АТФ-синтаза
| АТФ-синтаза | |||
|---|---|---|---|
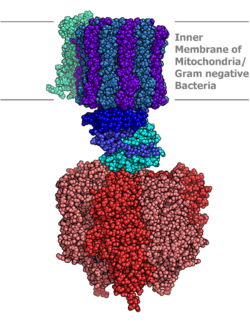 Молекулярная модель АТФ-синтазы, определенная методом рентгеновской кристаллографии . Статор здесь не показан. | |||
| Идентификаторы | |||
| Номер ЕС. | 7.1.2.2 | ||
| Номер CAS. | 9000-83-3 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
АТФ-синтаза представляет собой фермент , который катализирует образование молекулы-хранилища энергии аденозинтрифосфата (АТФ) с использованием аденозиндифосфата (АДФ) и неорганического фосфата (P i ). АТФ-синтаза представляет собой молекулярную машину . Общая реакция, катализируемая АТФ-синтазой:
- АДФ + П я + 2Н + выход ⇌ АТФ + Н 2 О + 2Н + в
АТФ-синтаза расположена через клеточную мембрану и образует отверстие, через которое протоны могут пересекать области с высокой концентрацией в области с низкой концентрацией, передавая энергию для синтеза АТФ. Этот электрохимический градиент генерируется цепью переноса электронов и позволяет клеткам хранить энергию в виде АТФ для последующего использования. В прокариотических клетках АТФ-синтаза располагается через плазматическую мембрану , а в эукариотических клетках — через внутреннюю митохондриальную мембрану . У организмов, способных к фотосинтезу, через тилакоидную мембрану имеется также АТФ-синтаза , которая у растений расположена в хлоропластах , а у цианобактерий — в цитоплазме .
Эукариотические АТФ-синтазы представляют собой F-АТФазы , работающие «обратно» по отношению к АТФазе . Данная статья посвящена в основном этому типу. F-АТФаза состоит из двух основных субъединиц, FO и F 1 , которые имеют вращательный моторный механизм, позволяющий производить АТФ. [1] [2]
Номенклатура [ править ]
Фракция F 1 получила свое название от термина «Фракция 1», а F O (записанная буквой «о», а не «ноль») получила свое название от того, что она является фракцией связывания олигомицина , типа антибиотика природного происхождения, который способен ингибировать FO - единицу АТФ-синтазы. [3] [4] Эти функциональные области состоят из разных белковых субъединиц — см. таблицы. Этот фермент используется в синтезе АТФ посредством аэробного дыхания.
Структура и функции [ править ]


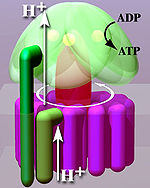
Расположенная внутри тилакоидной мембраны и внутренней мембраны митохондрий , АТФ-синтаза состоит из двух областей F O и F 1 . F O вызывает вращение F 1 и состоит из c-кольца и субъединиц a, двух b, F6. F 1 состоит из субъединиц α, β, γ и δ. F 1 имеет водорастворимую часть, способную гидролизовать АТФ. . области С другой стороны, FO имеет в основном гидрофобные F O F 1 создает путь для движения протонов через мембрану. [7]
F 1 регион [ править ]
Часть F 1 АТФ-синтазы гидрофильна и отвечает за гидролиз АТФ. Единица F 1 выступает в пространство митохондриального матрикса . Субъединицы α и β образуют гексамер с 6 сайтами связывания. Три из них каталитически неактивны и связывают АДФ.
Три другие субъединицы катализируют синтез АТФ. Остальные субъединицы F 1 γ, δ и ε являются частью вращательного моторного механизма (ротор/ось). Субъединица γ позволяет β претерпевать конформационные изменения (т.е. закрытое, полуоткрытое и открытое состояния), которые позволяют АТФ связываться и высвобождаться после синтеза. Частица F 1 имеет большие размеры и ее можно увидеть в просвечивающем электронном микроскопе путем негативного окрашивания. [8] Это частицы диаметром 9 нм, которые пронизывают внутреннюю мембрану митохондрий.
| Субъединица | Человеческий ген | Примечание |
|---|---|---|
| альфа | АТФ5А1 , АТФАФ2 | |
| бета | АТФ5В , АТФАФ1 | |
| гамма | АТФ5С1 | |
| дельта | АТФ5Д | Митохондриальная «дельта» — это бактериальный/хлоропластический эпсилон. |
| эпсилон | АТФ5Е | Уникальный для митохондрий. |
| ОСКП | АТФ5О | В бактериальной и хлоропластической версиях называется «дельта». |
F O регион [ править ]

F O представляет собой водонерастворимый белок с восемью субъединицами и трансмембранным кольцом. Кольцо имеет тетрамерную форму с белком спираль-петля-спираль , который претерпевает конформационные изменения при протонировании и депротонировании, заставляя соседние субъединицы вращаться, вызывая вращение FO , что затем также влияет на конформацию F 1 , что приводит к переключению состояний. альфа- и бета-субъединиц. Область F O АТФ-синтазы представляет собой протонную пору, встроенную в мембрану митохондрий. Он состоит из трех основных субъединиц: а, b и с. Шесть субъединиц c составляют кольцо ротора, а субъединица b образует стебель, соединяющийся с F 1 OSCP, который предотвращает вращение гексамера αβ. Субъединица a соединяет b с кольцом c. [11] У людей есть шесть дополнительных субъединиц: d , e , f , g , F6 и 8 (или A6L). Эта часть фермента расположена во внутренней мембране митохондрий и связывает транслокацию протона с вращением, вызывающим синтез АТФ в области F1 .
У эукариот митохондриальный FO образует димеры, изгибающие мембрану. Эти димеры самоорганизуются в длинные ряды на концах крист , что, возможно, является первым этапом формирования крист. [12] Атомная модель димерной области FO дрожжей была определена методом криоЭМ с общим разрешением 3,6 Å. [13]
| Субъединица | Человеческий ген |
|---|---|
| а | МТ-АТФ6 |
| б | АТФ5ПБ |
| с | АТФ5Г1 , АТФ5Г2 , АТФ5Г3 |
Модель привязки [ править ]


В 1960-1970-х годах Пол Бойер , профессор Калифорнийского университета в Лос- Анджелесе, разработал теорию механизма изменения связывания, или триггера, которая постулировала, что синтез АТФ зависит от конформационных изменений АТФ-синтазы, вызванных вращением гамма-субъединицы. Исследовательская группа Джона Э. Уокера , работавшего тогда в Лаборатории молекулярной биологии MRC в Кембридже , кристаллизовала каталитический домен F 1 АТФ-синтазы. Эта структура, являвшаяся на тот момент крупнейшей асимметричной белковой структурой, указывала на то, что модель ротационного катализа Бойера, по сути, верна. За объяснение этого Бойер и Уокер получили половину Нобелевской премии по химии 1997 года .
Кристаллическая структура F 1 демонстрирует чередующиеся альфа- и бета- субъединицы (по три каждой), расположенные как сегменты апельсина вокруг вращающейся асимметричной гамма-субъединицы. Согласно современной модели синтеза АТФ (известной как альтернирующая каталитическая модель), трансмембранный потенциал, создаваемый катионами протонов (H+), доставляемыми электрон-транспортной цепью, вытесняет катионы протонов (H+) из межмембранного пространства через мембрану через мембрану. F O- область АТФ-синтазы. Часть FO ( кольцо c-субъединиц ) вращается при прохождении протонов через мембрану. С -кольцо плотно прикреплено к асимметричному центральному стеблю (состоящему в основном из гамма-субъединицы), заставляя его вращаться внутри альфа- 3- бета- 3 F1 , в результате чего три каталитических сайта связывания нуклеотидов претерпевают ряд конформационных изменений, которые приводят к синтезу АТФ. Главным субъединицам F1 препятствует вращению в симпатии к центральному ротору стебля с помощью периферического стебля, который соединяет альфа- 3- бета -3 с невращающейся частью F. О. Структура интактной АТФ-синтазы в настоящее время известна с низким разрешением благодаря с помощью электронной криомикроскопии исследованиям комплекса (крио-ЭМ). Крио-ЭМ модель АТФ-синтазы предполагает, что периферический стебель представляет собой гибкую структуру, которая обволакивает комплекс при соединении F 1 с F O . При правильных условиях ферментативная реакция также может осуществляться в обратном направлении, при этом гидролиз АТФ вызывает перекачку протонов через мембрану.
Механизм изменения связывания включает в себя циклическое перемещение активного сайта β-субъединицы между тремя состояниями. [14] В «рыхлом» состоянии АДФ и фосфат поступают в активный центр; на соседней диаграмме это показано розовым цветом. Затем фермент претерпевает изменение формы и сближает эти молекулы, при этом активный центр оказывается в «плотном» состоянии (показано красным), связывая вновь полученную молекулу АТФ с очень высоким сродством . Наконец, активный сайт возвращается в открытое состояние (оранжевый), высвобождая АТФ и связывая больше АДФ и фосфата, готовясь к следующему циклу производства АТФ. [15]
роль Физиологическая
Как и другие ферменты, активность F 1 F O АТФ-синтазы обратима. Достаточно большие количества АТФ заставляют его создавать трансмембранный протонный градиент . Это используется ферментирующими бактериями, которые не имеют цепи переноса электронов, а скорее гидролизуют АТФ с образованием протонного градиента, который они используют для управления жгутиками и транспортировки питательные вещества попадают в клетку.
У дышащих бактерий в физиологических условиях АТФ-синтаза, как правило, работает в противоположном направлении, создавая АТФ, используя в движущую силу протонов , создаваемую цепью переноса электронов качестве источника энергии . Общий процесс создания энергии таким образом называется окислительным фосфорилированием .Тот же процесс происходит в митохондриях , где АТФ-синтаза расположена во внутренней митохондриальной мембране, а F 1 -часть выступает в митохондриальный матрикс . Накачивая протон-катионы в матрикс, АТФ-синтаза превращает АДФ в АТФ.
Эволюция [ править ]
АТФ Считается, что эволюция -синтазы имела модульный характер, в результате чего две функционально независимые субъединицы стали ассоциироваться и приобрели новую функциональность. [16] [17] Эта ассоциация, по-видимому, возникла на ранних этапах эволюционной истории, поскольку по существу одна и та же структура и активность ферментов АТФ-синтазы присутствуют во всех царствах жизни. [16] Синтаза F-АТФ демонстрирует большое функциональное и механистическое сходство с V-АТФазой . [18] Однако в то время как F-АТФ-синтаза генерирует АТФ, используя протонный градиент, V-АТФаза генерирует протонный градиент за счет АТФ, генерируя значения pH всего лишь 1. [19]
Область F 1 также демонстрирует значительное сходство с гексамерными ДНК-хеликазами (особенно с фактором Rho ), а вся область фермента демонстрирует некоторое сходство с H. +
-с питанием T3SS или жгутиковых двигательных комплексов. [18] [20] [21] α3β3 ; хеликазами Гексамер F1 области демонстрирует значительное структурное сходство с гексамерными ДНК- оба образуют кольцо с 3-кратной вращательной симметрией с центральной порой. Роль обоих зависит от относительного вращения макромолекулы внутри поры; ДНК-хеликазы используют спиральную форму ДНК, чтобы управлять своим движением вдоль молекулы ДНК и обнаруживать суперспирализацию, тогда как гексамер α 3 β 3 использует конформационные изменения за счет вращения субъединицы γ для запуска ферментативной реакции. [22]
Н +
двигатель частицы F O демонстрирует большое функциональное сходство с H +
моторы, приводящие в движение жгутики. [18] Оба имеют кольцо из множества небольших альфа-спиральных белков, которые вращаются относительно близлежащих неподвижных белков с помощью H. +
Градиент потенциала как источник энергии. Однако эта связь незначительна, поскольку общая структура жгутиковых моторов намного сложнее, чем структура частицы F O , а кольцо, состоящее примерно из 30 вращающихся белков, намного больше, чем 10, 11 или 14 спиральных белков в F O. сложный. Однако более поздние структурные данные показывают, что кольцо и ножка структурно подобны частице F 1 . [21]
Теория модульной эволюции происхождения АТФ-синтазы предполагает, что две субъединицы с независимыми функциями: ДНК-хеликаза с АТФазной активностью и H +
мотор, были способны связываться, и вращение мотора приводило к обратному изменению АТФазной активности геликазы. [16] [22] Затем этот комплекс стал более эффективным и в конечном итоге превратился в современные сложные АТФ-синтазы. Альтернативно, ДНК-хелиза/ H +
двигательный комплекс мог иметь H +
активность насоса с АТФазной активностью геликазы, управляющей H +
двигатель в обратном направлении. [16] Возможно, он развился, чтобы осуществлять обратную реакцию и действовать как АТФ-синтаза. [17] [23] [24]
Ингибиторы [ править ]
Открыто множество природных и синтетических ингибиторов АТФ-синтазы. [25] Они были использованы для исследования структуры и механизма АТФ-синтазы. Некоторые из них могут иметь терапевтическое значение. Существует несколько классов ингибиторов АТФ-синтазы, включая пептидные ингибиторы, полифенольные фитохимические вещества, поликетиды, оловоорганические соединения, полиеновые производные α-пирона, катионные ингибиторы, аналоги субстратов, модификаторы аминокислот и другие различные химические вещества. [25] Некоторыми из наиболее часто используемых ингибиторов АТФ-синтазы являются олигомицин и DCCD .
В разных организмах [ править ]
Бактерии [ править ]
АТФ-синтаза E. coli — это простейшая известная форма АТФ-синтазы, имеющая 8 различных типов субъединиц. [11]
Бактериальные F-АТФазы иногда могут действовать наоборот, превращая их в АТФазу. [26] Некоторые бактерии не имеют F-АТФазы и используют АТФазу A/V-типа двунаправленно. [9]
Дрожжи [ править ]
АТФ-синтаза дрожжей — одна из наиболее изученных АТФ-синтаз эукариот; пять субъединиц F 1 , восемь субъединиц FO и семь ассоциированных белков. и были идентифицированы [7] Большинство этих белков имеют гомологи у других эукариот. [27] [28] [29] [30]
Растение [ править ]
У растений АТФ-синтаза также присутствует в хлоропластах (CF 1 F O -АТФ-синтаза). Фермент интегрирован в тилакоидную мембрану; часть CF 1 прилипает к строме темновые реакции фотосинтеза (также называемые светонезависимыми реакциями или циклом Кальвина , где происходят ) и синтез АТФ. Общая структура и каталитический механизм АТФ-синтазы хлоропластов почти такие же, как у бактериального фермента. Однако в хлоропластах движущую силу протонов генерирует не дыхательная цепь транспорта электронов, а первичные фотосинтетические белки. Синтаза имеет вставку из 40 аминокислот в гамма-субъединицу, которая подавляет бесполезную активность в темноте. [31]
млекопитающее [ править ]
АТФ-синтаза, выделенная из митохондрий сердца быка ( Bos taurus ), с точки зрения биохимии и структуры является наиболее изученной АТФ-синтазой. Говяжье сердце используется в качестве источника фермента из-за высокой концентрации митохондрий в сердечной мышце. Их гены имеют близкую гомологию с АТФ-синтазами человека. [32] [33] [34]
Гены человека, кодирующие компоненты АТФ-синтаз:
- АТФ5А1
- АТФ5В
- ATP5C1 , ATP5D , ATP5E , ATP5F1 , ATP5G1 , ATP5G2 , ATP5G3 , ATP5H , ATP5I , ATP5J , ATP5J2 , ATP5L , ATP5O
- МТ-АТФ6 , МТ-АТФ8
Другие эукариоты [ править ]
Эукариоты, принадлежащие к некоторым дивергентным линиям, имеют совершенно особую организацию АТФ-синтазы. АТФ -синтаза эвгленозоа образует димер с головкой F 1 в форме бумеранга , как и другие митохондриальные АТФ-синтазы, но субкомплекс FO имеет множество уникальных субъединиц. Здесь используется кардиолипин . Ингибирующий IF 1 также связывается по-другому, как и трипаносоматида . [35]
Архея [ править ]
У архей обычно нет F-АТФазы. Вместо этого они синтезируют АТФ, используя А-АТФазу/синтазу, вращающуюся машину, структурно похожую на V-АТФазу , но в основном функционирующую как АТФ-синтазу. [26] Считается, что, как и бактериальная F-АТФаза, она также действует как АТФаза. [9]
LUCA и ранее [ править ]
Связь генов F-АТФазы и порядок генов широко сохраняются в древних линиях прокариот, что означает, что эта система уже существовала до появления последнего универсального общего предка , LUCA. [36]
См. также [ править ]
- Белок АТФ10, необходимый для сборки сектора ФО митохондриального АТФазного комплекса.
- хлоропласт
- Цепь переноса электронов
- флавопротеин
- Митохондрия
- Окислительное фосфорилирование
- Р-АТФаза
- Протонный насос
- Вращающееся передвижение в живых системах.
- Трансмембранная АТФаза
- V-АТФаза
Ссылки [ править ]
- ^ Окуно Д., Иино Р., Нодзи Х. (июнь 2011 г.). «Вращение и структура FoF1-АТФ-синтазы» . Журнал биохимии . 149 (6): 655–664. дои : 10.1093/jb/mvr049 . ПМИД 21524994 .
- ^ Юнге В., Нельсон Н. (июнь 2015 г.). «АТФ-синтаза» . Ежегодный обзор биохимии . 84 : 631–657. doi : 10.1146/annurev-biochem-060614-034124 . ПМИД 25839341 .
- ^ Кагава Ю., Рэкер Э. (май 1966 г.). «Частичное разрешение ферментов, катализирующих окислительное фосфорилирование. 8. Свойства фактора, придающего чувствительность к олигомицинам митохондриальной аденозинтрифосфатазы» . Журнал биологической химии . 241 (10): 2461–2466. дои : 10.1016/S0021-9258(18)96640-8 . ПМИД 4223640 .
- ^ Маккарти RE (ноябрь 1992 г.). «ВЗГЛЯД РАСТИТЕЛЬНОГО БИОХИМИКА НА H +-АТФАЗЫ И АТФ-СИНТАЗЫ». Журнал экспериментальной биологии . 172 (Часть 1): 431–441. дои : 10.1242/jeb.172.1.431 . ПМИД 9874753 .
- ^ ВВП : 5ARA ; Чжоу А., Рохоу А., Шеп Д.Г., Бэйсон Дж.В., Монтгомери М.Г., Уокер Дж.Е. и др. (октябрь 2015 г.). «Структура и конформационные состояния бычьей митохондриальной АТФ-синтазы с помощью крио-ЭМ» . электронная жизнь . 4 : е10180. doi : 10.7554/eLife.10180 . ПМЦ 4718723 . ПМИД 26439008 .
- ^ Гудселл Д (декабрь 2005 г.). «АТФ-синтаза» . Молекула месяца . дои : 10.2210/rcsb_pdb/mom_2005_12 .
- ↑ Перейти обратно: Перейти обратно: а б Велюр Дж., Помар П., Субанье В., Шпаннагель С., Вайе Дж., Арселин Г., Грейвс П.В. (май 2000 г.). «Организация дрожжевой АТФ-синтазы F (0): исследование, основанное на мутантах цистеина, тиоловой модификации и реагентах сшивания» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1458 (2–3): 443–456. дои : 10.1016/S0005-2728(00)00093-1 . ПМИД 10838057 .
- ^ Фернандес Моран Х., Ода Т., Блэр П.В., Грин Д.Э. (июль 1964 г.). «Макромолекулярная повторяющаяся единица структуры и функции митохондрий. Коррелированные электронно-микроскопические и биохимические исследования изолированных митохондрий и субмитохондриальных частиц говяжьей сердечной мышцы» . Журнал клеточной биологии . 22 (1): 63–100. дои : 10.1083/jcb.22.1.63 . ПМК 2106494 . ПМИД 14195622 .
- ↑ Перейти обратно: Перейти обратно: а б с Стюарт А.Г., Лэминг Э.М., Собти М., Сток Д. (апрель 2014 г.). «Вращающиеся АТФазы — динамические молекулярные машины» . Современное мнение в области структурной биологии . 25 : 40–48. дои : 10.1016/j.sbi.2013.11.013 . ПМИД 24878343 .
- ^ ПДБ : 1ВЗС ; Карбахо Р.Дж., Сильвестр Дж.А., Рансуик М.Дж., Уокер Дж.Э., Нойхаус Д. (сентябрь 2004 г.). «Структура раствора субъединицы F (6) из области периферического стебля АТФ-синтазы из митохондрий бычьего сердца». Журнал молекулярной биологии . 342 (2): 593–603. дои : 10.1016/j.jmb.2004.07.013 . ПМИД 15327958 .
- ↑ Перейти обратно: Перейти обратно: а б Ахмад З., Окафор Ф., Лафлин Т.Ф. (2011). «Роль заряженных остатков в каталитических сайтах АТФ-синтазы Escherichia coli» . Журнал аминокислот . 2011 : 785741. doi : 10.4061/2011/785741 . ПМК 3268026 . ПМИД 22312470 .
- ^ Блюм ТБ, Хан А., Мейер Т., Дэвис К.М., Кюльбрандт В. (март 2019 г.). «Димеры митохондриальной АТФ-синтазы вызывают искривление мембраны и самоорганизуются в ряды» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (10): 4250–4255. Бибкод : 2019PNAS..116.4250B . дои : 10.1073/pnas.1816556116 . ПМК 6410833 . ПМИД 30760595 .
- ^ Го Х., Бюлер С.А., Рубинштейн Дж.Л. (ноябрь 2017 г.). «Атомная модель димерной области FO митохондриальной АТФ-синтазы» . Наука . 358 (6365): 936–940. Бибкод : 2017Sci...358..936G . дои : 10.1126/science.aao4815 . ПМК 6402782 . ПМИД 29074581 .
- ^ Грессер М.Дж., Майерс Дж.А., Бойер П.Д. (октябрь 1982 г.). «Кооперативность каталитических участков митохондриальной аденозинтрифосфатазы F 1 говяжьего сердца . Корреляции начальной скорости, связанных промежуточных продуктов и измерений кислородного обмена с чередующейся трехсайтовой моделью» . Журнал биологической химии . 257 (20): 12030–12038. дои : 10.1016/S0021-9258(18)33672-X . ПМИД 6214554 .
- ^ Накамото Р.К., Бэйлис Скэнлон Дж.А., Аль-Шави М.К. (август 2008 г.). «Вращательный механизм АТФ-синтазы» . Архив биохимии и биофизики . 476 (1): 43–50. дои : 10.1016/j.abb.2008.05.004 . ПМК 2581510 . ПМИД 18515057 .
- ↑ Перейти обратно: Перейти обратно: а б с д Деринг С., Эрментраут Б., Остер Г. (декабрь 1995 г.). «Роторные ДНК-моторы» . Биофизический журнал . 69 (6): 2256–2267. Бибкод : 1995BpJ....69.2256D . дои : 10.1016/S0006-3495(95)80096-2 . ПМЦ 1236464 . ПМИД 8599633 .
- ↑ Перейти обратно: Перейти обратно: а б Крофтс А. «Лекция 10: АТФ-синтаза» . Науки о жизни в Университете Иллинойса в Урбане-Шампейне .
- ↑ Перейти обратно: Перейти обратно: а б с «АТФ-синтаза» . База данных ИнтерПро .
- ^ Бейенбах К.В., Вечорек Х (февраль 2006 г.). «H+ АТФаза V-типа: молекулярная структура и функции, физиологическая роль и регуляция» . Журнал экспериментальной биологии . 209 (Часть 4): 577–589. дои : 10.1242/jeb.02014 . ПМИД 16449553 .
- ^ Скордалакес Э., Бергер Дж. М. (июль 2003 г.). «Структура терминатора транскрипции Rho: механизм узнавания мРНК и загрузки геликазы» . Клетка . 114 (1): 135–146. дои : 10.1016/S0092-8674(03)00512-9 . ПМИД 12859904 . S2CID 5765103 .
- ↑ Перейти обратно: Перейти обратно: а б Имада К., Минамино Т., Учида Ю., Киношита М., Намба К. (март 2016 г.). «Понимание экспорта жгутиков типа III, выявленное благодаря сложной структуре АТФазы типа III и ее регулятора» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (13): 3633–3638. Бибкод : 2016PNAS..113.3633I . дои : 10.1073/pnas.1524025113 . ПМЦ 4822572 . ПМИД 26984495 .
- ↑ Перейти обратно: Перейти обратно: а б Мартинес Л.О., Жаке С., Эстев Х.П., Роллан С., Кабесон Е., Шампанское Е. и др. (январь 2003 г.). «Эктопическая бета-цепь АТФ-синтазы является рецептором аполипопротеина AI при эндоцитозе ЛПВП в печени». Природа . 421 (6918): 75–79. Бибкод : 2003Natur.421...75M . дои : 10.1038/nature01250 . ПМИД 12511957 . S2CID 4333137 .
- ^ Кросс Р.Л., Таиз Л. (январь 1990 г.). «Дупликация генов как средство изменения соотношения H + / АТФ во время эволюции FO F 1 » АТФаз и синтаз . Письма ФЭБС . 259 (2): 227–229. дои : 10.1016/0014-5793(90)80014-а . ПМИД 2136729 . S2CID 32559858 .
- ^ Кросс Р.Л., Мюллер В. (октябрь 2004 г.). «Эволюция АТФ-синтаз и АТФаз A-, F- и V-типов: изменения в функции и изменения в соотношении H + / АТФ» . Письма ФЭБС . 576 (1–2): 1–4. дои : 10.1016/j.febslet.2004.08.065 . ПМИД 15473999 . S2CID 25800744 .
- ↑ Перейти обратно: Перейти обратно: а б Хонг С., Педерсен П.Л. (декабрь 2008 г.). «АТФ-синтаза и действие ингибиторов, используемых для изучения ее роли в здоровье человека, заболеваниях и других научных областях» . Обзоры микробиологии и молекулярной биологии . 72 (4): 590–641, Оглавление. дои : 10.1128/MMBR.00016-08 . ПМК 2593570 . ПМИД 19052322 .
- ↑ Перейти обратно: Перейти обратно: а б Кюльбрандт В., Дэвис К.М. (январь 2016 г.). «Вращающиеся АТФазы: новый поворот в древней машине». Тенденции биохимических наук . 41 (1): 106–116. дои : 10.1016/j.tibs.2015.10.006 . ПМИД 26671611 .
- ^ Девениш Р.Дж., Прескотт М., Руку X, Нэгли П. (май 2000 г.). «Понимание сборки и функционирования АТФ-синтазы посредством молекулярно-генетических манипуляций субъединицами митохондриального ферментного комплекса дрожжей» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1458 (2–3): 428–442. дои : 10.1016/S0005-2728(00)00092-X . ПМИД 10838056 .
- ^ Кабалисваран В., Пури Н., Уокер Дж.Э., Лесли А.Г., Мюллер Д.М. (ноябрь 2006 г.). «Новые особенности ротационного каталитического механизма, выявленные в структуре дрожжевой F 1 АТФазы» . Журнал ЭМБО . 25 (22): 5433–5442. дои : 10.1038/sj.emboj.7601410 . ПМК 1636620 . ПМИД 17082766 .
- ^ Сток Д., Лесли А.Г., Уокер Дж.Э. (ноябрь 1999 г.). «Молекулярная архитектура роторного двигателя в АТФ-синтазе». Наука . 286 (5445): 1700–1705. дои : 10.1126/science.286.5445.1700 . ПМИД 10576729 .
- ^ Лю С., Чарльзворт Т.Дж., Бэйсон Дж.В., Монтгомери М.Г., Харбор М.Э., Фернли И.М., Уокер Дж.Е. (май 2015 г.). «Очистка и характеристика АТФ-синтазных комплексов из митохондрий четырех видов грибов» . Биохимический журнал . 468 (1): 167–175. дои : 10.1042/BJ20150197 . ПМЦ 4422255 . ПМИД 25759169 .
- ^ Хан А., Вонк Дж., Миллс DJ, Мейер Т., Кюльбрандт В. (май 2018 г.). «Структура, механизм и регуляция АТФ-синтазы хлоропластов» . Наука . 360 (6389): eaat4318. дои : 10.1126/science.aat4318 . ПМК 7116070 . ПМИД 29748256 .
- ^ Абрахамс Дж.П., Лесли А.Г., Люттер Р., Уокер Дж.Э. (август 1994 г.). «Структура F1-АТФазы из митохондрий бычьего сердца с разрешением 2,8 А». Природа . 370 (6491): 621–628. Бибкод : 1994Natur.370..621A . дои : 10.1038/370621a0 . ПМИД 8065448 . S2CID 4275221 .
- ^ Гиббонс С., Монтгомери М.Г., Лесли А.Г., Уокер Дж.Э. (ноябрь 2000 г.). «Структура центрального стебля бычьей F (1)-АТФазы при разрешении 2,4 А». Структурная биология природы . 7 (11): 1055–1061. дои : 10.1038/80981 . ПМИД 11062563 . S2CID 23229994 .
- ^ Менц Р.И., Уокер Дж.Э., Лесли А.Г. (август 2001 г.). «Структура бычьей митохондриальной F (1)-АТФазы с нуклеотидом, связанным со всеми тремя каталитическими сайтами: значение для механизма ротационного катализа» . Клетка . 106 (3): 331–341. дои : 10.1016/s0092-8674(01)00452-4 . ПМИД 11509182 . S2CID 1266814 .
- ^ Мюлейп А., МакКомас С.Э., Амунц А. (ноябрь 2019 г.). «Структура митохондриальной АТФ-синтазы со связанным нативным кардиолипином» . электронная жизнь . 8 : е51179. дои : 10.7554/eLife.51179 . ПМК 6930080 . ПМИД 31738165 .
- «Отличный от остальных» . электронная жизнь . 24 декабря 2019 г.
- ^ Мацке, Нью-Джерси, Лин А., Стоун М., Бейкер М.А. (июль 2021 г.). «Аппарат экспорта жгутиков и АТФ-синтетаза: гомология, подтвержденная синтенией, предшествовавшей последнему универсальному общему предку». Биоэссе . 43 (7): e2100004. doi : 10.1002/bies.202100004 . hdl : 2292/55176 . ПМИД 33998015 . S2CID 234747849 .
Дальнейшее чтение [ править ]
- Ник Лейн: Жизненно важный вопрос: энергия, эволюция и происхождение сложной жизни , WW Norton, 20 июля 2015 г., ISBN 978-0393088816 (ссылка указывает на рисунок 10, показывающий модель АТФ-синтазы)
Внешние ссылки [ править ]
- Борис А. Фенюк: «АТФ-синтаза — великолепная молекулярная машина»
- Хорошо иллюстрированная лекция Энтони Крофтса по АТФ-синтазе из Университета Иллинойса в Урбана-Шампейн .
- Протон и натрий, транслоцирующие АТФазы F-типа, V-типа и A-типа в базе данных OPM
- Нобелевская премия по химии 1997 г. Полу Д. Бойеру и Джону Э. Уокеру за ферментативный механизм синтеза АТФ; и Йенсу К. Скоу за открытие фермента, переносящего ионы, Na. +
, К +
-АТФаза. - Гарвардский мультимедийный производственный сайт — Видео — анимация синтеза АТФ
- Дэвид Гудселл: «АТФ-синтаза — молекула месяца». Архивировано 5 сентября 2015 г. в Wayback Machine.