Комплекс пируватдегидрогеназы

Комплекс пируватдегидрогеназы ( PDC ) представляет собой комплекс из трех ферментов , которые превращают пируват в ацетил-КоА посредством процесса, называемого декарбоксилированием пирувата . [1] Ацетил-КоА затем может использоваться в цикле лимонной кислоты для осуществления клеточного дыхания , и этот комплекс связывает гликолиза метаболический путь с циклом лимонной кислоты . Декарбоксилирование пирувата также известно как «реакция пируватдегидрогеназы», поскольку оно также включает окисление пирувата. [2]
Этот мультиферментный комплекс структурно и функционально связан с мультиферментными комплексами оксоглутаратдегидрогеназы и оксокислотной дегидрогеназы с разветвленной цепью .
Реакция
[ редактировать ]Реакция, катализируемая пируватдегидрогеназным комплексом:
| пируват | комплекс пируватдегидрогеназы | ацетил-КоА | |
 | |||
| КоА-SH + НАД + | СО 2 + НАДН + Н + | ||
Структура
[ редактировать ]Пируватдегидрогеназа (E1)
[ редактировать ]
Субъединица E1, называемая субъединицей пируватдегидрогеназы , представляет собой либо гомодимер (содержащий две цепи «ɑ», например, у Escherichia coli ), либо гетеротетрамер из двух разных цепей (две цепи «ɑ» и две «ꞵ»). Ион магния образует 4-координационный комплекс с тремя полярными аминокислотными остатками (Asp, Asn и Tyr), расположенными в альфа-цепи, и тиаминдифосфата (TPP), непосредственно участвующим в декарбоксилировании пирувата кофактором . [3] [4]
Дигидролипоилтрансацетилаза (E2)
[ редактировать ]Субъединица E2, или дигидролипоилацетилтрансфераза, как у прокариот, так и у эукариот, обычно состоит из трех доменов. N-концевой домен (липоильный домен) состоит из 1–3 липоильных групп примерно по 80 аминокислот каждая. Домен, связывающий периферическую субъединицу (PSBD), служит местом селективного связывания для других доменов субъединиц E1 и E3. Наконец, С-концевой (каталитический) домен катализирует перенос ацетильных групп и синтез ацетил-КоА. [5] У гаммапротеобактерий 24 копии Е2 образуют кубическое ядро пируватдегидрогеназного комплекса, в котором 8 гомотримеров Е2 расположены в вершинах кубического ядра частицы.
Дигидролипоилдегидрогеназа (E3)
[ редактировать ]
Субъединица Е3, называемая ферментом дигидролипоилдегидрогеназой , характеризуется как гомодимерный белок, в котором два остатка цистеина , участвующие в дисульфидной связи , и кофактор FAD в активном центре облегчают его основную функцию в качестве окислительного катализатора. Один из примеров структуры E3, обнаруженной у Pseudomonas putida , сформирован таким образом, что каждая отдельная субъединица гомодимера содержит два связывающих домена, ответственных за связывание FAD и связывание NAD, а также центральный домен и интерфейсный домен. [6] [7]
Белок, связывающий дигидролипоилдегидрогеназу (E3BP)
[ редактировать ]Вспомогательным белком, уникальным для большинства эукариот, является белок, связывающий Е3 (E3BP), который служит для связывания субъединицы E3 с комплексом PDC. В случае человеческого E3BP гидрофобные остатки пролина и лейцина в BP взаимодействуют с поверхностным сайтом узнавания, образованным при связывании двух идентичных мономеров E3. [8]
Механизм
[ редактировать ]Эта статья нуждается в дополнительных цитатах для проверки . ( декабрь 2023 г. ) |
| Ферменты | Сокращение. | Кофакторы | # субъединицы прокариот | # субъединицы эукариот |
|---|---|---|---|---|
| пируватдегидрогеназа ( ЕС 1.2.4.1 ) | Е1 | ТПП (тиаминпирофосфат) , Мг 2+ | 32 | 30 |
| дигидролипоилтрансацетилаза ( ЕС 2.3.1.12 ) | Е2 | альфа- липоевая кислота (липоат) | 24 | 60 |
| дигидролипоилдегидрогеназа ( ЕС 1.8.1.4 ) | Е3 | ФАД | 16 | 12 |
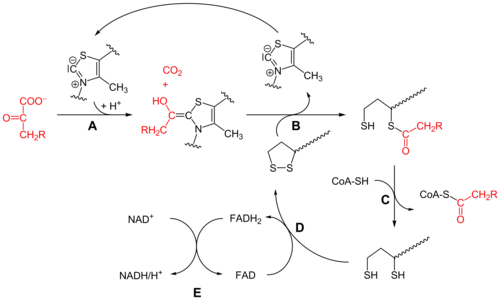
Пируватдегидрогеназа (E1)
[ редактировать ]Первоначально пируват и тиаминпирофосфат (TPP или витамин B 1 ) связываются субъединицами пируватдегидрогеназы . [1] Тиазолиевое цвиттер - кольцо TPP находится в ионной форме, а анионный углерод C2 осуществляет нуклеофильную атаку на карбонил C2 (кетон) пирувата. Полученное промежуточное соединение подвергается декарбоксилированию с образованием эквивалента ацил-аниона (см. Химию циангидрина или альдегид-дитиана умполунга , а также бензоиновую конденсацию ). Этот анион атакует S1 окисленного липоата, присоединенного к остатку лизина . В механизме, подобном раскрытию кольца S N 2 , S2 замещается в виде сульфидного или сульфгидрильного фрагмента. Последующий коллапс тетраэдрического промежуточного продукта выбрасывает тиазол, высвобождая кофактор TPP и образуя тиоацетат на S1 липоата. Процесс, катализируемый E1, является стадией, лимитирующей скорость всего пируватдегидрогеназного комплекса.
Дигидролипоилтрансацетилаза (E2)
[ редактировать ]В этот момент липоат-тиоэфирная функциональность перемещается в активный центр дигидролипоилтрансацетилазы (Е2), [1] где реакция трансацилирования переносит ацетил из «качающегося плеча» липоила в тиол А. кофермента При этом образуется ацетил-КоА , который высвобождается из ферментного комплекса и впоследствии входит в цикл лимонной кислоты . E2 также может быть известен как липоамидредуктаза-трансацетилаза.
Дигидролипоилдегидрогеназа (E3)
[ редактировать ]Дигидролипоат , ковалентно связанный с остатком лизина комплекса, затем переносится в активный центр дигидролипоилдегидрогеназы (Е3), [1] где он подвергается флавин -опосредованному окислению, аналогичному по химическому составу, например, тиоредоксинредуктазе. Во-первых, FAD окисляет дигидролипоат обратно в липоатное (дисульфидное) состояние покоя, образуя FADH 2 . Затем субстрат НАД + окисляет ФАДН 2 обратно в состояние покоя ФАД, образуя НАДН и H. + .
Структурные различия между видами
[ редактировать ]PDC представляет собой большой комплекс, состоящий из множества копий 3 или 4 субъединиц в зависимости от вида.
Грамотрицательные бактерии
[ редактировать ]У грамотрицательных бактерий, например Escherichia coli , PDC состоит из центрального кубического ядра, состоящего из 24 молекул дигидролипоилтрансацетилазы (Е2).До 16 гомодимеров пируватдегидрогеназы (E1) и 8 гомодимеров дигидролипоилдегидрогеназы (E3) связываются с 24 доменами связывания периферических субъединиц (PSBD) 24-мера E2.У Gammaproteobacteria специфичность PSBD к связыванию E1 или E3 определяется олигомерным состоянием PSBD. В каждом гомотримере E2 димеризуются два из трех PSBD. В то время как два гомодимера E1 кооперативно связывают димерный PSBD, оставшийся неспаренный PSBD специфически взаимодействует с одним гомодимером E3. Димеризация PSBD, таким образом, определяет субъединичный состав комплекса пируватдегидрогеназы при полном насыщении периферическими субъединицами E1 и E3, который имеет стехиометрию E1:E2:E3 (мономеры) = 32:24:16. [9]
Грамположительные бактерии и эукариоты
[ редактировать ]Напротив, у грамположительных бактерий (например, Bacillus stearothermophilus ) и эукариотов центральное ядро PDC содержит 60 молекул E2, организованных в икосаэдр. Это «ядро» субъединицы E2 координирует 30 субъединиц E1 и 12 копий E3.
Эукариоты также содержат 12 копий дополнительного корового белка, E3-связывающего белка (E3BP), который связывает субъединицы E3 с ядром E2. [10] Точное расположение E3BP не совсем ясно. Криоэлектронная микроскопия установила, что E3BP связывается с каждой из икосаэдрических граней дрожжей. [11] Однако было высказано предположение, что он заменяет эквивалентное количество молекул E2 в ядре бычьего PDC.
До 60 молекул E1 или E3 могут связываться с ядром E2 грамположительных бактерий - связывание является взаимоисключающим. У эукариот E1 специфически связывается с E2, а E3 — с E3BP. Считается, что присутствует до 30 ферментов E1 и 6 E3, хотя точное количество молекул может варьироваться in vivo и часто отражает метаболические потребности рассматриваемой ткани.
Регулирование
[ редактировать ]Пируватдегидрогеназа ингибируется, когда увеличивается одно или несколько из трех следующих соотношений: АТФ / АДФ , НАДН /НАД. + и ацетил-КоА / КоА . [ нужна ссылка ]
У эукариот PDC жестко регулируется собственной специфической киназой пируватдегидрогеназы (PDK) и фосфатазой пируватдегидрогеназы (PDP), деактивируя и активируя ее соответственно. [12]
- PDK фосфорилирует три специфических остатка серина на E1 с разным сродством. Фосфорилирование любого из них (с использованием АТФ ) делает E1 (и, как следствие, весь комплекс) неактивным. [12]
- Дефосфорилирование E1 с помощью PDP восстанавливает комплексную активность. [12]
Продукты реакции действуют как аллостерические ингибиторы ПДК, поскольку активируют ПДК. Субстраты, в свою очередь, ингибируют PDK, реактивируя PDC.
Во время голодания количество PDK увеличивается в большинстве тканей, включая скелетные мышцы , за счет увеличения транскрипции генов . В этих же условиях количество ПДП уменьшается. В результате ингибирование PDC предотвращает катаболизм глюкозы и предшественников глюконеогенеза в мышцах и других тканях . Метаболизм смещается в сторону утилизации жира , в то время как распад мышечного белка для обеспечения предшественников глюконеогенеза сводится к минимуму, а доступная глюкоза сохраняется для использования мозгом . [ нужна ссылка ]
Ионы кальция играют роль в регуляции PDC в мышечной ткани, поскольку они активируют PDP, стимулируя гликолиз при его высвобождении в цитозоль – во время мышечного сокращения . Некоторые продукты этой транскрипции выделяют H2 в мышцы. Это может привести к распаду ионов кальция с течением времени. [ нужна ссылка ]
Локализация декарбоксилирования пирувата
[ редактировать ]В эукариотических клетках декарбоксилирование пирувата происходит внутри митохондриального матрикса после транспорта субстрата пирувата из цитозоля . Транспорт пирувата в митохондрии осуществляется посредством транспортного белка пируват-транслоказы. Пируваттранслоказа переносит пируват симпортным способом с протоном (через внутреннюю митохондриальную мембрану), что можно рассматривать как форму вторичного активного транспорта, но для использования дескриптора «вторичный активный транспорт» может потребоваться дальнейшее подтверждение/подтверждение. здесь (Примечание: метод транспортировки пирувата посредством пируваттранслоказы, по-видимому, связан с протонным градиентом согласно S. Papa et al., 1971, что, по-видимому, соответствует по определению вторичному активному транспорту). [13]
Альтернативные источники говорят, что «транспорт пирувата через внешнюю митохондриальную мембрану, по-видимому, легко осуществляется через крупные неселективные каналы, такие как потенциал-зависимые анионные каналы , которые обеспечивают пассивную диффузию», а транспорт через внутреннюю митохондриальную мембрану опосредуется митохондриальным переносчиком пирувата 1 ( MPC1) и митохондриальный пируватный носитель 2 (MPC2). [14]
При попадании в митохондриальный матрикс пируват декарбоксилируется с образованием ацетил-КоА (а также диоксида углерода и НАДН). Эта необратимая реакция захватывает ацетил-КоА внутри митохондрий (ацетил-КоА может транспортироваться из митохондриального матрикса только в условиях высокого содержания оксалоацетата через цитратный челнок, промежуточный продукт ТСА, которого обычно мало). Диоксид углерода, образующийся в результате этой реакции, неполярен и мал и может диффундировать из митохондрий и из клетки. [ нужна ссылка ]
У прокариот , не имеющих митохондрий, эта реакция либо осуществляется в цитозоле, либо не осуществляется вообще. [ нужна ссылка ]
Эволюционная история
[ редактировать ]Было обнаружено, что фермент пируватдегидрогеназа, обнаруженный в митохондриях эукариотических клеток, очень похож на фермент Geobacillus stearothermophilus , который является разновидностью грамположительных бактерий . Несмотря на сходство комплекса пируватдегидрогеназы с грамположительными бактериями, сходства с таковым у грамотрицательных бактерий мало . Сходство четвертичных структур между пируватдегидрогеназой и ферментами грамположительных бактерий указывает на общую эволюционную историю, которая отличается от истории эволюции соответствующих ферментов, обнаруженных у грамотрицательных бактерий. В результате эндосимбиоза пируватдегидрогеназа, обнаруженная в митохондриях эукариот, указывает на наследственные связи, восходящие к грамположительным бактериям. [15]
Комплексы пируватдегидрогеназы имеют много общего с дегидрогеназой 2-оксокислот с разветвленной цепью (BCOADH), особенно в их субстратной специфичности в отношении альфа-кетокислот. В частности, BCOADH катализирует распад аминокислот, и эти ферменты должны были преобладать в те периоды, когда на доисторической Земле доминировала среда, богатая аминокислотами. Субъединица E2 пируватдегидрогеназы произошла от гена E2, обнаруженного в BCOADH, в то время как оба фермента содержат идентичные субъединицы E3 из-за присутствия только одного гена E3. Поскольку субъединицы E1 обладают отличительной специфичностью к конкретным субстратам, субъединицы E1 пируватдегидрогеназы и BCOADH различаются, но имеют генетическое сходство. Грамположительные бактерии и цианобактерии, которые позже дали начало митохондриям и хлоропластам, обнаруженным в эукариотических клетках, сохранили субъединицы E1, которые генетически связаны с субъединицами, обнаруженными в ферментах BCOADH. [16] [17]
Клиническая значимость
[ редактировать ]Дефицит пируватдегидрогеназы (PDCD) может быть результатом мутаций любого из ферментов или кофакторов, используемых для построения комплекса. Его основным клиническим признаком является лактоацидоз . [18] Такие мутации PDCD, приводящие к последующему дефициту продукции NAD и FAD, препятствуют процессам окислительного фосфорилирования, которые являются ключевыми в аэробном дыхании. Таким образом, вместо этого ацетил-КоА восстанавливается посредством анаэробных механизмов до других молекул, таких как лактат, что приводит к избытку лактата в организме и связанным с ним неврологическим патологиям. [19]
Хотя дефицит пируватдегидрогеназы встречается редко, существует множество различных генов, мутировавших или нефункциональных, которые могут вызвать этот дефицит. Во-первых, субъединица Е1 пируватдегидрогеназы содержит четыре различных субъединицы: две альфа-субъединицы, обозначаемые как Е1-альфа, и две бета-субъединицы, обозначаемые как Е1-бета. Ген PDHA1, обнаруженный в субъединицах E1-альфа, при мутации вызывает 80% случаев дефицита пируватдегидрогеназы, поскольку эта мутация сокращает белок E1-альфа. Снижение функционального уровня E1-альфа препятствует достаточному связыванию пируватдегидрогеназы с пируватом, тем самым снижая активность всего комплекса. [20] Мутация гена PDHB, обнаруженного в бета-субъединице E1 комплекса, также приводит к дефициту пируватдегидрогеназы. [21] Аналогичным образом, были прослежены мутации, обнаруженные в других субъединицах комплекса, таких как ген DLAT, обнаруженный в субъединице E2, ген PDHX, обнаруженный в субъединице E3, а также мутация гена фосфатазы пируватдегидрогеназы, известного как PDP1. обратно к дефициту пируватдегидрогеназы, а их конкретный вклад в болезненное состояние неизвестен. [22] [23] [24]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: Перейти обратно: а б с д ДеБросс С.Д., Керр Д.С. (01 января 2016 г.), Сането Р.П., Парих С., Коэн Б.Х. (ред.), «Глава 12 - Дефицит комплекса пируватдегидрогеназы» , Тематические исследования митохондрий , Бостон: Academic Press, стр. 93–101 , doi : 10.1016/b978-0-12-800877-5.00012-7 , ISBN 978-0-12-800877-5 , получено 16 ноября 2020 г.
- ^ Дж. М. Берг, Дж. Л. Тимочко, Л. Страйер (2007). Биохимия (6-е изд.). Фриман. ISBN 978-0-7167-8724-2 .
- ^ Сгриньяни Дж., Чен Дж., Алимонти А. (2018). «Как фосфорилирование влияет на пируватдегидрогеназу субъединицы E1: вычислительное исследование» . Научные отчеты . 8 (14683): 14683. Бибкод : 2018NatSR...814683S . дои : 10.1038/s41598-018-33048-z . ПМК 6168537 . ПМИД 30279533 . S2CID 52910721 .
- ^ [Идентификатор PBD: 2QTC] Кале С., Арджунан П., Фьюри В., Джордан Ф. (2007). «Динамическая петля в активном центре КОМПЛЕКСА E1 пируватдегидрогеназы Escherichia coli модулирует использование СУБСТРАТА и ХИМИЧЕСКУЮ связь с компонентом E2» . Журнал биологической химии . 282 (38): 28106–28116. дои : 10.1074/jbc.m704326200 . ПМИД 17635929 . S2CID 25199383 .
- ^ Патель М.С., Немерия Н.С., Фьюри В., Джордан Ф. (2014). «Комплексы пируватдегидрогеназы: структурная функция и регуляция» . Журнал биологической химии . 289 (24): 16615–16623. дои : 10.1074/jbc.R114.563148 . ПМК 4059105 . ПМИД 24798336 .
- ^ Биллгрен Э.С., Чикчилло Р.М., Несбитт Н.М., Букер С.Дж. (2010). «Биосинтез и энзимология липоевой кислоты». Комплексные натуральные продукты . 2 (7): 181–212. дои : 10.1016/B978-008045382-8.00137-4 .
- ^ [ID PDB: 1LVL] Маттевиа А., Обмолова Г., Сокач Дж.Р., Бетцель С., Хол В.Г. (1992). «Уточненная кристаллическая СТРУКТУРА псевдомонас Putida ЛИПОАМИДДЕГИДРОГЕНАЗЫ в комплексе с НАД + при разрешении 2,45 Å». Белки: структура, функции и генетика . 13 (4): 336–351. дои : 10.1002/прот.340130406 . ПМИД 1325638 . S2CID 23288363 .
- ^ Чишак Э.М., Макал А., Хонг Ю.С., Веттаиккоруманкаув АК, Корочкина Л.Г., Патель М.С. (2006). «Как белок, связывающий дигидролипоамиддегидрогеназу, связывает дигидролипоамиддегидрогеназу в комплексе пируватдегидрогеназы человека» . Журнал биологической химии . 281 (1): 648–655. дои : 10.1074/jbc.m507850200 . ПМИД 16263718 . S2CID 26797600 .
- ^ Мейнхольд С., Зданович Р., Гизе К., Глокшубер Р. (2024). «Димеризация домена 5-кДа определяет архитектуру комплекса гаммапротеобактериальной пируватдегидрогеназы 5-МДа» . наук. Адв . 10 (6): eadj6358. Бибкод : 2024SciA...10J6358M . дои : 10.1126/sciadv.adj6358 . ПМЦ 10849603 . ПМИД 38324697 . [1]
- ^ Бротигам К.А., Винн Р.М., Чуанг Дж.Л., Мачиус М., Томчик Д.Р., Чуанг Д.Т. (2006). «Структурное понимание взаимодействий между дигидролипоамиддегидрогеназой (E3) и E3-связывающим белком комплекса пируватдегидрогеназы человека» . Структура . 14 (3): 611–621. doi : 10.1016/j.str.2006.01.001 . ПМЦ 2879633 . ПМИД 16442803 .
- ^ Ступс, Дж.К., Ченг, Р.Х., Язди, М.А., Маенг, С.Ю., Шретер, Дж.П., Клюппельберг, У., Колодзей, С.Дж., Бейкер, Т.С., Рид, Л.Дж. (1997) Об уникальной структурной организации пирувата Saccharomyces cerevisiae дегидрогеназный комплекс. Ж. Биол. хим. 272, 5757-5764.
- ^ Jump up to: Перейти обратно: а б с Пелли Дж.В. (01.01.2012), Пелли Дж.В. (редактор), «6 - Гликолиз и окисление пирувата» , Комплексный обзор биохимии Elsevier (второе издание) , Филадельфия: WB Saunders, стр. 49–55, doi : 10.1016/ b978-0-323-07446-9.00006-4 , ISBN 978-0-323-07446-9 , получено 16 ноября 2020 г.
- ^ Папа С. (30 января 1971 г.). «Транспорт пирувата в митохондриях печени крысы» . ФЭБС Летт . 12 (5): 285–288. дои : 10.1016/0014-5793(71)80200-4 . ПМИД 11945601 .
- ^ Раттер Дж. (23 января 2013 г.). «Долгий и извилистый путь к митохондриальному переносчику пирувата» . Рак и обмен веществ . 1 (1): 6. дои : 10.1186/2049-3002-1-6 . ПМЦ 3834494 . ПМИД 24280073 .
- ^ Хендерсон CE, Перхэм Р.Н., Финч Дж.Т. (май 1979 г.). «Структура и симметрия мультиферментного комплекса пируватдегидрогеназы B. stearothermophilus и значение для эволюции эукариот» . Клетка . 17 (1): 85–93. дои : 10.1016/0092-8674(79)90297-6 . ISSN 0092-8674 . ПМИД 455461 . S2CID 35282258 .
- ^ Шрайнер М.Э., Фиур Д., Холатко Дж., Патек М., Эйкманнс Б.Дж. (01 сентября 2005 г.). «Фермент E1 комплекса пируватдегидрогеназы в Corynebacterium Glutamicum: молекулярный анализ гена и филогенетические аспекты» . Журнал бактериологии . 187 (17): 6005–6018. дои : 10.1128/jb.187.17.6005-6018.2005 . ISSN 0021-9193 . ПМК 1196148 . ПМИД 16109942 .
- ^ Шнарренбергер К., Мартин В. (1 февраля 2002 г.). «Эволюция ферментов цикла лимонной кислоты и глиоксилатного цикла высших растений» . Европейский журнал биохимии . 269 (3): 868–883. дои : 10.1046/j.0014-2956.2001.02722.x . ISSN 0014-2956 . ПМИД 11846788 .
- ^ «Дефицит пируватдегидрогеназы» . Домашний справочник по генетике . Проверено 17 марта 2013 г.
- ^ Гупта Н., Ратледж С. (2019). «Дефицит комплекса пируватдегидрогеназы: необычная причина рецидивирующего лактоацидоза в педиатрическом отделении интенсивной терапии» . Журнал медицины критических состояний . 5 (2): 71–75. doi : 10.2478/jccm-2019-0012 . ПМК 6534940 . ПМИД 31161145 .
- ^ Лиссенс В., Де Мейрлейр Л., Сенека С., Либерс И., Браун Г.К., Браун Р.М., Ито М., Найто Э., Курода Ю., Керр Д.С., Векслер И.Д. (март 2000 г.). «Мутации в гене субъединицы Х-связанной пируватдегидрогеназы (E1) ? (PDHA1) у пациентов с дефицитом комплекса пируватдегидрогеназы» . Человеческая мутация . 15 (3): 209–219. doi : 10.1002/(sici)1098-1004(200003)15:3<209::aid-humu1>3.0.co;2-k . ISSN 1059-7794 . ПМИД 10679936 . S2CID 29926332 .
- ^ Окаджима К., Корочкина Л., Прасад С., Рупар Т., Филлипс III Дж., Фичичиоглу С., Хертекант Дж., Патель М., Керр Д. (апрель 2008 г.). «Мутации гена субъединицы E1β (PDHB) в четырех семьях с дефицитом пируватдегидрогеназы» . Молекулярная генетика и обмен веществ . 93 (4): 371–380. дои : 10.1016/j.ymgme.2007.10.135 . ISSN 1096-7192 . ПМИД 18164639 .
- ^ Руководитель РА, Браун Р.М., Золкипли З., Шахдадпури Р., Кинг М.Д., Клейтон П.Т., Браун Г.К. (27 июля 2005 г.). «Клинический и генетический спектр дефицита пируватдегидрогеназы: дефицит дигидролипоамидацетилтрансферазы (Е2)» . Анналы неврологии . 58 (2): 234–241. дои : 10.1002/ана.20550 . ISSN 0364-5134 . ПМИД 16049940 . S2CID 38264402 .
- ^ Павлу-Перейра Х, Силва МЮ, Флориндо К, Секейра С, Феррейра АС, Дуарте С, Родригеш А.Л., Жанейро П., Оливейра А, Гомес Д, Бандейра А, Мартинс Е, Гомес Р., Соареш С., Таварес де Алмейда I, Висенте Дж. Б., Ривера I (декабрь 2020 г.). «Дефицит комплекса пируватдегидрогеназы: обновление клинической, метаболической и мутационной картины в когорте португальских пациентов» . Сиротский журнал редких заболеваний . 15 (1): 298. дои : 10.1186/s13023-020-01586-3 . ПМЦ 7579914 . ПМИД 33092611 .
- ^ Хо Х.Дж., Ким Х.К., Юм Дж.Б., Чо С.В., Сонг И.С., Ли С.И., Ко Т.Х., Ким Н., Ко КС, Ри Б.Д., Хан Дж. (август 2016 г.). «Митохондриальная пируватдегидрогеназа-фосфатаза 1 регулирует раннюю дифференцировку кардиомиоцитов из эмбриональных стволовых клеток мыши» . Экспериментальная и молекулярная медицина . 48 (8): е254. дои : 10.1038/эмм.2016.70 . ISSN 2092-6413 . ПМК 5007642 . ПМИД 27538372 .
Внешние ссылки
[ редактировать ]- https://web.archive.org/web/20070405211049/http://www.dentistry.leeds.ac.uk/biochem/MBWeb/mb1/part2/krebs.htm#animat1 - анимация общего механизма ПДК (ссылка вверху справа) в Университете Лидса
- Комплекс пируват+дегидрогеназа+ в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
3D-структуры
[ редактировать ]- Чжоу Х., Маккарти Б., О'Коннор М., Рид Дж., Ступс К. (декабрь 2001 г.). «Замечательная структурная и функциональная организация эукариотических комплексов пируватдегидрогеназы» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (26): 14802–14807. Бибкод : 2001PNAS...9814802Z . дои : 10.1073/pnas.011597698 . ISSN 0027-8424 . ПМК 64939 . ПМИД 11752427 . , бычьих почек комплекс пируватдегидрогеназы
- Ю Х, Хиромаса Ю, Цен Х, Ступс К, Рош Э, Чжоу Х (январь 2008 г.). «Структуры ядер комплекса пируватдегидрогеназы человека: высококонсервативный каталитический центр с гибкими N-концевыми доменами» . Структура . 16 (1): 104–114. дои : 10.1016/j.str.2007.10.024 . ISSN 0969-2126 . ПМК 4807695 . ПМИД 18184588 . , полноразмерные и усеченные ядра E2 (tE2) человека PDC, экспрессированные в E. coli









