Электрохимический градиент

Электрохимический градиент — это градиент электрохимического потенциала , обычно для иона , который может перемещаться через мембрану . Градиент состоит из двух частей:
- Химический градиент или разница в концентрации растворенного вещества на мембране.
- Электрический градиент или разница зарядов на мембране.
Когда концентрации ионов на проницаемой мембране неравны, ион будет перемещаться через мембрану из области с более высокой концентрацией в область с более низкой концентрацией посредством простой диффузии . Ионы также несут электрический заряд, который формирует электрический потенциал на мембране. Если существует неравномерное распределение зарядов по мембране, то разница в электрическом потенциале создает силу, которая вызывает диффузию ионов до тех пор, пока заряды не уравновесятся по обе стороны мембраны.
Электрохимические градиенты необходимы для работы батарей и других электрохимических элементов , фотосинтеза и клеточного дыхания , а также некоторых других биологических процессов.
Обзор
[ редактировать ]Электрохимическая энергия — одна из многих взаимозаменяемых форм потенциальной энергии, посредством которой энергия может сохраняться . Он появляется в электроаналитической химии и находит промышленное применение, например, в батареях и топливных элементах. В биологии электрохимические градиенты позволяют клеткам контролировать направление движения ионов через мембраны. В митохондриях и хлоропластах используемый протонные градиенты генерируют хемиосмотический потенциал, для синтеза АТФ . [1] а градиент натрия и калия помогает нейронным синапсам быстро передавать информацию. [ нужна ссылка ]
Электрохимический градиент состоит из двух компонентов: разной концентрации электрического заряда на мембране и разной концентрации химических веществ на той же мембране. В первом случае концентрированный заряд притягивает заряды противоположного знака; в последнем случае концентрированные вещества имеют тенденцию диффундировать через мембрану до выравнивания концентраций. Сочетание этих двух явлений определяет термодинамически предпочтительное направление движения иона через мембрану. [2] : 403 [3]
Суммарный эффект можно количественно оценить как градиент термодинамического электрохимического потенциала : [ нужна ссылка ] с

- μ i химический потенциал вида ионов i
- z i заряд на ион вида i
- F , постоянная Фарадея (электрохимический потенциал измеряется в неявном виде в пермольных единицах )
- φ , местный электрический потенциал .
Иногда термином «электрохимический потенциал» злоупотребляют для описания электрического потенциала, создаваемого градиентом концентрации ионов; то есть φ .
Электрохимический градиент аналогичен давлению воды на плотине гидроэлектростанции . Маршруты, разблокированные мембраной (например, мембранный транспортный белок или электроды ), соответствуют турбинам, которые преобразуют потенциальную энергию воды в другие формы физической или химической энергии, а ионы, которые проходят через мембрану, соответствуют воде, перемещающейся в нижнее течение реки. [ тон ] И наоборот, энергия может использоваться для перекачки воды в озеро над плотиной , а химическая энергия может использоваться для создания электрохимических градиентов. [4] [5]
Химия
[ редактировать ]Этот термин обычно применяется в электрохимии , когда электрическая энергия в виде приложенного напряжения используется для модуляции термодинамической благоприятности химической реакции . В аккумуляторе электрохимический потенциал, возникающий в результате движения ионов, уравновешивает энергию реакции электродов. Максимальное напряжение, которое может произвести реакция батареи, иногда называют стандартным электрохимическим потенциалом этой реакции.
Биологический контекст
[ редактировать ]Генерация трансмембранного электрического потенциала посредством движения ионов через клеточную мембрану управляет биологическими процессами, такими как нервная проводимость, сокращение мышц , гормонов секреция и ощущения . Условно физиологические напряжения измеряются относительно внеклеточной области; типичная животная клетка имеет внутренний электрический потенциал (-70)–(-50) мВ. [2] : 464
Электрохимический градиент необходим для митохондрий окислительного фосфорилирования . Заключительным этапом клеточного дыхания является цепь переноса электронов , состоящая из четырех комплексов, встроенных во внутреннюю мембрану митохондрий. Комплексы I, III и IV перекачивают протоны из матрикса в межмембранное пространство (ИМП); на каждую электронную пару , вступающую в цепочку, в IMS перемещается десять протонов. В результате получается электрический потенциал более 200 мВ . Возникающий в результате поток протонов обратно в матрикс усиливает усилия АТФ-синтазы по объединению неорганического фосфата и АДФ . [6] [2] : 743–745
Подобно цепи переноса электронов, светозависимые реакции фотосинтеза перекачивают протоны в тилакоидов просвет хлоропластов, чтобы стимулировать синтез АТФ. Протонный градиент может быть создан посредством нециклического или циклического фотофосфорилирования. Из белков, участвующих в нециклическом фотофосфорилировании, фотосистема II (PSII), пластихинон и цитохрома b6f комплекс непосредственно способствуют генерации протонного градиента. На каждые четыре фотона, поглощенные PSII, в просвет закачивается восемь протонов. [2] : 769–770
Несколько других транспортеров и ионных каналов играют роль в создании электрохимического градиента протонов. Одним из них является TPK 3 , калиевый канал , который активируется Ca. 2+ и проводит К + из просвета тилакоида в строму , которая помогает установить электрическое поле . С другой стороны, электронейтральный K + оттока антипортер (KEA 3 ) транспортирует K + в просвет тилакоида и H + в строму, что помогает установить градиент pH . [7]
Ионные градиенты
[ редактировать ]
Поскольку ионы заряжены, они не могут проходить через клеточные мембраны путем простой диффузии. Переносить ионы через мембрану могут два разных механизма: активный и пассивный транспорт. [ нужна ссылка ]
Примером активного транспорта ионов является Na + -К + -АТФаза (НКА). NKA питается за счет гидролиза АТФ до АДФ и неорганического фосфата; на каждую гидролизованную молекулу АТФ приходится три Na + выносятся наружу и два K + транспортируются внутри клетки. Это делает внутреннюю часть клетки более отрицательной, чем внешняя, и, более конкретно, создает мембранный потенциал V мембраны около -60 мВ . [5]
Примером пассивного транспорта являются потоки ионов через Na. + , К + , Как 2+ и Cl − каналы. В отличие от активного транспорта, пассивный транспорт осуществляется за счет суммы осмоса арифметической (градиент концентрации) и электрического поля (трансмембранный потенциал). Формально изменение молярной свободной энергии Гиббса , связанное с успешным транспортом, равно [ нужна ссылка ] где R представляет собой газовую постоянную , T представляет собой абсолютную температуру , z — заряд иона, а F представляет собой постоянную Фарадея . [2] : 464–465

На примере На + , оба члена имеют тенденцию поддерживать транспорт: отрицательный электрический потенциал внутри клетки притягивает положительный ион, и поскольку Na + концентрируется вне клетки, осмос обеспечивает диффузию через Na + канал в клетку. В случае с К. + , эффект осмоса обратный: хотя внешние ионы притягиваются отрицательным внутриклеточным потенциалом, энтропия стремится рассеять ионы, уже сконцентрированные внутри клетки. Обратное явление (осмос поддерживает транспорт, электрический потенциал противодействует) может быть достигнуто для Na + в клетках с аномальными трансмембранными потенциалами: при +70 мВ Na + приток прекращается; при более высоких потенциалах он становится истечением. [ нужна ссылка ]
| Ион | млекопитающее | Аксон кальмара | С. cerevisiae | кишечная палочка | Морская вода | ||
|---|---|---|---|---|---|---|---|
| Клетка | Кровь | Клетка | Кровь | ||||
| К + | 100 - 140 | 4-5 | 400 | 10 - 20 | 300 | 30 - 300 | 10 |
| Уже + | 5-15 | 145 | 50 | 440 | 30 | 10 | 500 |
| мг 2+ | 10 [а] 0.5 - 0.8 [б] | 1 - 1.5 | 50 | 30 - 100 [а] 0.01 - 1 [б] | 50 | ||
| Что 2+ | 10 −4 | 2.2 - 2.6 [с] 1.3 - 1.5 [д] | 10 −4 - 3×10 −4 | 10 | 2 | 3 [а] 10 −4 [б] | 10 |
| кл. − | 4 | 110 | 40 - 150 | 560 | 10 - 200 [и] | 500 | |
| Х − (отрицательно заряженные белки) | 138 | 9 | 300 - 400 | 5-10 | |||
| HCOHCO3 − | 12 | 29 | |||||
| рН | 7.1 - 7.3 [12] | с 7.35 до 7.45 [12] (нормальный pH артериальной крови) 6.9 - 7.8 [12] (общий диапазон) | 7.2 - 7.8 [13] | 8.1 - 8.2 [14] | |||
- ^ Jump up to: а б с Граница
- ^ Jump up to: а б с Бесплатно
- ^ Итого
- ^ Ионизированный
- ^ Среднезависимый
Протонные градиенты
[ редактировать ]Протонные градиенты особенно важны во многих типах клеток как форма хранения энергии. Градиент обычно используется для управления АТФ-синтазой, вращением жгутиков или транспортом метаболитов . [15] В этом разделе основное внимание будет уделено трем процессам, которые помогают устанавливать протонные градиенты в соответствующих клетках: бактериородопсину , нециклическому фотофосфорилированию и окислительному фосфорилированию. [ нужна ссылка ]
Бактериородопсин
[ редактировать ]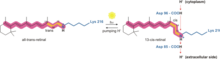
посредством Бактериородопсин генерирует протонный градиент у архей протонного насоса . Протонный насос использует переносчики протонов для перемещения протонов со стороны мембраны с низким H. + концентрации в сторону мембраны с высоким H + концентрация. В бактериородопсине протонный насос активируется за счет поглощения фотонов с длиной волны 568 нм , что приводит к изомеризации основания Шиффа (SB) в сетчатке с образованием К-состояния. Это отодвигает SB от Asp85 и Asp212, вызывая H + переход от SB к Asp85 с образованием состояния M1. Затем белок переходит в состояние М2, отделяя Glu204 от Glu194, что высвобождает протон Glu204 во внешнюю среду. SB репротонируется Asp96, который образует состояние N. Важно, что второй протон происходит от Asp96, поскольку его депротонированное состояние нестабильно и быстро репротонируется протоном из цитозоля . Протонирование Asp85 и Asp96 вызывает реизомеризацию SB с образованием состояния O. Наконец, бактериородопсин возвращается в состояние покоя, когда Asp85 отдает свой протон Glu204. [15] [16]
Фотофосфорилирование
[ редактировать ]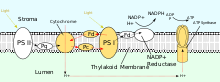
PSII также полагается на свет , чтобы стимулировать образование протонных градиентов в хлоропластах, однако для достижения этой цели PSII использует векторную окислительно-восстановительную химию. Вместо физической транспортировки протонов через белок реакции, требующие связывания протонов, будут происходить на внеклеточной стороне, тогда как реакции, требующие высвобождения протонов, будут происходить на внутриклеточной стороне. Поглощение фотонов с длиной волны 680 нм используется для возбуждения двух электронов в P 680 на более высокий энергетический уровень . Эти электроны с более высокой энергией передаются на связанный с белком пластохинон (PQ A ), а затем на несвязанный пластохинон (PQ B ). Это восстанавливает пластохинон (PQ) до пластохинола (PQH 2 ), который высвобождается из PSII после получения двух протонов из стромы. Электроны в P 680 пополняются за счет окисления воды через комплекс, выделяющий кислород (OEC). Это приводит к высвобождению O 2 и H. + в просвет, для полной реакции [15]

После высвобождения из PSII PQH 2 перемещается к комплексу цитохрома b 6 f , который затем переносит два электрона от PQH 2 к пластоцианину в двух отдельных реакциях. Происходящий процесс аналогичен Q-циклу в Комплексе III цепи переноса электронов. В первой реакции PQH 2 связывается с комплексом на стороне просвета, и один электрон переносится в железо-серный центр , который затем передает его цитохрому f , который затем передает его пластоцианину. Второй электрон переносится на гем b L , который затем передает его на гем b H , который затем передает его на PQ. Во второй реакции второй PQH 2 окисляется, присоединяя электрон к другому пластоцианину и PQ. Обе реакции вместе переносят четыре протона в просвет. [2] : 782–783 [17]
Окислительное фосфорилирование
[ редактировать ]В цепи переноса электронов комплекс I (CI) катализирует восстановление убихинона ( UQ ) до убихинола (UQH 2 ) путем переноса двух электронов от восстановленного никотинамидадениндинуклеотида (НАДН), который перемещает четыре протона из митохондриального матрикса в IMS. : [18]

Комплекс III (CIII) катализирует Q-цикл . Первый этап включает перенос двух электронов от UQH 2, восстановленного CI, к двум молекулам окисленного цитохрома с в месте Q o . На втором этапе еще два электрона восстанавливают UQ до UQH 2 в позиции Q i . Суммарная реакция: [18]

Комплекс IV (CIV) катализирует перенос двух электронов от цитохрома с, восстановленного CIII до половины полного кислорода. Использование одного полного кислорода при окислительном фосфорилировании требует переноса четырех электронов. Затем кислород поглотит четыре протона из матрицы с образованием воды, в то время как еще четыре протона закачиваются в IMS, чтобы дать полную реакцию. [18]
См. также
[ редактировать ]
Ссылки
[ редактировать ]- ^ Нат, Сунил; Вилладсен, Джон (01 марта 2015 г.). «Возвращение к окислительному фосфорилированию». Биотехнология и биоинженерия . 112 (3): 429–437. дои : 10.1002/бит.25492 . ISSN 1097-0290 . ПМИД 25384602 . S2CID 2598635 .
- ^ Jump up to: а б с д и ж Нельсон, Дэвид; Кокс, Майкл (2013). Ленингерские принципы биохимии . Нью-Йорк: WH Freeman. ISBN 978-1-4292-3414-6 .
- ^ Ян, Хуанхэ; Чжан, Гохуэй; Цуй, Цзяньминь (01 января 2015 г.). «Каналы БК: несколько датчиков, один ворота активации» . Границы в физиологии . 6:29 . дои : 10.3389/fphys.2015.00029 . ПМЦ 4319557 . ПМИД 25705194 .
- ^ Шатток, Майкл Дж.; Оттолия, Микела; Берс, Дональд М.; Блауштайн, Мордехай П.; Богуславский Андрей; Боссайт, Джули; Бридж, Джон Х.Б.; Чен-Идзу, Йе; Клэнси, Коллин Э. (15 марта 2015 г.). «Обмен Na+/Ca2+ и Na+/K+-АТФаза в сердце» . Журнал физиологии . 593 (6): 1361–1382. дои : 10.1113/jphysicalol.2014.282319 . ISSN 1469-7793 . ПМК 4376416 . ПМИД 25772291 .
- ^ Jump up to: а б Аперия, Анита; Аккуратов Евгений Евгеньевич; Фонтана, Якопо Мария; Брисмар, Ялмар (01 апреля 2016 г.). «Na+-K+-АТФаза, новый класс рецепторов плазматической мембраны» . Американский журнал физиологии. Клеточная физиология . 310 (7): C491–C495. doi : 10.1152/ajpcell.00359.2015 . ISSN 0363-6143 . ПМИД 26791490 .
- ^ Побурко, Дэймон; Деморекс, Николя (24 апреля 2012 г.). «Регуляция митохондриального протонного градиента цитозольными сигналами Ca2+» (PDF) . Архив Pflügers: Европейский журнал физиологии . 464 (1): 19–26. дои : 10.1007/s00424-012-1106-y . ISSN 0031-6768 . ПМИД 22526460 . S2CID 18133149 .
- ^ Хёнер, Рикарда; Абукила, Али; Кунц, Ханс-Хеннинг; Венема, Кес (01 января 2016 г.). «Протонные градиенты и протонзависимые процессы транспорта в хлоропластах» . Границы в науке о растениях . 7 : 218. doi : 10.3389/fpls.2016.00218 . ПМК 4770017 . ПМИД 26973667 .
- ^ Филипс, Рон Майло и Рон. «»Какова концентрация различных ионов в клетках?» . Проверено 7 июня 2019 г.
- ^ Лодиш, Харви; Берк, Арнольд; Зипурски, С. Лоуренс; Мацудайра, Пол; Балтимор, Дэвид; Дарнелл, Джеймс (2000). «Таблица 15-1. Типичные концентрации ионов у беспозвоночных и позвоночных» . www.ncbi.nlm.nih.gov . Проверено 7 июня 2019 г.
- ^ «Следующая таблица дает представление о внутри- и внеклеточной концентрации ионов в аксоне кальмара и клетке млекопитающего» . www.chm.bris.ac.uk. Проверено 7 июня 2019 г.
- ^ Дием К., Лентер К. Научные таблицы . Том. 565 (Седьмое изд.). Базель: Ciba-Geigy Limited. стр. 653–654. ISBN 978-3-9801244-0-9 .
- ^ Jump up to: а б с Спитцер, Кеннет В.; Воан-Джонс, Ричард Д. (2003), Кармазин, Моррис; Авкиран, Метин; Флигель, Ларри (ред.), «Регуляция внутриклеточного pH в клетках млекопитающих», Натрий-водородный обменник: от молекулы к его роли в заболевании , Springer US, стр. 1–15, doi : 10.1007/978-1-4615 -0427-6_1 , ISBN 9781461504276
- ^ Слончевски, Джоан Л.; Уилкс, Джессика К. (1 августа 2007 г.). «РН цитоплазмы и периплазмы Escherichia coli: быстрое измерение с помощью флуориметрии зеленых флуоресцентных белков» . Журнал бактериологии . 189 (15): 5601–5607. дои : 10.1128/JB.00615-07 . ISSN 0021-9193 . ЧВК 1951819 . ПМИД 17545292 .
- ^ Брюэр, Питер Г. (1 сентября 2008 г.). «Повышение кислотности океана: еще одна проблема CO2» . doi : 10.1038/scientificamericanearth0908-22 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Jump up to: а б с Ганнер, MR; Амин, Мухамед; Чжу, Суюй; Лу, Цзяньсюнь (01 августа 2013 г.). «Молекулярные механизмы генерации трансмембранных протонных градиентов» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . Металлы в биоэнергетике и биомиметических системах. 1827 (8–9): 892–913. дои : 10.1016/j.bbabio.2013.03.001 . ПМЦ 3714358 . ПМИД 23507617 .
- ^ Викстранд, Сесилия; Додс, Роберт; Руайан, Антуан; Нойце, Ричард (01 марта 2015 г.). «Бактериородопсин: не могли бы настоящие структурные промежуточные соединения встать?» . Biochimica et Biophysical Acta (BBA) – Общие предметы . Структурная биохимия и биофизика мембранных белков. 1850 (3): 536–553. дои : 10.1016/j.bbagen.2014.05.021 . ПМИД 24918316 .
- ^ Шеттлер, Марк Аурел; Тот, Сильвия З.; Булуи, Аликс; Калау, Сабина (01 мая 2015 г.). «Динамика стехиометрии фотосинтетического комплекса у высших растений: биогенез, функция и оборот АТФ-синтазы и комплекса цитохрома b 6 f» . Журнал экспериментальной ботаники . 66 (9): 2373–2400. дои : 10.1093/jxb/eru495 . ISSN 0022-0957 . ПМИД 25540437 .
- ^ Jump up to: а б с Сунь, Фэй; Чжоу, Цянцзюнь; Пан, Сяоюнь; Сюй, Инчжи; Рао, Зихе (01 августа 2013 г.). «Выявление различных взаимосвязей переноса электронов и перекачки протонов в дыхательной цепи митохондрий». Современное мнение в области структурной биологии . 23 (4): 526–538. дои : 10.1016/j.sbi.2013.06.013 . ПМИД 23867107 .
- Кэмпбелл и Рис (2005). Биология . Пирсон Бенджамин Каммингс. ISBN 978-0-8053-7146-8 .
- Стивен Т. Абедон, «Важные слова и концепции из главы 8, Campbell & Reece, 2002 (14 января 2005 г.)», для биологии 113 в Университете штата Огайо.