Комплекс цитохрома b 6 f
| Цитохромный комплекс b6f | |
|---|---|
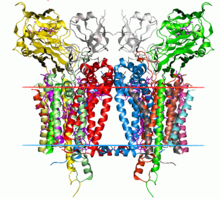 Кристаллическая структура комплекса цитохрома b6f C. Reinhardtii ( 1q90 ). Углеводородные границы липидного бислоя показаны красными и синими линиями (со стороны тилакоидного пространства и со стороны стромы соответственно). | |
| Идентификаторы | |
| Символ | Б6Ф |
| TCDB | 3.D.3 |
| Суперсемейство OPM | 92 |
| белок OPM | 4pv1 |
| Мембраном | 258 |
| Комплекс цитохрома b 6 f | |||
|---|---|---|---|
| Идентификаторы | |||
| Номер ЕС. | 7.1.1.6 | ||
| Номер CAS. | 79079-13-3 | ||
| Альт. имена | Пластохинол/пластоцианинредуктаза | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| |||
цитохрома ( пластохинол b 6 f Комплекс /пластоцианинредуктаза или пластохинол/пластоцианиноксидоредуктаза; EC 7.1.1.6 ) представляет собой фермент, обнаруженный в тилакоидной мембране хлоропластов растений, цианобактерий и зеленых водорослей , который катализирует перенос электронов от пластохинола к пластоцианин :
- пластохинол + 2 окисленный пластоцианин + 2 H + [сторона 1] пластохинон + 2 восстановленный пластоцианин + 4 H + [сторона 2]. [1]

Реакция аналогична реакции, катализируемой цитохромом bc 1 (Комплекс III) митохондрий электрон-транспортной цепи . В ходе фотосинтеза комплекс цитохрома b 6 f является одной из ступеней цепи, переносящей электроны из Фотосистемы II в Фотосистему I , и одновременно перекачивает протоны в пространство тилакоидов, способствуя образованию электрохимического (энергетического) градиента. [2] который позже используется для синтеза АТФ из АДФ .
Структура фермента
[ редактировать ]Комплекс цитохрома b 6 f представляет собой димер, каждый мономер которого состоит из восьми субъединиц. [3] Они состоят из четырех крупных субъединиц: цитохрома f массой 32 кДа с цитохромом c-типа, цитохрома b6 массой 25 кДа с низко- и высокопотенциальной гемовой группой, железо-серного белка Риске массой 19 кДа, содержащего [2Fe-2S ] кластер и субъединицу IV массой 17 кДа; вместе с четырьмя небольшими субъединицами (3-4 кДа): PetG, PetL, PetM и PetN. [3] [4] Общая молекулярная масса составляет 217 кДа.
Кристаллические структуры комплексов цитохрома b 6 f Chlamydomonas reinhardtii , Mastigocladus laminosus и Nostoc sp. PCC 7120 определены. [2] [5] [6] [7] [8] [9]
Ядро комплекса структурно сходно с ядром цитохрома bc1 . Цитохром b 6 и субъединица IV гомологичны цитохрому b , [10] и железо-серные белки Риске двух комплексов гомологичны. [11] Однако цитохром f и цитохром с 1 не гомологичны. [12]
Цитохром b 6 f содержит семь простетических групп . [13] [14] Четыре из них обнаружены в обоих цитохромах b 6 f и bc 1 : гем c-типа цитохромов c 1 и f, два гема b-типа (b p и bn ) в bc 1 и b 6 f и [2Fe -2S] кластер белка Риске. обнаружены три уникальные простетические группы В цитохроме b6f : хлорофилл a , β-каротин и гем cn (также известный как гем x). [5]
Межмономерное пространство внутри ядра димера комплекса цитохрома b6f занято липидами, [9] который обеспечивает направленность переноса электронов гем-гем за счет модуляции внутрибелковой диэлектрической среды. [15]
| ||||||||||||||
Биологическая функция
[ редактировать ]
При фотосинтезе комплекс цитохрома b 6 f обеспечивает перенос электронов и энергии между двумя комплексами фотосинтетических реакционных центров, Фотосистемой II и Фотосистемой I , одновременно перенося протоны из стромы хлоропласта через мембрану тилакоида в просвет . [2] Транспорт электронов через цитохром b 6 f отвечает за создание протонного градиента , который управляет синтезом АТФ в хлоропластах. [4]
В отдельной реакции комплекс цитохрома b 6 f играет центральную роль в циклическом фотофосфорилировании , когда НАДФ + не способен принимать электроны от восстановленного ферредоксина . [16] Этот цикл, движимый энергией P700 + , способствует созданию протонного градиента, который можно использовать для управления синтезом АТФ. Было показано, что этот цикл важен для фотосинтеза. [17] помогает поддерживать правильное соотношение производства АТФ/НАДФН для фиксации углерода . [18] [19]
Реакции депротонирования-окисления p-сторонних хинолов в комплексе цитохрома b6f участвуют в образовании активных форм кислорода. [20] Было высказано предположение, что интегральная молекула хлорофилла, расположенная в месте окисления хинола, выполняет структурную, нефотохимическую функцию, повышая скорость образования активных форм кислорода, возможно, обеспечивая окислительно-восстановительный путь для внутриклеточной коммуникации. [21]
Механизм реакции
[ редактировать ]Комплекс цитохрома b 6 f отвечает за « нециклический » (1) и « циклический » (2) перенос электронов между двумя мобильными окислительно-восстановительными переносчиками, пластохинолом (QH 2 ) и пластоцианином (Pc):
| Н 2 О | → | фотосистема II | → | Национальное собрание 2 | → | Цит б 6 ф | → | ПК | → | фотосистема I | → | НАДФН | (1) |
| Национальное собрание 2 | → | Цит б 6 ф | → | ПК | → | фотосистема I | → | вопрос | (2) | ||||
Цитохром b 6 f катализирует перенос электронов от пластохинола к пластоцианину, перекачивая при этом два протона из стромы в просвет тилакоида:
- БЧ 2 + 2шт(Cu 2+ ) + 2Н + (строма) → Q + 2Pc(Cu + ) + 4H + (люмен) [16]
Эта реакция происходит через цикл Q, как и в Комплексе III. [22] Пластохинол действует как переносчик электронов, передавая два своих электрона в цепи переноса электронов (ETC) с высоким и низким потенциалом посредством механизма, называемого бифуркацией электронов . [23] Комплекс содержит до трех молекул пластохинона, которые образуют сеть переноса электронов, отвечающую за работу Q-цикла и его окислительно-восстановительную и каталитическую функции при фотосинтезе. [24]
Q-цикл
[ редактировать ]
Первая половина цикла Q
- QH 2 связывается с положительной стороной «p» (стороной просвета) комплекса. Он окисляется до семихинона (SQ) железо-серным центром (высокопотенциальный ЭТЦ) и высвобождает два протона в просвет тилакоида. [ нужна ссылка ] .
- Восстановленный железо-серный центр передает свой электрон через цитохром f к Pc.
- В низкопотенциальном ЭТЦ SQ переносит свой электрон на гем b p цитохрома b6.
- Затем гем b p переносит электрон на гем b n .
- Гем b n восстанавливает Q одним электроном с образованием SQ.
Вторая половина цикла Q
- Второй QH 2 связывается с комплексом.
- В высокопотенциальном ЭТЦ один электрон восстанавливает другой окисленный ПК.
- В низкопотенциальном ЭТЦ электрон от гема b n переносится на SQ, а полностью восстановленный Q 2− захватывает два протона стромы с образованием QH 2 .
- Окисленный Q и восстановленный QH 2 диффундируют в мембрану.
Циклический перенос электронов
[ редактировать ]В отличие от Комплекса III, цитохром b 6 f катализирует другую реакцию переноса электрона, которая играет центральную роль в циклическом фотофосфорилировании . Электрон от ферредоксина (Fd) переносится на пластохинон, а затем на комплекс цитохрома b 6 f для восстановления пластоцианина, который повторно окисляется P700 в фотосистеме I. [25] Точный механизм восстановления пластохинона ферредоксином все еще исследуется. Одно из предположений состоит в том, что существует ферредоксин: пластохинонредуктаза или НАДФ-дегидрогеназа. [25] Поскольку гем x, по-видимому, не требуется для цикла Q и не обнаружен в Комплексе III, было предложено использовать его для циклического фотофосфорилирования по следующему механизму: [23] [26]
- Fd (красный) + гем x (ox) → Fd (ox) + гем x (красный)
- гем x (красный) + Fd (красный) + Q + 2H + → гем x (ox) + Fd (ox) + QH 2
Ссылки
[ редактировать ]- ^ ExplorEnz: EC 7.1.1.6
- ^ Перейти обратно: а б с Хасан С.С., Ямашита Э., Баниулис Д., Крамер В.А. (март 2013 г.). «Хинон-зависимые пути переноса протонов в фотосинтетическом комплексе цитохрома b6f» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (11): 4297–302. дои : 10.1073/pnas.1222248110 . ПМК 3600468 . ПМИД 23440205 .
- ^ Перейти обратно: а б Уайтлегг Дж. П., Чжан Х., Агилера Р., Тейлор Р. М., Крамер В. А. (октябрь 2002 г.). «Жидкостная хроматография с полным охватом субъединиц, масс-спектрометрия с ионизацией электрораспылением (LCMS +) олигомерного мембранного белка: комплекс цитохрома b (6) f из шпината и цианобактерии Mastigocladus laminosus» . Молекулярная и клеточная протеомика . 1 (10): 816–27. дои : 10.1074/mcp.m200045-mcp200 . ПМИД 12438564 .
- ^ Перейти обратно: а б Фут-ди-джей, Фут JG (2011). Биохимия . Нью-Йорк, штат Нью-Йорк: Wiley, J. ISBN 978-0-470-57095-1 .
- ^ Перейти обратно: а б Штробель Д., Шоке Ю., Попот Дж.Л., Пико Д. (ноябрь 2003 г.). «Атипичный гем в комплексе цитохрома b (6) f». Природа . 426 (6965): 413–8. дои : 10.1038/nature02155 . ПМИД 14647374 . S2CID 130033 .
- ^ Ямасита Э., Чжан Х., Крамер В.А. (июнь 2007 г.). «Структура комплекса цитохрома b6f: ингибиторы хиноновых аналогов как лиганды гема cn» . Журнал молекулярной биологии . 370 (1): 39–52. дои : 10.1016/j.jmb.2007.04.011 . ЧВК 1993820 . ПМИД 17498743 .
- ^ Баниулис Д., Ямасита Э., Уайтлегг Дж.П., Зацман А.И., Хендрих М.П., Хасан С.С., Райан К.М., Крамер В.А. (апрель 2009 г.). «Структура-функция, стабильность и химическая модификация цианобактериального комплекса цитохрома b6f от Nostoc sp. PCC 7120» . Журнал биологической химии . 284 (15): 9861–9. дои : 10.1074/jbc.M809196200 . ПМК 2665108 . ПМИД 19189962 .
- ^ Хасан С.С., Стофлет Дж.Т., Ямашита Э., Крамер В.А. (апрель 2013 г.). «Липид-индуцированные конформационные изменения в комплексе цитохрома b6f оксигенного фотосинтеза» . Биохимия . 52 (15): 2649–54. дои : 10.1021/bi301638h . ПМК 4034689 . ПМИД 23514009 .
- ^ Перейти обратно: а б Хасан С.С., Крамер В.А. (июль 2014 г.). «Внутренняя липидная архитектура гетероолигомерного комплекса цитохрома b6f» . Структура . 22 (7): 1008–15. doi : 10.1016/j.str.2014.05.004 . ПМК 4105968 . ПМИД 24931468 .
- ^ Видгер В.Р., Крамер В.А., Херрманн Р.Г., Требст А. (февраль 1984 г.). «Гомология последовательностей и структурное сходство между цитохромом b митохондриального комплекса III и хлоропластным комплексом b6-f: положение гемов цитохрома b в мембране» . Труды Национальной академии наук Соединенных Штатов Америки . 81 (3): 674–8. дои : 10.1073/pnas.81.3.674 . ПМЦ 344897 . ПМИД 6322162 .
- ^ Каррелл С.Дж., Чжан Х., Крамер В.А., Смит Дж.Л. (декабрь 1997 г.). «Биологическая идентичность и разнообразие фотосинтеза и дыхания: структура просветного домена хлоропластного белка Риске» . Структура . 5 (12): 1613–25. дои : 10.1016/s0969-2126(97)00309-2 . ПМИД 9438861 .
- ^ Мартинес С.Е., Хуанг Д., Щепаньяк А., Крамер В.А., Смит Дж.Л. (февраль 1994 г.). «Кристаллическая структура цитохрома f хлоропласта обнаруживает новую складку цитохрома и неожиданное лигирование гема» . Структура . 2 (2): 95–105. дои : 10.1016/s0969-2126(00)00012-5 . ПМИД 8081747 .
- ^ Баниулис Д., Ямашита Э., Чжан Х., Хасан С.С., Крамер В.А. (2008). «Структура-функция комплекса цитохрома b6f». Фотохимия и фотобиология . 84 (6): 1349–58. дои : 10.1111/j.1751-1097.2008.00444.x . ПМИД 19067956 . S2CID 44992397 .
- ^ Крамер В.А., Чжан Х., Ян Дж., Курису Г., Смит Дж.Л. (май 2004 г.). «Эволюция фотосинтеза: независимая от времени структура комплекса цитохрома b6f». Биохимия . 43 (20): 5921–9. дои : 10.1021/bi049444o . ПМИД 15147175 .
- ^ Хасан С.С., Захаров С.Д., Шове А., Стадницкий В., Савихин С., Крамер В.А. (июнь 2014 г.). «Карта диэлектрической гетерогенности мембранного белка: гетероолигомерный комплекс цитохрома b6f» . Журнал физической химии Б. 118 (24): 6614–25. дои : 10.1021/jp501165k . ПМК 4067154 . ПМИД 24867491 .
- ^ Перейти обратно: а б Берг Дж. М., Тимочко Дж. Л., Страйер Л., Страйер Л. (2007). Биохимия . Нью-Йорк: WH Freeman. ISBN 978-0-7167-8724-2 .
- ^ Мунекаге И., Хасимото М., Мияке С., Томидзава К., Эндо Т., Тасака М., Шиканай Т. (июнь 2004 г.). «Циклический поток электронов вокруг фотосистемы I необходим для фотосинтеза». Природа . 429 (6991): 579–82. Бибкод : 2004Natur.429..579M . дои : 10.1038/nature02598 . ПМИД 15175756 . S2CID 4421776 .
- ^ Бланкеншип RE (2002). Молекулярные механизмы фотосинтеза . Оксфорд; Молден, Массачусетс: Blackwell Science. ISBN 978-0-632-04321-7 .
- ^ Бендалл Д. (1995). «Циклическое фотофосфорилирование и транспорт электронов» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1229 : 23–38. дои : 10.1016/0005-2728(94)00195-Б .
- ^ Баниулис Д., Хасан С.С., Стофлет Дж.Т., Крамер В.А. (декабрь 2013 г.). «Механизм усиленного образования супероксида в комплексе цитохрома b(6)f оксигенного фотосинтеза» . Биохимия . 52 (50): 8975–83. дои : 10.1021/bi4013534 . ПМК 4037229 . ПМИД 24298890 .
- ^ Хасан С.С., Проктор Э.А., Ямасита Э., Дохолян Н.В., Крамер В.А. (октябрь 2014 г.). «Трафик внутри липопротеинового комплекса цитохрома b6f: ворота хинонового портала» . Биофизический журнал . 107 (7): 1620–8. Бибкод : 2014BpJ...107.1620H . дои : 10.1016/j.bpj.2014.08.003 . ПМК 4190601 . ПМИД 25296314 .
- ^ Крамер В.А., Сориано Г.М., Пономарев М., Хуан Д., Чжан Х., Мартинес С.Е., Смит Дж.Л. (июнь 1996 г.). «Некоторые новые структурные аспекты и старые споры относительно комплекса цитохрома b6f кислородного фотосинтеза». Ежегодный обзор физиологии растений и молекулярной биологии растений . 47 : 477–508. дои : 10.1146/annurev.arplant.47.1.477 . ПМИД 15012298 .
- ^ Перейти обратно: а б Крамер В.А., Чжан Х., Ян Дж., Курису Г., Смит Дж.Л. (2006). «Трансмембранный трафик в комплексе цитохрома b6f». Ежегодный обзор биохимии . 75 : 769–90. doi : 10.1146/annurev.biochem.75.103004.142756 . ПМИД 16756511 .
- ^ Мэлоун Л.А., Цянь П., Мейнорд Г.Е., Хичкок А., Фармер Д.А., Томпсон Р.Ф. и др. (ноябрь 2019 г.). «Крио-ЭМ-структура комплекса цитохрома B 6 F шпината при разрешении 3,6 Å» (PDF) . Природа . 575 (7783): 535–539. дои : 10.1038/s41586-019-1746-6 . ПМИД 31723268 . S2CID 207987984 .
- ^ Перейти обратно: а б Жолио П., Жолио А. (июль 2002 г.). «Циклический перенос электронов в листе растения» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (15): 10209–14. Бибкод : 2002PNAS...9910209J . дои : 10.1073/pnas.102306999 . ПМК 126649 . ПМИД 12119384 .
- ^ Крамер В.А., Ян Дж., Чжан Х., Курису Г., Смит Дж.Л. (2005). «Структура комплекса цитохрома b6f: новые простетические группы, Q-пространство и гипотеза закуски» для сборки комплекса». Исследования фотосинтеза . 85 (1): 133–43. Бибкод : 2005PhoRe..85..133C . дои : 10.1007/s11120-004-2149-5 . ПМИД 15977064 . S2CID 20731696 .
Дальнейшее чтение
[ редактировать ]- Саревич, М; Пинчер, С; Пьетрас, Р; Борек, А; Буйнович, Л.; Ханке, Г; Крамер, Вашингтон; Финацци, Дж; Осичка, А (24 февраля 2021 г.). «Каталитические реакции и сохранение энергии в цитохромных комплексах bc(1) и b(6)f энергопередающих мембран» . Химические обзоры . 121 (4): 2020–2108. doi : 10.1021/acs.chemrev.0c00712 . ПМК 7908018 . ПМИД 33464892 .
Внешние ссылки
[ редактировать ]- цитохрома b 6 f Структурно-функциональные исследования комплекса - текущие исследования цитохрома b 6 f в лаборатории Уильяма Крамера в Университете Пердью, США
- UMich Ориентация белков в семействах мембран/суперсемействе-3 - Расчетные положения b6f и родственных комплексов в мембранах
- Комплекс Цитохром + b6f + в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Пластохинол-пластоцианин+редуктаза в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)