Тилакоид
| Клеточная биология | |
|---|---|
| хлоропласт | |
 Компоненты типичного хлоропласта
1 Гранум 3 Тилакоида ◄ Вы здесь
4 Стромальный тилакоид |

Тилакоиды представляют собой мембраносвязанные отсеки внутри хлоропластов и цианобактерий . Они являются местом светозависимых фотосинтеза реакций . Тилакоиды состоят из тилакоидной мембраны, окружающей просвет тилакоида . Тилакоиды хлоропластов часто образуют стопки дисков, называемые гранами (единственное число: granum ). Граны соединены межгранальными или стромальными тилакоидами, которые объединяют стопки гранумов в единый функциональный отсек.
В мембранах тилакоидов пигменты хлорофилла находятся в пакетах, называемых квантазомами . Каждая квантасома содержит от 230 до 250 молекул хлорофилла.
Этимология
[ редактировать ]Слово «тилакоид» происходит от греческого слова «thilakos» или θύλακος , что означает «мешочек» или «мешочек». [ 1 ] Таким образом, тилакоид означает «мешкообразный» или «мешочек».
Структура
[ редактировать ]


Тилакоиды представляют собой мембраносвязанные структуры, встроенные в строму хлоропласта . Стопка тилакоидов называется гранумом и напоминает стопку монет.
Мембрана
[ редактировать ]Тилакоидная мембрана является местом светозависимых реакций фотосинтеза, при этом фотосинтетические пигменты встроены непосредственно в мембрану. Это чередующийся узор из темных и светлых полос размером 1 нанометр каждая . [ 3 ] Липидный бислой тилакоида имеет общие характеристики с мембранами прокариот и внутренней мембраной хлоропластов. Например, кислые липиды содержатся в мембранах тилакоидов, цианобактерий и других фотосинтезирующих бактерий и участвуют в функциональной целостности фотосистем. [ 4 ] Тилакоидные мембраны высших растений состоят преимущественно из фосфолипидов. [ 5 ] и галактолипиды , которые асимметрично расположены вдоль и поперек мембран. [ 6 ] Мембраны тилакоидов богаче галактолипидами, чем фосфолипидами; также они преимущественно состоят из гексагональной фазы II, образующей моногалактозилдиглицеридный липид. Было показано, что, несмотря на этот уникальный состав, мембраны тилакоидов растений в основном принимают динамическую организацию липидного бислоя. [ 7 ] Липиды, образующие мембраны тилакоидов, наиболее богаты высокотекучей линоленовой кислотой. [ 8 ] синтезируются сложным путем, включающим обмен липидными предшественниками между эндоплазматическим ретикулумом и внутренней мембраной пластидной оболочки и транспортируются от внутренней мембраны к тилакоидам через везикулы. [ 9 ]
Люмен
[ редактировать ]тилакоида представляет собой сплошную водную фазу , Просвет окруженную тилакоидной мембраной . Он играет важную роль в фотофосфорилировании во время фотосинтеза . Во время светозависимой реакции протоны перекачиваются через тилакоидную мембрану в полость, делая ее кислой до pH 4.
Гранулы и пластинки стромы
[ редактировать ]У высших растений тилакоиды организованы в мембранный комплекс гранум-строма. Гранум грана (множественное число ) представляет собой стопку тилакоидных дисков. Хлоропласты могут иметь от 10 до 100 гран. Граны соединены тилакоидами стромы, также называемыми межгранальными тилакоидами или пластинками . Тилакоиды грана и тилакоиды стромы можно отличить по разному белковому составу. Граны способствуют большому соотношению площади поверхности к объему хлоропластов. Недавнее исследование электронной томографии тилакоидных мембран показало, что ламели стромы организованы в широкие листы, перпендикулярные оси стопки гран, и образуют множественные правосторонние спиральные поверхности на границе раздела гран. [ 2 ] Левовинтовые поверхности консолидируются между правовинтовыми спиралями и листами. Было показано, что эта сложная сеть чередующихся спиральных мембранных поверхностей разного радиуса и шага минимизирует поверхностную энергию и энергию изгиба мембран. [ 2 ] Эта новая модель, самая обширная из созданных на сегодняшний день, показала, что функции двух, казалось бы, противоречивых старых моделей [ 10 ] [ 11 ] сосуществовать в структуре. Примечательно, что подобное расположение спиральных элементов чередующейся направленности, часто называемых структурами «гаража», было предложено для присутствия в эндоплазматическом ретикулуме. [ 12 ] и в сверхплотной ядерной материи. [ 13 ] [ 14 ] [ 15 ] Эта структурная организация может представлять собой фундаментальную геометрию для соединения между плотно упакованными слоями или листами. [ 2 ]
Формирование
[ редактировать ]Хлоропласты развиваются из пропластид , когда всходы выходят из земли. Для формирования тилакоидов необходим свет. В зародыше растения и в отсутствие света пропластиды развиваются в этиопласты , содержащие полукристаллические мембранные структуры, называемые проламеллярными тельцами. Под воздействием света эти проламеллярные тельца превращаются в тилакоиды. Этого не происходит у выращенных в темноте сеянцев, подвергающихся этиоляции . Недостаточное воздействие света может привести к отказу тилакоидов. Это приводит к выходу из строя хлоропластов, что приводит к гибели растения.
Для формирования тилакоидов необходимо действие индуцирующего пузырьки белка в пластидах 1 (VIPP1). Растения не могут выжить без этого белка, а снижение уровня VIPP1 приводит к замедлению роста и бледности растений с пониженной способностью к фотосинтезу. VIPP1, по-видимому, необходим для формирования основной тилакоидной мембраны, но не для сборки белковых комплексов тилакоидной мембраны. [ 16 ] Он консервативен у всех организмов, содержащих тилакоиды, включая цианобактерии, [ 17 ] зеленые водоросли, такие как хламидомонада , [ 18 ] и высшие растения, такие как Arabidopsis thaliana . [ 19 ]
Выделение и фракционирование
[ редактировать ]Тилакоиды можно очистить из растительных клеток, используя комбинацию дифференциального и градиентного центрифугирования . [ 20 ] Разрушение изолированных тилакоидов, например, путем механического сдвига, приводит к высвобождению просветной фракции. Периферийные и интегральные мембранные фракции можно выделить из оставшейся мембранной фракции. Обработка карбонатом натрия (Na 2 CO 3 ) отщепляет периферические мембранные белки , тогда как обработка детергентами и органическими растворителями солюбилизирует интегральные мембранные белки .
Белки
[ редактировать ]
Тилакоиды содержат множество интегральных и периферических мембранных белков, а также просветных белков. Недавние протеомные исследования фракций тилакоидов предоставили дополнительную информацию о белковом составе тилакоидов. [ 21 ] Эти данные были обобщены в нескольких базах данных пластидных белков, доступных в Интернете. [ 22 ] [ 23 ]
Согласно этим исследованиям, протеом тилакоида состоит как минимум из 335 различных белков. Из них 89 находятся в просвете, 116 являются интегральными мембранными белками, 62 являются периферическими белками на стороне стромы и 68 периферическими белками на стороне просвета. Дополнительные белки с низким содержанием просвета можно предсказать с помощью вычислительных методов. [ 20 ] [ 24 ] Из белков тилакоидов с известными функциями в фотосинтезе участвуют 42%. Следующие по величине функциональные группы включают белки, участвующие в нацеливании , процессинге и сворачивании белков (11%), реакции на окислительный стресс (9%) и трансляции (8%). [ 22 ]
Интегральные мембранные белки
[ редактировать ]Мембраны тилакоидов содержат интегральные мембранные белки , которые играют важную роль в светособирании и светозависимых реакциях фотосинтеза. В мембране тилакоида имеются четыре основных белковых комплекса:
Фотосистема II расположена преимущественно в тилакоидах гран, тогда как фотосистема I и АТФ-синтаза преимущественно расположены в тилакоидах стромы и наружных слоях гран. Комплекс цитохрома b6f равномерно распределен по мембранам тилакоидов. Из-за раздельного расположения двух фотосистем в мембранной системе тилакоидов мобильные переносчики электронов необходимы для перемещения электронов между ними. Этими носителями являются пластохинон и пластоцианин. Пластохинон переносит электроны от фотосистемы II к комплексу цитохрома b6f, тогда как пластоцианин переносит электроны от комплекса цитохрома b6f к фотосистеме I.
Вместе эти белки используют энергию света для управления цепями переноса электронов , которые генерируют хемиосмотический потенциал через тилакоидную мембрану и НАДФН , продукт терминальной окислительно-восстановительной реакции. АТФ -синтаза использует хемиосмотический потенциал для образования АТФ во время фотофосфорилирования .
Фотосистемы
[ редактировать ]Эти фотосистемы представляют собой управляемые светом окислительно-восстановительные центры, каждый из которых состоит из антенного комплекса , который использует хлорофиллы и вспомогательные фотосинтетические пигменты, такие как каротиноиды и фикобилипротеины, для сбора света с различными длинами волн. Каждый антенный комплекс содержит от 250 до 400 молекул пигмента, и поглощаемая ими энергия передается посредством резонансной передачи энергии специализированному хлорофиллу а в реакционном центре каждой фотосистемы. Когда любая из двух молекул хлорофилла а в реакционном центре поглощает энергию, электрон возбуждается и передается молекуле-акцептору электронов. Фотосистема I содержит в своем реакционном центре пару молекул хлорофилла А , обозначенную P700 , которые максимально поглощают свет с длиной волны 700 нм. Фотосистема II содержит хлорофилл P680 , который лучше всего поглощает свет с длиной волны 680 нм (обратите внимание, что эти длины волн соответствуют темно-красному цвету — см. видимый спектр ). P — это сокращение от пигмента, а число — это удельный пик поглощения в нанометрах для молекул хлорофилла в каждом реакционном центре. Это зеленый пигмент, присутствующий в растениях, который не виден невооруженным глазом.
Цитохромный комплекс b6f
[ редактировать ]Комплекс цитохрома b6f является частью цепи переноса электронов тилакоида и связывает перенос электронов с перекачкой протонов в просвет тилакоида. Энергетически он расположен между двумя фотосистемами и переносит электроны от фотосистемы II-пластохинон к пластоцианин-фотосистеме I.
АТФ-синтаза
[ редактировать ]Тилакоидная АТФ-синтаза представляет собой CF1FO-АТФ-синтазу, аналогичную митохондриальной АТФазе. Он интегрирован в тилакоидную мембрану, а часть CF1 прикрепляется к строме. Таким образом, синтез АТФ происходит на стромальной стороне тилакоидов, где АТФ необходим для светонезависимых реакций фотосинтеза.
Белки люмена
[ редактировать ]Белок-переносчик электронов пластоцианин присутствует в просвете и переносит электроны от белкового комплекса цитохрома b6f к фотосистеме I. Хотя пластохиноны являются жирорастворимыми и, следовательно, перемещаются внутри тилакоидной мембраны, пластоцианин перемещается через просвет тилакоида.
Просвет тилакоидов также является местом окисления воды комплексом, выделяющим кислород, связанным с люменальной стороной фотосистемы II.
Люменальные белки можно предсказать вычислительно на основе их нацеливающих сигналов. У арабидопсиса из предсказанных люменальных белков, обладающих сигналом Tat , самые крупные группы с известными функциями: 19% участвуют в процессинге белков (протеолиз и сворачивание), 18% - в фотосинтезе, 11% - в метаболизме и 7% - в окислительно-восстановительных носителях и защите. . [ 20 ]
Экспрессия белка
[ редактировать ]Хлоропласты имеют собственный геном , который кодирует ряд тилакоидных белков. Однако в ходе эволюции пластид от их цианобактериальных эндосимбиотических обширный перенос генов из генома хлоропластов в ядро клетки предков произошел . Это приводит к тому, что четыре основных комплекса белков тилакоидов кодируются частично геномом хлоропластов, а частично ядерным геномом. Растения разработали несколько механизмов совместной регуляции экспрессии различных субъединиц, закодированных в двух разных органеллах, чтобы обеспечить правильную стехиометрию и сборку этих белковых комплексов. Например, транскрипция ядерных генов, кодирующих части фотосинтетического аппарата, регулируется светом . Биогенез, стабильность и оборот тилакоидных белковых комплексов регулируются фосфорилированием с помощью редокс-чувствительных киназ в тилакоидных мембранах. [ 25 ] Скорость трансляции белков, кодируемых хлоропластами, контролируется наличием или отсутствием партнеров сборки (контроль эпистазией синтеза). [ 26 ] Этот механизм включает отрицательную обратную связь посредством связывания избыточного белка с 5'-нетранслируемой областью мРНК хлоропласта . [ 27 ] Хлоропластам также необходимо сбалансировать соотношение фотосистем I и II для цепи переноса электронов. Редокс-состояние переносчика электронов пластохинона в тилакоидной мембране напрямую влияет на транскрипцию генов хлоропластов, кодирующих белки реакционных центров фотосистем, противодействуя тем самым дисбалансам в цепи переноса электронов. [ 28 ]
Белок нацеливается на тилакоиды
[ редактировать ]
Тилакоидные белки доставляются к месту назначения с помощью сигнальных пептидов прокариотического типа и секреторных путей внутри хлоропласта. Большинству тилакоидных белков, кодируемых ядерным геномом растения, для правильной локализации необходимы два нацеливающих сигнала: N-концевой пептид, нацеливающий хлоропласты (показан желтым на рисунке), за которым следует пептид, нацеливающий тилакоиды (показан синим цветом). Белки импортируются через транслокон комплексов внешней и внутренней мембраны ( Toc и Tic ). После попадания в хлоропласт первый целевой пептид отщепляется протеазой, обрабатывающей импортированные белки. Это демаскирует второй сигнал нацеливания, и белок экспортируется из стромы в тилакоид на втором этапе нацеливания. Этот второй этап требует действия белковых транслокационных компонентов тилакоидов и является энергозависимым. Белки встраиваются в мембрану по SRP-зависимому пути (1), Tat-зависимому пути (2) или спонтанно через свои трансмембранные домены (на рисунке не показаны). Люменальные белки экспортируются через тилакоидную мембрану в просвет либо по Tat-зависимому пути (2), либо по Sec-зависимому пути (3) и высвобождаются путем расщепления сигнала, направляющего тилакоиды. Различные пути используют разные сигналы и источники энергии. Путь Sec (секреторный) требует АТФ в качестве источника энергии и состоит из SecA, который связывается с импортированным белком, и мембранного комплекса Sec, обеспечивающего транспортировку белка. Белки с двойником Аргининовый мотив в их сигнальном пептиде тилакоида перемещается по пути Tat (двойная транслокация аргинина), который требует мембраносвязанного комплекса Tat и градиента pH в качестве источника энергии. Некоторые другие белки встраиваются в мембрану по пути SRP ( частица распознавания сигнала ). SRP хлоропласта может взаимодействовать со своими белками-мишенями либо посттрансляционно, либо котрансляционно, транспортируя таким образом импортированные белки, а также те, которые транслируются внутри хлоропласта. Путь SRP требует GTP и градиента pH в качестве источников энергии. Некоторые трансмембранные белки могут также спонтанно встраиваться в мембрану со стороны стромы без затрат энергии. [ 29 ]
Функция
[ редактировать ]
Тилакоиды являются местом светозависимых реакций фотосинтеза. К ним относятся управляемое светом окисление воды и выделение кислорода , перекачка протонов через тилакоидные мембраны в сочетании с цепью переноса электронов фотосистем и цитохромного комплекса, а также синтез АТФ АТФ-синтазой с использованием генерируемого протонного градиента.
Фотолиз воды
[ редактировать ]Первым шагом фотосинтеза является восстановление (расщепление) воды под действием света с целью получения электронов для фотосинтетических цепей переноса электронов, а также протонов для создания протонного градиента. Реакция расщепления воды происходит на просветной стороне тилакоидной мембраны и приводится в действие световой энергией, улавливаемой фотосистемами. В результате окисления воды образуются отходы O 2 , жизненно важные для клеточного дыхания . Молекулярный кислород, образующийся в результате реакции, выбрасывается в атмосферу.
Электронно-транспортные цепи
[ редактировать ]Во время фотосинтеза используются два различных варианта транспорта электронов:
- Нециклический транспорт электронов или нециклическое фотофосфорилирование производит НАДФН + H. + и АТФ.
- Циклический транспорт электронов или циклическое фотофосфорилирование производит только АТФ.
Нециклическая разновидность предполагает участие обеих фотосистем, тогда как циклический поток электронов зависит только от фотосистемы I.
- Фотосистема I использует энергию света для снижения НАДФ. + в НАДФН + Н + и активен как в нециклическом, так и в циклическом транспорте электронов. В циклическом режиме заряженный электрон передается по цепи, которая в конечном итоге возвращает его (в базовом состоянии) к хлорофиллу, который его возбудил.
- Фотосистема II использует энергию света для окисления молекул воды, производя электроны (е − ), протоны (H + ) и молекулярный кислород (O 2 ) и активен только в нециклическом транспорте. Электроны в этой системе не сохраняются, а постоянно поступают из окисленного 2H 2 O (O 2 + 4 H + + 4 и − ) и выход с НАДП + когда он окончательно восстанавливается до НАДФН.
хемиосмос
[ редактировать ]Основной функцией тилакоидной мембраны и ее целостных фотосистем является создание хемиосмотического потенциала. Переносчики в цепи переноса электронов используют часть энергии электронов для активной транспортировки протонов из стромы в просвет . Во время фотосинтеза просвет становится кислым , уровень pH достигает 4 по сравнению с pH 8 в строме. [ 30 ] Это представляет собой 10000-кратный градиент концентрации протонов через тилакоидную мембрану.
Источник протонного градиента
[ редактировать ]Протоны в просвете происходят из трех основных источников.
- Фотолиз фотосистемой II окисляет воду до кислорода , протонов и электронов в просвете.
- Перенос электронов от фотосистемы II к пластохинону при нециклическом транспорте электронов потребляет два протона из стромы. Они высвобождаются в просвет, когда восстановленный пластохинол окисляется белковым комплексом цитохрома b6f на просветной стороне тилакоидной мембраны. Из пула пластохинона электроны проходят через комплекс цитохрома b6f. Эта целостная мембранная сборка напоминает цитохром bc1.
- Восстановление пластохинона ферредоксином . во время циклического транспорта электронов также переносит два протона из стромы в просвет
Протонный градиент также вызван потреблением протонов в строме для образования НАДФН из НАДФ+ на НАДФ-редуктазе.
Генерация АТФ
[ редактировать ]Молекулярный механизм образования АТФ (аденозинтрифосфата) в хлоропластах аналогичен таковому в митохондриях и забирает необходимую энергию за счет движущей силы протонов (ПМФ). [ нужна ссылка ] Однако хлоропласты больше полагаются на химический потенциал ПМФ для выработки потенциальной энергии, необходимой для синтеза АТФ. PMF представляет собой сумму химического потенциала протона (задаваемого градиентом концентрации протонов) и трансмембранного электрического потенциала (задаваемого разделением зарядов на мембране). По сравнению с внутренними мембранами митохондрий, которые имеют значительно более высокий мембранный потенциал из-за разделения зарядов, тилакоидные мембраны лишены градиента заряда. [ нужна ссылка ] Чтобы компенсировать это, 10 000-кратный градиент концентрации протонов на тилакоидной мембране намного выше по сравнению с 10-кратным градиентом на внутренней мембране митохондрий. Результирующий хемиосмотический потенциал между просветом и стромой достаточно высок, чтобы стимулировать синтез АТФ с использованием АТФ-синтазы . Когда протоны движутся обратно по градиенту через каналы АТФ-синтазы , АДФ + P i объединяются в АТФ. Таким образом, светозависимые реакции связаны с синтезом АТФ через протонный градиент. [ нужна ссылка ]
Тилакоидные мембраны цианобактерий.
[ редактировать ]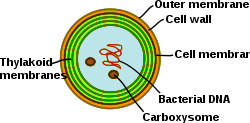
Цианобактерии — фотосинтезирующие прокариоты с высокодифференцированной мембранной системой. Цианобактерии имеют внутреннюю систему тилакоидных мембран, в которой находятся полнофункциональные цепи переноса электронов фотосинтеза и дыхания . Наличие различных мембранных систем придает этим клеткам уникальную сложность среди бактерий . Цианобактерии должны быть способны реорганизовать мембраны, синтезировать новые мембранные липиды и правильно направлять белки в правильную мембранную систему. , Наружная мембрана плазматическая мембрана и тилакоидные мембраны выполняют специализированные функции в цианобактериальной клетке. Понимание организации, функциональности, белкового состава и динамики мембранных систем остается серьезной проблемой в биологии цианобактерий. [ 31 ]
В отличие от сети тилакоидов высших растений, которая дифференцирована на пластинки граны и стромы, тилакоиды у цианобактерий организованы в многочисленные концентрические оболочки, которые расщепляются и сливаются в параллельные слои, образуя высокосвязанную сеть. В результате образуется непрерывная сеть, которая окружает один просвет (как в хлоропластах высших растений) и позволяет водорастворимым и жирорастворимым молекулам диффундировать через всю мембранную сеть. Кроме того, внутри параллельных листов тилакоидов часто наблюдаются перфорации. Эти промежутки в мембране обеспечивают перемещение частиц разного размера по клетке, включая рибосомы, гранулы гликогена и липидные тельца. [ 32 ] Относительно большое расстояние между тилакоидами обеспечивает пространство для внешних светособирающих антенн — фикобилисом . [ 33 ] Эта макроструктура, как и у высших растений, проявляет некоторую гибкость при изменении физико-химической среды. [ 34 ]
См. также
[ редактировать ]- Артур Мейер (ботаник)
- Андре Ягендорф
- хемиосмос
- Электрохимический градиент
- Эндосимбиоз
- Выделение кислорода
- Фотосинтез
Ссылки
[ редактировать ]- ^ θύλακος . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей»
- ^ Jump up to: а б с д и Бусси Ю, Шимони Э, Вайнер А, Капон Р, Чаруви Д, Нево Р, Эфрати Э, Райх З (2019). «Фундаментальная спиральная геометрия укрепляет фотосинтетическую мембрану растения» . Proc Natl Acad Sci США . 116 (44): 22366–22375. Бибкод : 2019PNAS..11622366B . дои : 10.1073/pnas.1905994116 . ПМК 6825288 . ПМИД 31611387 .
- ^ «Фотосинтез» Энциклопедия науки и технологий МакГроу Хилла, 10-е изд. 2007. Том. 13 с. 469
- ^ Сато Н (2004). «Роль кислых липидов сульфохиновозилдиацилглицерина и фосфатидилглицерина в фотосинтезе: их специфичность и эволюция». J Plant Res . 117 (6): 495–505. дои : 10.1007/s10265-004-0183-1 . ПМИД 15538651 . S2CID 27225926 .
- ^ «фотосинтез». Британская энциклопедия. 2008. Британская энциклопедия 2006, Ultimate Reference Suite, DVD, 9 апреля 2008 г.
- ^ Спрак С.Г. (1987). «Структурная и функциональная организация галактолипидов на мембранной организации тилакоидов». J Bioenerg Biomembr . 19 (6): 691–703. дои : 10.1007/BF00762303 . ПМИД 3320041 . S2CID 6076741 .
- ^ ЯшРой, RC (1990). «Магнитно-резонансные исследования динамической организации липидов в мембранах хлоропластов» (PDF) . Журнал биологических наук . 15 (4): 281–288. дои : 10.1007/bf02702669 . S2CID 360223 .
- ^ ЯшРой, RC (1987). «Исследование ЯМР 13С липидных жирно-ацильных цепей мембран хлоропластов» . Индийский журнал биохимии и биофизики . 24 (3): 177–178. ПМИД 3428918 .
- ^ Беннинг С., Сюй С., Авай К. (2006). «Невезикулярный и везикулярный транспорт липидов с участием пластид». Карр Опин Растительная Биол . 9 (3): 241–7. дои : 10.1016/j.pbi.2006.03.012 . ПМИД 16603410 .
- ^ Шимони Э, Рав-Хон О, Охад И, Брумфельд В, Райх З (2005). «Трехмерная организация тилакоидных мембран хлоропластов высших растений, выявленная с помощью электронной томографии» . Растительная клетка . 17 (9): 2580–6. дои : 10.1105/tpc.105.035030 . ПМЦ 1197436 . ПМИД 16055630 .
- ^ Мустарди, Л.; Баттл, К.; Штайнбах, Г.; Гараб, Г. (2008). «Трехмерная сеть тилакоидных мембран у растений: квазиспиральная модель сборки гранум-строма» . Растительная клетка . 20 (10): 2552–2557. дои : 10.1105/tpc.108.059147 . ПМК 2590735 . ПМИД 18952780 .
- ^ Терасаки М, Шемеш Т, Кастури Н, Клемм Р, Шалек Р, Хейворт К, Хэнд А, Янкова М, Хубер Г, Лихтман Дж, Рапопорт Т, Козлов М (2013). «Сложенные друг на друга листы эндоплазматического ретикулума соединены спиральными мембранными мотивами» . Клетка . 154 (2): 285–96. дои : 10.1016/j.cell.2013.06.031 . ПМЦ 3767119 . ПМИД 23870120 .
- ^ Берри Д.К.; Каплан М.Э.; Горовиц CJ; Хубер Дж; Шнайдер А.С. (2016). « Структуры «парковки» в ядерной астрофизике и клеточной биофизике» . Физический обзор C . 94 (5). Американское физическое общество: 055801. arXiv : 1509.00410 . Бибкод : 2016PhRvC..94e5801B . doi : 10.1103/PhysRevC.94.055801 . S2CID 36462725 .
- ^ Горовиц CJ; Берри Д.К.; Бриггс CM; Каплан М.Э.; Камминг А; Шнайдер А.С. (2015). «Неупорядоченная ядерная паста, распад магнитного поля и охлаждение коры нейтронных звезд». Преподобный Летт по физике . 114 (3): 031102. arXiv : 1410.2197 . Бибкод : 2015PhRvL.114c1102H . doi : 10.1103/PhysRevLett.114.031102 . ПМИД 25658989 . S2CID 12021024 .
- ^ Шнайдер АС; Берри Д.К.; Каплан М.Э.; Горовиц CJ; Лин З (2016). «Влияние топологических дефектов на наблюдаемые «ядерной пасты». Физический обзор C . 93 (6): 065806. arXiv : 1602.03215 . Бибкод : 2016PhRvC..93f5806S . дои : 10.1103/PhysRevC.93.065806 . S2CID 28272522 .
- ^ Елена Асеева; Фридрих Оссенбюль; Клаудия Сиппель; Вон К. Чо; Бернхард Штайн; Лутц А. Эйхакер; Йорг Мёрер; Герхард Ваннер; Питер Вестхофф; Юрген Золь; Уте К. Воткнехт (2007). «Vipp1 необходим для формирования основных тилакоидных мембран, но не для сборки тилакоидных белковых комплексов». Растительная Физиол Биохимия . 45 (2): 119–28. дои : 10.1016/j.plaphy.2007.01.005 . ПМИД 17346982 .
- ^ Вестфаль С., Хайнс Л., Солл Дж., Воткнехт У. (2001). «Мутант с делецией Vipp1 Synechocystis: связь между бактериальным фаговым шоком и биогенезом тилакоидов?» . Proc Natl Acad Sci США . 98 (7): 4243–8. дои : 10.1073/pnas.061501198 . ПМК 31210 . ПМИД 11274448 .
- ^ Лю С., Уиллмунд Ф., Голецки Дж., Какаче С., Маркерт С., Хесс Б., Шрода М., Шрода М. (2007). «Хлоропластные шапероны HSP70B-CDJ2-CGE1 катализируют сборку и разборку олигомеров VIPP1 у хламидомонады». Плант Дж . 50 (2): 265–77. дои : 10.1111/j.1365-313X.2007.03047.x . ПМИД 17355436 .
- ^ Кролл Д., Мейерхофф К., Бехтольд Н., Киношита М., Вестфаль С., Воткнехт У., Солл Дж., Вестхофф П. (2001). «VIPP1, ядерный ген Arabidopsis thaliana, необходимый для формирования тилакоидной мембраны» . Proc Natl Acad Sci США . 98 (7): 4238–42. дои : 10.1073/pnas.061500998 . ПМК 31209 . ПМИД 11274447 .
- ^ Jump up to: а б с Пельтье Дж., Эмануэльссон О., Калуме Д., Иттерберг Дж., Фризо Г., Руделла А., Либерлес Д., Седерберг Л., Ропсторфф П., фон Хейне Г. , ван Вейк К.Дж. (2002). «Центральные функции люменального и периферического тилакоидного протеома Arabidopsis, определенные экспериментальным путем и общегеномным прогнозированием» . Растительная клетка . 14 (1): 211–36. дои : 10.1105/tpc.010304 . ПМК 150561 . ПМИД 11826309 .
- ^ ван Вейк К. (2004). «Пластидная протеомика». Растительная Физиол Биохимия . 42 (12): 963–77. дои : 10.1016/j.plaphy.2004.10.015 . ПМИД 15707834 .
- ^ Jump up to: а б Фрисо Г., Джакомелли Л., Иттерберг А., Пельтье Дж., Руделла А., Сан К., Вейк К. (2004). «Углубленный анализ протеома тилакоидной мембраны хлоропластов Arabidopsis thaliana: новые белки, новые функции и база данных пластидных протеомов» . Растительная клетка . 16 (2): 478–99. дои : 10.1105/tpc.017814 . ПМК 341918 . ПМИД 14729914 . - База данных пластидных протеомов
- ^ Клеффманн Т., Хирш-Хоффманн М., Груиссем В., Багинский С. (2006). «plprot: обширная база данных протеомов для различных типов пластид». Физиол растительной клетки . 47 (3): 432–6. дои : 10.1093/pcp/pcj005 . ПМИД 16418230 . – База данных пластидных белков
- ^ Пельтье Дж., Фрисо Г., Калуме Д., Ропсторфф П., Нильссон Ф., Адамска И., ван Вейк К. (2000). «Протеомика хлоропластов: систематическая идентификация и целевой анализ люменальных и периферических тилакоидных белков» . Растительная клетка . 12 (3): 319–41. дои : 10.1105/tpc.12.3.319 . ПМК 139834 . ПМИД 10715320 .
- ^ Венер А.В., Охад И., Андерссон Б. (1998). «Фосфорилирование белков и окислительно-восстановительное зондирование в тилакоидах хлоропластов». Карр Опин Растительная Биол . 1 (3): 217–23. дои : 10.1016/S1369-5266(98)80107-6 . ПМИД 10066592 .
- ^ Шоке Ю., Вострикофф К., Рембо Б., Зито Ф., Жирар-Баску Дж., Драпье Д., Воллман Ф. (2001). «Контролируемая сборкой регуляция трансляции генов хлоропластов». Биохим Соц Транс . 29 (Часть 4): 421–6. дои : 10.1042/BST0290421 . ПМИД 11498001 .
- ^ Минай Л., Вострикофф К., Воллман Ф., Шоке Ю. (2006). «Биогенез хлоропластов ядер фотосистемы II включает в себя серию контролируемых сборкой шагов, которые регулируют трансляцию» . Растительная клетка . 18 (1): 159–75. дои : 10.1105/tpc.105.037705 . ПМЦ 1323491 . ПМИД 16339851 .
- ^ Аллен Дж., Пфанншмидт Т. (2000). «Балансирование двух фотосистем: фотосинтетический перенос электронов управляет транскрипцией генов реакционных центров в хлоропластах» . Philos Trans R Soc Lond B Biol Sci . 355 (1402): 1351–9. дои : 10.1098/rstb.2000.0697 . ПМК 1692884 . ПМИД 11127990 .
- ^ Jump up to: а б Гутенсон М., Фан Э., Фрилингсдорф С., Ханнер П., Хоу Б., Хуст Б., Клёсген Р. (2006). «Ток, Тик, Тат и др.: структура и функции механизмов транспорта белков в хлоропластах». Дж. Физиол растений . 163 (3): 333–47. дои : 10.1016/j.jplph.2005.11.009 . ПМИД 16386331 .
- ^ Ягендорф А.Т. и Э. Урибе (1966). «Образование АТФ, вызванное кислотно-щелочным переходом хлоропластов шпината» . Учеб. Натл. акад. наук. США . 55 (1): 170–177. Бибкод : 1966PNAS...55..170J . дои : 10.1073/pnas.55.1.170 . ПМЦ 285771 . ПМИД 5220864 .
- ^ Эрреро, Антония; Флорес, Энрике, ред. (2008). Цианобактерии: молекулярная биология, геномика и эволюция (1-е изд.). Кайстер Академик Пресс. ISBN 978-1-904455-15-8 .
- ^ Нево Р., Чаруви Д., Шимони Э., Шварц Р., Каплан А., Охад И., Райх З. (2007). «Перфорация и соединение тилакоидных мембран обеспечивают внутриклеточный транспорт цианобактерий» . ЭМБО Дж . 26 (5): 1467–1473. дои : 10.1038/sj.emboj.7601594 . ПМЦ 1817639 . ПМИД 17304210 .
- ^ Олив, Дж; Аджлани, Дж; Астье, К; Рекуврер, М; Вернотт, К. (1997). «Ультраструктура и световая адаптация мутантов фикобилисом Synechocystis PCC 6803». Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1319 (2–3): 275–282. дои : 10.1016/S0005-2728(96)00168-5 .
- ^ Надь, Г; Поссельт, Д; Ковач, Л; Холм, Дж. К.; Сабо, М; Уги, Б; Роста, Л; Питерс, Дж; Тимминс, П; Гараб, Г. (1 июня 2011 г.). «Обратимые мембранные реорганизации во время фотосинтеза in vivo: выявлены методом малоуглового рассеяния нейтронов» (PDF) . Биохимический журнал . 436 (2): 225–30. дои : 10.1042/BJ20110180 . ПМИД 21473741 .
Источники учебников
[ редактировать ]- Хеллер, Х. Крейг; Орианс, Гордан Х.; Первс, Уильям К. и Садава, Дэвид (2004). ЖИЗНЬ: Наука биологии (7-е изд.). Sinauer Associates, Inc. ISBN 978-0-7167-9856-9 .
- Рэйвен, Питер Х.; Рэй Ф. Эверт; Сьюзан Э. Эйххорн (2005). Биология растений (7-е изд.). Нью-Йорк: Издатели WH Freeman and Company. стр. 115–127 . ISBN 978-0-7167-1007-3 .
- Эрреро, Антония; Флорес, Энрике, ред. (2008). Цианобактерии: молекулярная биология, геномика и эволюция (1-е изд.). Кайстер Академик Пресс. ISBN 978-1-904455-15-8 .