кислоты), генерируя энергию в виде двух чистых молекул АТФ . Фактически производятся четыре молекулы АТФ на глюкозу, но две расходуются в рамках подготовительной фазы . Первоначальное фосфорилирование глюкозы необходимо для повышения реакционной способности (снижения ее стабильности) для того, чтобы молекула могла расщепиться на две молекулы пирувата ферментом альдолазой . Во время фазы гликолиза четыре фосфатные группы передаются на четыре АДФ путем фосфорилирования на уровне субстрата с образованием четырех АТФ, а две НАДН образуются при пирувата окислении . Общую реакцию можно выразить так: [ нужна ссылка ]
- Глюкоза + 2 НАД + + 2 P i + 2 АДФ → 2 пируват + 2 НАДН + 2 АТФ + 2 Н + + 2 H 2 O + энергия
Начиная с глюкозы, 1 АТФ используется для передачи фосфата глюкозе с образованием глюкозо-6-фосфата . Гликоген также может превращаться в глюкозо-6-фосфат с помощью гликогенфосфорилазы . В ходе энергетического обмена глюкозо-6-фосфат превращается в фруктозо-6-фосфат . Дополнительный АТФ используется для фосфорилирования фруктозо-6-фосфата во фруктозо-1,6-бисфосфат с помощью фосфофруктокиназы . Затем фруктозо-1,6-бифосфат расщепляется на две фосфорилированные молекулы с тремя углеродными цепями, которые позже разлагаются до пирувата. [7] : 88–90
Окислительное декарбоксилирование пирувата
[ редактировать ]Пируват окисляется до ацетил-КоА и CO 2 ( пируватдегидрогеназным комплексом PDC). PDC содержит множественные копии трех ферментов и расположен в митохондриях эукариотических клеток и в цитозоле прокариот. При превращении пирувата в ацетил-КоА одна молекула НАДН и одна молекула СО 2 . образуется [ нужна ссылка ]
Цикл лимонной кислоты
[ редактировать ]Цикл лимонной кислоты также называют циклом Кребса или циклом трикарбоновых кислот . Когда присутствует кислород, ацетил-КоА образуется из молекул пирувата, образующихся в результате гликолиза. После ацетил-КоА образования может возникнуть аэробное или анаэробное дыхание. При наличии кислорода митохондрии подвергаются аэробному дыханию, которое приводит к циклу Кребса. Однако если кислорода нет, произойдет ферментация молекулы пирувата. В присутствии кислорода, когда образуется ацетил-КоА, молекула затем вступает в цикл лимонной кислоты (цикл Кребса) внутри митохондриального матрикса и окисляется до CO 2 , одновременно восстанавливая НАД до НАДН . НАДН может использоваться цепью переноса электронов для создания дальнейшего АТФ в рамках окислительного фосфорилирования. Чтобы полностью окислить эквивалент одной молекулы глюкозы, два ацетил-КоА должны метаболизироваться в цикле Кребса. два низкоэнергетических отходов : H 2 O и CO 2 . В ходе этого цикла образуются [9] [10]
Цикл лимонной кислоты представляет собой 8-ступенчатый процесс, в котором участвуют 18 различных ферментов и коферментов. В ходе цикла ацетил-КоА (2 атома углерода) + оксалоацетат (4 атома углерода) дает цитрат (6 атомов углерода), который перегруппировывается в более реакционноспособную форму, называемую изоцитратом (6 атомов углерода). Изоцитрат модифицируется с образованием α-кетоглутарата (5 атомов углерода), сукцинил-КоА , сукцината , фумарата , малата и, наконец, оксалоацетата . [ нужна ссылка ]
Чистый прирост от одного цикла составляет 3 НАДН и 1 ФАДН 2 в виде соединений, несущих водород (протон плюс электрон), и 1 высокоэнергетический ГТФ , который впоследствии может быть использован для производства АТФ. Таким образом, общий выход из 1 молекулы глюкозы (2 молекул пирувата) составляет 6 НАДН, 2 ФАДН 2 и 2 АТФ. [9] [10] [7] : 90–91
Окислительное фосфорилирование
[ редактировать ]У эукариот окислительное фосфорилирование происходит в кристах митохондрий . Он включает в себя цепь переноса электронов, которая устанавливает протонный градиент (хемиосмотический потенциал) через границу внутренней мембраны путем окисления НАДН, образующегося в цикле Кребса. АТФ синтезируется ферментом АТФ-синтазой, когда хемиосмотический градиент используется для управления фосфорилированием АДФ. Электроны в конечном итоге передаются экзогенному кислороду и при присоединении двух протонов образуется вода. [ нужна ссылка ]
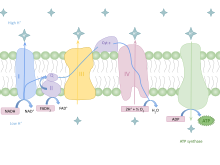
Эффективность производства АТФ
[ редактировать ]В таблице ниже описаны реакции, происходящие при полном окислении одной молекулы глюкозы в углекислый газ. Предполагается, что все восстановленные коферменты окисляются по цепи переноса электронов и используются для окислительного фосфорилирования.
| Шаг | выход кофермента | выход АТФ | Источник АТФ |
|---|---|---|---|
| Подготовительный этап гликолиза | −2 | Фосфорилирование глюкозы и фруктозо-6-фосфата использует два АТФ из цитоплазмы. | |
| Фаза окупаемости гликолиза | 4 | Фосфорилирование на уровне субстрата | |
| 2 НАДН | 3 или 5 | Окислительное фосфорилирование: каждый НАДН производит чистые 1,5 АТФ (вместо обычных 2,5) за счет транспорта НАДН через митохондриальную мембрану. | |
| Окислительное декарбоксилирование пирувата | 2 НАДН | 5 | Окислительное фосфорилирование |
| Цикл Кребса | 2 | Фосфорилирование на уровне субстрата | |
| 6 НАДН | 15 | Окислительное фосфорилирование | |
| 2 FADH2ФАДХ2 | 3 | Окислительное фосфорилирование | |
| Общий доход | 30 или 32 АТФ | От полного окисления одной молекулы глюкозы до углекислого газа и окисления всех восстановленных коферментов. | |
Хотя теоретический выход составляет 38 молекул АТФ на глюкозу во время клеточного дыхания, такие условия обычно не реализуются из-за таких потерь, как стоимость перемещения пирувата (в результате гликолиза), фосфата и АДФ (субстратов для синтеза АТФ) в митохондрии. . Все они активно транспортируются с помощью переносчиков, которые используют запасенную энергию в электрохимическом градиенте протонов .
- Пируват захватывается специфическим переносчиком с низким K m и переносит его в митохондриальный матрикс для окисления комплексом пируватдегидрогеназы.
- Фосфатный переносчик (PiC) опосредует электронейтральный обмен ( антипорт ) фосфата ( ЧАС 2 ПО - 4 ; P i ) для ОН − или симпорт фосфата и протонов (H + ) через внутреннюю мембрану, а движущей силой перемещения ионов фосфата в митохондрии является движущая сила протонов .
- Транслоказа АТФ -АДФ (также называемая адениннуклеотидтранслоказой, ANT ) является антипортером и обменивает АДФ и АТФ через внутреннюю мембрану . Движущая сила возникает из-за того, что АТФ (-4) имеет более отрицательный заряд, чем АДФ (-3), и, таким образом, он рассеивает часть электрического компонента протонного электрохимического градиента.
Результатом этих процессов переноса с использованием электрохимического градиента протонов является то, что более 3 H + необходимы для образования 1 АТФ. Очевидно, это снижает теоретическую эффективность всего процесса и вероятный максимум приближается к 28–30 молекулам АТФ. [5] На практике эффективность может быть еще ниже, поскольку внутренняя мембрана митохондрий слегка пропускает протоны. [11] Другие факторы также могут рассеивать протонный градиент, создавая предположительно «протекающие» митохондрии. Разобщающий белок, известный как термогенин, экспрессируется в некоторых типах клеток и является каналом, который может транспортировать протоны. Когда этот белок активен во внутренней мембране, он замыкает связь между цепью переноса электронов и синтезом АТФ . Потенциальная энергия протонного градиента не используется для производства АТФ, а генерирует тепло. Это особенно важно для термогенеза бурого жира у новорожденных и зимующих млекопитающих.
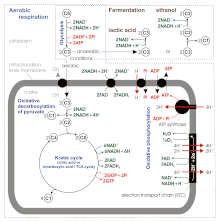
По некоторым более новым источникам, выход АТФ при аэробном дыхании составляет не 36–38, а всего около 30–32 молекул АТФ/1 молекула глюкозы. [12] , потому что:
- АТФ: НАДН+Н + Соотношения АТФ : ФАДН 2 при окислительном фосфорилировании оказываются не 3 и 2, а 2,5 и 1,5 соответственно. В отличие от фосфорилирования на уровне субстрата , стехиометрию здесь сложно установить .
- АТФ-синтаза производит 1 АТФ/3 Н. + . Однако обмен матриксного АТФ на цитозольный АДФ и Pi (антипорт с ОН − или симпортировать с помощью H + ) опосредованный транслоказой АТФ-АДФ и фосфатным переносчиком, потребляет 1 H + / 1 АТФ в результате регенерации трансмембранного потенциала, измененного во время этого переноса, поэтому чистое соотношение составляет 1 АТФ : 4 H. + .
- митохондриальной цепи переноса электронов Протонный насос переносит через внутреннюю мембрану 10 H + / 1 НАДН+Н + (4 + 2 + 4) или 6 Н + /1 ФАДХ 2 (2 + 4).
- Итак, окончательная стехиометрия равна
- 1 НАДН+Н + : 10 ч. + : 10/4 АТФ = 1 НАДН+Н + : 2,5 АТФ
- 1 ФАДХ 2 : 6 Ч + : 6/4 АТФ = 1 ФАДН 2 : 1,5 АТФ
- АТФ: НАДН+Н + исходящий из коэффициента гликолиза при окислительном фосфорилировании,
- 1,5, как для ФАДН 2 , если атомы водорода (2H + +2е − ) переносятся из цитозоля НАДН+Н + к митохондриальному FAD с помощью глицеринфосфатного челнока, расположенного во внутренней мембране митохондрий.
- 2.5 в случае малатно-аспартатного челнока, переносящего атомы водорода от цитозольного НАДН+Н + это митохондриальный НАД +
В итоге мы имеем на молекулу глюкозы
- Субстратное фосфорилирование : 2 АТФ из гликолиза + 2 АТФ (непосредственно ГТФ) из цикла Кребса .
- Окислительное фосфорилирование
- 2 НАДН+Н + от гликолиза: 2×1,5 АТФ (если глицеринфосфатный челнок переносит атомы водорода) или 2×2,5 АТФ (малатно-аспартатный челнок)
- 2 НАДН+Н + от окислительного декарбоксилирования пирувата и 6 из цикла Кребса: 8 × 2,5 АТФ
- 2 ФАДГ 2 из цикла Кребса: 2 × 1,5 АТФ
В сумме это дает 4 + 3 (или 5) + 20 + 3 = 30 (или 32) АТФ на молекулу глюкозы.
Эти цифры могут потребовать дальнейшей корректировки по мере появления новых структурных деталей. Вышеуказанное значение 3 H + / АТФ для синтазы предполагает, что синтаза перемещает 9 протонов и производит 3 АТФ за один оборот. Число протонов зависит от количества c-субъединиц в Fo c-кольце , и теперь известно, что у дрожжей Fo оно равно 10. [13] и 8 для позвоночных. [14] В том числе один Н. + для реакций транспорта это означает, что для синтеза одной АТФ требуется 1 + 10/3 = 4,33 протона у дрожжей и 1 + 8/3 = 3,67 у позвоночных . Это означало бы, что в митохондриях человека 10 протонов при окислении НАДН будут производить 2,72 АТФ (вместо 2,5), а 6 протонов при окислении сукцината или убихинола будут производить 1,64 АТФ (вместо 1,5). Это согласуется с экспериментальными результатами в пределах погрешности, описанными в недавнем обзоре. [15]
Общий выход АТФ при этанольном или молочнокислом брожении составляет всего 2 молекулы, поступающие в результате гликолиза , поскольку пируват не переносится в митохондрии и окончательно не окисляется до углекислого газа (СО 2 ), а восстанавливается до этанола или молочной кислоты в цитоплазме . [12]
Ферментация
[ редактировать ]Без кислорода пируват ( пировиноградная кислота ) не метаболизируется при клеточном дыхании, а подвергается процессу ферментации . Пируват не транспортируется в митохондрии, а остается в цитоплазме, где превращается в продукты жизнедеятельности , которые могут быть удалены из клетки. Это служит цели окисления переносчиков электронов, чтобы они могли снова выполнять гликолиз, и удаления избытка пирувата. Ферментация окисляет НАДН до НАД. + поэтому его можно повторно использовать в гликолизе. В отсутствие кислорода ферментация предотвращает накопление НАДН в цитоплазме и обеспечивает НАД. + для гликолиза. Этот продукт жизнедеятельности варьируется в зависимости от организма. В скелетных мышцах продуктом жизнедеятельности является молочная кислота . Этот тип брожения называется молочнокислым брожением . При напряженных физических нагрузках, когда потребности в энергии превышают запасы энергии, дыхательная цепь не может переработать все атомы водорода, к которым присоединен НАДН. Во время анаэробного гликолиза НАД + регенерирует, когда пары водорода соединяются с пируватом с образованием лактата. Образование лактата катализируется лактатдегидрогеназой в обратимой реакции. Лактат также можно использовать в качестве непрямого предшественника гликогена печени. Во время восстановления, когда кислород становится доступным, НАД + присоединяется к водороду из лактата с образованием АТФ. У дрожжей отходами являются этанол и углекислый газ . Этот тип ферментации известен как алкогольная или этаноловая ферментация . АТФ, образующийся в этом процессе, производится путем фосфорилирования на уровне субстрата , для которого не требуется кислород.
Ферментация менее эффективно использует энергию глюкозы: на глюкозу вырабатывается только 2 АТФ по сравнению с 38 АТФ на глюкозу, номинально вырабатываемыми при аэробном дыхании. Однако гликолитический АТФ вырабатывается быстрее. Чтобы прокариоты могли продолжать быстрый рост при переходе из аэробной среды в анаэробную, им необходимо увеличить скорость гликолитических реакций. У многоклеточных организмов во время коротких всплесков напряженной деятельности мышечные клетки используют ферментацию для дополнения выработки АТФ в результате более медленного аэробного дыхания, поэтому ферментация может использоваться клеткой даже до того, как уровень кислорода истощится, как это имеет место в видах спорта, где не требовать от спортсменов поддержания темпа, например, при спринте .
Анаэробное дыхание
[ редактировать ]Клеточное дыхание — это процесс, при котором биологическое топливо окисляется в присутствии неорганического акцептора электронов, такого как кислород, с образованием большого количества энергии и стимулирования массового производства АТФ.
Анаэробное дыхание используется микроорганизмами, бактериями или архей, у которых ни кислород (аэробное дыхание), ни производные пирувата (брожение) не являются конечными акцепторами электронов. Скорее, неорганический акцептор, такой как сульфат ( SO 2− 4 ), нитрат ( NO - 3 ) или серу (S). [16] Такие организмы можно встретить в необычных местах, например, в подводных пещерах или возле гидротермальных источников на дне океана. [7] : 66–68 а также в бескислородных почвах или отложениях водно-болотных экосистем.
В июле 2019 года в ходе научного исследования шахты Кидд в Канаде были обнаружены серодышащие организмы , которые живут на глубине 7900 футов (2400 метров) под поверхностью земли и дышат серой, чтобы выжить. Эти организмы также примечательны тем, что потребляют такие минералы, как пирит, в качестве источника пищи. [17] [18] [19]
См. также
[ редактировать ]- Поддерживающее дыхание : поддержание как функциональный компонент клеточного дыхания.
- Микрофизиометрия
- Пастор Пойнт
- Респирометрия : исследовательский инструмент для изучения клеточного дыхания.
- Тетразолия хлорид : индикатор клеточного дыхания.
- Комплекс 1 : НАДН: оксидоредукты убихинона.
Ссылки
[ редактировать ]- ^ Бэйли, Регина. «Клеточное дыхание» . Архивировано из оригинала 5 мая 2012 г.
- ^ «Клеточное дыхание и почему оно важно - Дыхание - AQA Synergy - Комбинированный научный пересмотр GCSE - AQA Synergy - BBC Bitesize» . www.bbc.co.uk. Проверено 7 декабря 2023 г.
- ^ «2.30 Анаэробное и аэробное дыхание» .
- ^ «Сколько АТФ образуется при аэробном дыхании» .
- ^ Jump up to: а б с Рич, PR (2003). «Молекулярный механизм дыхательной цепи Кейлина» . Труды Биохимического общества . 31 (Часть 6): 1095–1105. дои : 10.1042/BST0311095 . ПМИД 14641005 .
- ^ О'Лири, Брендан М.; Плакстон, Уильям К. (2016). «Дыхание растений». ЭЛС . стр. 1–11. дои : 10.1002/9780470015902.a0001301.pub3 . ISBN 9780470016176 .
- ^ Jump up to: а б с д Маннион, AM (12 января 2006 г.). Углерод и его приручение . Спрингер. ISBN 978-1-4020-3956-0 .
- ^ Рис, Джейн; Урри, Лиза; Каин, Майкл; Вассерман, Стивен; Минорский, Петр; Джексон, Роберт (2010). Кэмпбелл Биология, девятое издание . Pearson Education, Inc. с. 168.
- ^ Jump up to: а б Р. Каспи (14 ноября 2012 г.). «Путь: цикл ТСА III (животные)» . База данных метаболических путей MetaCyc . Проверено 20 июня 2022 г.
- ^ Jump up to: а б Р. Каспи (19 декабря 2011 г.). «Путь: цикл TCA I (прокариотический)» . База данных метаболических путей MetaCyc . Проверено 20 июня 2022 г.
- ^ Портер, Р.; Бранд, М. (1 сентября 1995 г.). «Митохондриальная протонная проводимость и H + /O не зависят от скорости транспорта электронов в изолированных гепатоцитах» . The Biochemical Journal (бесплатный полный текст). 310 (Pt 2): 379–382. doi : /bj3100379 . ISSN 0264-6021 . PMC 1135905. 10.1042 PMID 7654171 .
- ^ Jump up to: а б с Страйер, Люберт (1995). Биохимия (четвертое изд.). Нью-Йорк – Бейзингсток: WH Freeman and Company. ISBN 978-0716720096 .
- ^ Сток, Даниэла; Лесли, Эндрю Г.В.; Уокер, Джон Э. (1999). «Молекулярная архитектура роторного двигателя в АТФ-синтазе». Наука . 286 (5445): 1700–5. дои : 10.1126/science.286.5445.1700 . ПМИД 10576729 .
- ^ Ватт, Ян Н.; Монтгомери, Мартин Г.; Рансуик, Майкл Дж.; Лесли, Эндрю Г.В.; Уокер, Джон Э. (2010). «Биоэнергетическая стоимость создания молекулы аденозинтрифосфата в митохондриях животных» . Учеб. Натл. акад. наук. США . 107 (39): 16823–16827. дои : 10.1073/pnas.1011099107 . ПМЦ 2947889 . ПМИД 20847295 .
- ^ П.Хинкль (2005). «Соотношения P/O митохондриального окислительного фосфорилирования». Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1706 (1–2): 1–11. дои : 10.1016/j.bbabio.2004.09.004 . ПМИД 15620362 .
- ^ Люмен: Безграничная микробиология. «Анаэробное дыхание: доноры и акцепторы электронов при анаэробном дыхании» . Courses.lumenlearning.org . Boundless.com . Проверено 19 ноября 2020 г.
Анаэробное дыхание – это образование АТФ без кислорода. Этот метод по-прежнему включает дыхательную цепь транспорта электронов, но без использования кислорода в качестве конечного акцептора электронов. Вместо этого такие молекулы, как сульфат ( SO 2− 4 ), нитрат ( NO - 3 ) или сера (S) используются в качестве акцепторов электронов.
- ^ Лоллар, Гранат С.; Уорр, Оливер; Рассказываю, Джон; Осберн, Магдалена Р.; Шервуд Лоллар, Барбара (2019). « Следуй за водой: гидрогеохимические ограничения на микробные исследования на глубине 2,4 км под поверхностью в Обсерватории глубоких жидкостей и глубокой жизни Кидд-Крик». Геомикробиологический журнал . 36 (10): 859–872. Бибкод : 2019GmbJ...36..859L . дои : 10.1080/01490451.2019.1641770 . S2CID 199636268 .
- ↑ Старейшие в мире подземные воды поддерживают жизнь благодаря химическому составу воды и горных пород. Архивировано 10 сентября 2019 г. в Wayback Machine , 29 июля 2019 г., deepcarbon.net.
- ↑ Странные формы жизни, обнаруженные глубоко в шахте, указывают на огромные «подземные Галапагосы». Архивировано 9 сентября 2019 г. в Wayback Machine , Кори С. Пауэлл, 7 сентября 2019 г., nbcnews.com.
Внешние ссылки
[ редактировать ]- Подробное описание дыхания и ферментации.
- Интернет-ресурс Кимбалла по клеточному дыханию
- Клеточное дыхание и ферментация в Клермонском колледже
метаболизма Карта |
|---|

| Базы данных органов управления : Национальные |
|---|