Пролин
| |||
| |||
| Имена | |||
|---|---|---|---|
| Название ИЮПАК Пролин | |||
| Систематическое название ИЮПАК Пирролидин-2-карбоновая кислота [1] | |||
| Идентификаторы | |||
3D model ( JSmol ) | |||
| 80812 | |||
| ЧЭБИ |
| ||
| ЧЕМБЛ |
| ||
| ХимическийПаук | |||
| Лекарственный Банк |
| ||
| Информационная карта ECHA | 100.009.264 | ||
| Номер ЕС |
| ||
| 26927 | |||
| КЕГГ |
| ||
| МеШ | Пролин | ||
ПабХим CID | |||
| номер РТЭКС |
| ||
| НЕКОТОРЫЙ |
| ||
Панель управления CompTox ( EPA ) | |||
| Характеристики | |||
| C5H9NOC5H9NO2 | |||
| Молярная масса | 115.132 g·mol −1 | ||
| Появление | Прозрачные кристаллы | ||
| Температура плавления | От 205 до 228 ° C (от 401 до 442 ° F; от 478 до 501 К) (разлагается) | ||
| Растворимость | 1,5 г/100 г этанола 19 град.С [2] | ||
| войти P | -0.06 | ||
| Кислотность ( pKa ) | 1,99 (карбоксил), 10,96 (амино) [3] | ||
| Страница дополнительных данных | |||
| Пролин (страница данных) | |||
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |||




Пролин (символ Pro или P ) [4] представляет собой органическую кислоту, относящуюся к протеиногенным аминокислотам (используемым в биосинтезе белков ), хотя она не содержит аминогруппы -NH.
2 , а скорее является вторичным амином . Азот вторичного амина находится в протонированной форме (NH 2 + ) в биологических условиях, а карбоксильная группа находится в депротонированном состоянии -COO − форма. «Боковая цепь» α-углерода соединяется с азотом, образуя пирролидиновую петлю, классифицируя ее как алифатическую аминокислоту . Он не является незаменимым для человека, то есть организм может синтезировать его из незаменимой аминокислоты L – глутамата . Он кодируется всеми кодонами, начинающимися с CC (CCU, CCC, CCA и CCG).
Пролин — единственная протеиногенная вторичная аминокислота , которая является вторичным амином, поскольку атом азота присоединен как к α-углероду, так и к цепи из трех атомов углерода, которые вместе образуют пятичленное кольцо.
История и этимология [ править ]
Пролин был впервые выделен в 1900 году Рихардом Вильштеттером , который получил аминокислоту при изучении N -метилпролина и синтезировал пролин реакцией натриевой соли диэтилмалоната с 1,3-дибромпропаном . В следующем году Эмиль Фишер выделил пролин из казеина и продуктов распада γ-фталимидопропилмалонового эфира. [5] и опубликовал синтез пролина из фталимидно-пропилмалонового эфира. [6]
Название пролин происходит от пирролидина , одного из его компонентов. [7]
Биосинтез [ править ]
Пролин получают биосинтетически из аминокислоты L - глутамата . Глутамат-5-полуальдегид сначала образуется глутамат-5-киназой (АТФ-зависимой) и глутамат-5-полуальдегиддегидрогеназой (которая требует НАДН или НАДФН). Затем он может либо спонтанно циклизоваться с образованием 1-пирролин-5-карбоновой кислоты , которая восстанавливается до пролина пирролин-5-карбоксилатредуктазой (с использованием НАДН или НАДФН), либо превращается в орнитин с помощью орнитинаминотрансферазы с последующей циклизацией орнитинциклодеаминазой . с образованием пролина. [8]

Биологическая активность [ править ]
L Было обнаружено, что -пролин действует как слабый агонист глициновых рецепторов , а также NMDA и не-NMDA ( AMPA / каинат ) ионотропных глутаматных рецепторов . [9] [10] [11] Было высказано предположение, что это потенциальный эндогенный экситотоксин . [9] [10] [11] У растений накопление пролина является обычной физиологической реакцией на различные стрессы, но также является частью программы развития генеративных тканей (например, пыльцы ). [12] [13] [14] [15]
Диета, богатая пролином, была связана с повышенным риском депрессии у людей в исследовании 2022 года, которое было проверено в ходе ограниченных доклинических испытаний на людях и, прежде всего, на других организмах. Результаты были значительными у других организмов. [16]
Свойства структуры белка [ править ]
Отличительная циклическая структура боковой цепи пролина придает пролину исключительную конформационную жесткость по сравнению с другими аминокислотами. Это также влияет на скорость образования пептидных связей между пролином и другими аминокислотами. Когда пролин связан в виде амида пептидной связью, его азот не связан ни с каким водородом, то есть он не может действовать как донор водородной связи , но может быть акцептором водородной связи.
Образование пептидной связи с поступающей про-тРНК Про в рибосоме происходит значительно медленнее, чем в любых других тРНК, что является общей особенностью N -алкиламинокислот. [17] Образование пептидной связи между входящей тРНК и цепью, оканчивающейся пролином, также происходит медленно; при этом создание пролин-пролиновых связей происходит медленнее всего. [18]
Исключительная конформационная жесткость пролина влияет на вторичную структуру белков вблизи остатка пролина и может быть причиной более высокой распространенности пролина в белках термофильных организмов. Вторичную структуру белка можно описать с помощью двугранных углов φ, ψ и ω основной цепи белка. Циклическая структура боковой цепи пролина фиксирует угол φ примерно на уровне -65 °. [19]
Пролин действует как структурный разрушитель в середине регулярных элементов вторичной структуры, таких как альфа-спирали и бета-листы ; однако пролин обычно встречается в виде первого остатка альфа-спирали , а также в краевых нитях бета-листов . Пролин также часто встречается в витках (еще один вид вторичной структуры) и способствует образованию бета-витков. Это может объяснить тот любопытный факт, что пролин обычно подвергается воздействию растворителей, несмотря на то, что он имеет полностью алифатическую боковую цепь.
Несколько пролинов и/или подряд могут образовывать полипролиновую спираль , преобладающую вторичную структуру коллагена гидроксипролинов . Гидроксилирование пролилгидроксилазой пролина фтор (или другими добавками электроноакцепторных заместителей, таких как ) значительно увеличивает конформационную стабильность коллагена . [20] Следовательно, гидроксилирование пролина является важнейшим биохимическим процессом для поддержания соединительной ткани высших организмов. Тяжелые заболевания, такие как цинга, могут возникнуть в результате дефектов этого гидроксилирования, например, мутаций фермента пролилгидроксилазы или отсутствия необходимого кофактора аскорбата (витамина С) .
Цис - транс- изомеризация [ править ]
Пептидные связи с пролином и другими N -замещенными аминокислотами (такими как саркозин ) способны заселять как цис- , так и транс- изомеры. Большинство пептидных связей в подавляющем большинстве принимают транс- изомер (обычно 99,9% в ненапряженных условиях), главным образом потому, что амидный водород ( транс -изомер) оказывает меньшее стерическое отталкивание к предыдущему атому Cα, чем следующий цис атом Cα ( - изомер). Напротив, цис- и транс -изомеры пептидной связи X-Pro (где X представляет собой любую аминокислоту) испытывают стерические столкновения с соседними заменами и имеют гораздо меньшую разницу в энергии. Следовательно, доля пептидных связей X-Pro в цис- изомере в ненапряженных условиях значительно повышена, при этом доля цис-цис обычно находится в диапазоне 3-10%. [21] Однако эти значения зависят от предыдущей аминокислоты: Gly [22] и ароматный [23] остатки, дающие повышенную долю цис- изомера. цис- фракции до 40%. Для пептидных связей ароматика-пролин идентифицированы [24]
С кинетической точки зрения цис - транс пролина -изомеризация представляет собой очень медленный процесс, который может препятствовать прогрессу сворачивания белка за счет захвата одного или нескольких остатков пролина, имеющих решающее значение для сворачивания, в ненативном изомере, особенно когда нативный белок требует цис- изомера . Это связано с тем, что остатки пролина синтезируются исключительно в рибосоме в форме транс- изомера. Все организмы обладают пролилизомеразой, ферментами катализирующими эту изомеризацию, а некоторые бактерии имеют специализированные пролилизомеразы, связанные с рибосомой. Однако не все пролины необходимы для сворачивания, и сворачивание белка может происходить с нормальной скоростью, несмотря на наличие ненативных конформеров многих пептидных связей X-Pro.
Использует [ править ]
Пролин и его производные часто используются в качестве асимметричных катализаторов в реакциях органокатализа пролина . Яркими примерами являются восстановление CBS и альдольная конденсация , катализируемая пролином .
В пивоварении белки, богатые пролином, соединяются с полифенолами, образуя дымку (мутность). [25]
L -Пролин является осмопротектором и поэтому используется во многих фармацевтических и биотехнологических приложениях.
используемая Ростовая среда, в культуре тканей растений, может быть дополнена пролином. Это может ускорить рост, возможно, потому, что помогает растению переносить стрессы, связанные с тканевой культурой. [26] [ нужен лучший источник ] О роли пролина в реакции растений на стресс см. § Биологическая активность .
Специальности [ править ]
Пролин – одна из двух аминокислот, которые не следуют типичному графику Рамачандрана , наряду с глицином . Из-за образования кольца, связанного с бета-углеродом, углы ψ и φ вокруг пептидной связи имеют меньше допустимых градусов вращения. В результате он часто встречается в «витках» белков, поскольку его свободная энтропия (Δ S ) не так велика, как у других аминокислот, и, следовательно, в свернутой форме по сравнению с развернутой формой изменение энтропии меньше. Кроме того, пролин редко встречается в α- и β-структурах, поскольку он снижает стабильность таких структур, поскольку α-азот его боковой цепи может образовывать только одну азотистую связь.
Кроме того, пролин — единственная аминокислота, которая не окрашивается в красно-фиолетовый цвет при распылении нингидрина для использования в хроматографии . Вместо этого пролин дает оранжево-желтый цвет.
Синтез [ править ]
Рацемический пролин можно синтезировать из диэтилмалоната и акрилонитрила : [27]
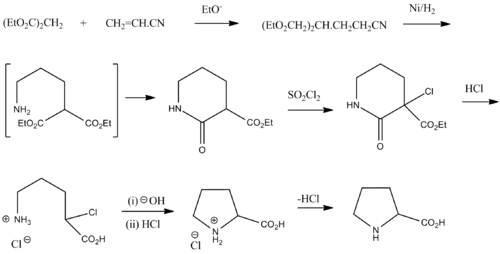
См. также [ править ]
Ссылки [ править ]
- ^ «Пролин» . ПабХим . Национальная медицинская библиотека США. Архивировано из оригинала 16 января 2014 года . Проверено 8 мая 2018 г.
- ^ Белитц Х.Д., Грош В., Шиберле П. (15 января 2009 г.). Пищевая химия . Спрингер. п. 15. ISBN 978-3-540-69933-0 . Архивировано из оригинала 15 мая 2016 г.
- ^ Нельсон Д.Л., Кокс М.М. Принципы биохимии . Нью-Йорк: WH Freeman and Company.
- ^ «Номенклатура и символика аминокислот и пептидов» . Совместная комиссия IUPAC-IUB по биохимической номенклатуре. 1983. Архивировано из оригинала 9 октября 2008 года . Проверено 5 марта 2018 г.
- ^ Плиммер Р.Х. (1912) [1908], Плиммер Р.Х., Хопкинс Ф.Г. (ред.), Химический состав белков , Монографии по биохимии, вып. Часть I. Анализ (2-е изд.), Лондон: Longmans, Green and Co., стр. 130 , получено 20 сентября 2010 г.
- ^ «Пролин» . Руководство по аминокислотам . Архивировано из оригинала 27 ноября 2015 г.
- ^ «Пролин» . Словарь английского языка американского наследия, 4-е издание . Архивировано из оригинала 15 сентября 2015 г. Проверено 6 декабря 2015 г.
- ^ Ленинджер А.Л., Нельсон Д.Л., Кокс М.М. (2000). Принципы биохимии (3-е изд.). Нью-Йорк: WH Freeman. ISBN 1-57259-153-6 . .
- ^ Jump up to: Перейти обратно: а б Справочник по ионным каналам: внеклеточные лиганд-управляемые каналы . Академическая пресса. 16 ноября 1995 г. с. 126. ИСБН 978-0-08-053519-7 . Архивировано из оригинала 26 апреля 2016 года.
- ^ Jump up to: Перейти обратно: а б Хензи В., Райхлинг Д.Б., Хелм С.В., Макдермотт AB (апрель 1992 г.). «L-пролин активирует рецепторы глутамата и глицина в культивируемых нейронах спинного рога крысы» . Молекулярная фармакология . 41 (4): 793–801. ПМИД 1349155 . [ постоянная мертвая ссылка ]
- ^ Jump up to: Перейти обратно: а б Арслан О.Э. (7 августа 2014 г.). Нейроанатомические основы клинической неврологии (Второе изд.). ЦРК Пресс. п. 309. ИСБН 978-1-4398-4833-3 . Архивировано из оригинала 14 мая 2016 года.
- ^ Вербрюгген Н., Херманс К. (ноябрь 2008 г.). «Накопление пролина в растениях: обзор». Аминокислоты . 35 (4): 753–759. дои : 10.1007/s00726-008-0061-6 . ПМИД 18379856 . S2CID 21788988 .
- ^ Шреста А., Фендель А., Нгуен Т.Х., Адебабай А., Куллик А.С., Бенндорф Дж. и др. (сентябрь 2022 г.). «Природное разнообразие раскрывает регуляцию P5CS1 и ее роль в устойчивости к засухе и устойчивости урожайности ячменя» . Растение, клетка и окружающая среда . 45 (12): 3523–3536. дои : 10.1111/шт.14445 . ПМИД 36130879 . S2CID 252438394 .
- ^ Шреста А., Каджо Д.К., Камруззаман М., Сиддик С., Фиорани Ф., Леон Дж., Наз А.А. (июнь 2021 г.). «Факторы транскрипции, связывающие элементы, чувствительные к абсцизовой кислоте, способствуют синтезу пролина и адаптации к стрессу у арабидопсиса». Журнал физиологии растений . 261 : 153414. doi : 10.1016/j.jplph.2021.153414 . ПМИД 33895677 . S2CID 233397785 .
- ^ Музаммил С., Шреста А., Дадшани С., Пиллен К., Сиддик С., Леон Дж., Наз А.А. (октябрь 2018 г.). «Наследственный аллель пирролин-5-карбоксилатсинтазы1 способствует накоплению пролина и адаптации к засухе у культивируемого ячменя» . Физиология растений . 178 (2): 771–782. дои : 10.1104/стр.18.00169 . ПМК 6181029 . ПМИД 30131422 .
- ^ Майнерис-Перксакс Х., Кастельс-Нобау А., Арнориага-Родригес М., Мартин М., де ла Вега-Корреа Л., Сапата С. и др. (май 2022 г.). «Изменения микробиоты в метаболизме пролина влияют на депрессию» . Клеточный метаболизм . 34 (5): 681–701.e10. doi : 10.1016/j.cmet.2022.04.001 . hdl : 10230/53513 . ПМИД 35508109 . S2CID 248528026 .
- ^ Павлов М.Ю., Уоттс Р.Э., Тан З., Корниш В.В., Эренберг М., Форстер А.С. (январь 2009 г.). «Медленное образование пептидной связи пролином и другими N -алкиламинокислотами при трансляции» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (1): 50–54. Бибкод : 2009ПНАС..106...50П . дои : 10.1073/pnas.0809211106 . ПМК 2629218 . ПМИД 19104062 . .
- ^ Бускирк А.Р., Грин Р. (январь 2013 г.). «Биохимия. Преодоление полипролиновых пауз» . Наука . 339 (6115): 38–39. Бибкод : 2013Sci...339...38B . дои : 10.1126/science.1233338 . ПМЦ 3955122 . ПМИД 23288527 .
- ^ Моррис А.Л., Макартур М.В., Хатчинсон Э.Г., Торнтон Дж.М. (апрель 1992 г.). «Стереохимическое качество координат белковой структуры». Белки . 12 (4): 345–364. дои : 10.1002/прот.340120407 . ПМИД 1579569 . S2CID 940786 .
- ^ Шпак П (2011). «Химия и ультраструктура рыбьих костей: значение для тафономии и анализа стабильных изотопов» . Журнал археологической науки . 38 (12): 3358–3372. дои : 10.1016/j.jas.2011.07.022 . Архивировано из оригинала 18 января 2012 г.
- ^ Олдерсон Т.Р., Ли Дж.Х., Шарлье С., Инь Дж., Бакс А. (январь 2018 г.). «Склонность к образованию цис -пролина в развернутых белках» . ХимБиоХим . 19 (1): 37–42. дои : 10.1002/cbic.201700548 . ПМЦ 5977977 . ПМИД 29064600 .
- ^ Саркар С.К., Янг П.Е., Салливан С.Э., Торчиа Д.А. (август 1984 г.). «Обнаружение цис- и транс -пептидных связей X-Pro в белках методом ЯМР 13С: применение к коллагену» . Труды Национальной академии наук Соединенных Штатов Америки . 81 (15): 4800–4803. Бибкод : 1984PNAS...81.4800S . дои : 10.1073/pnas.81.15.4800 . ПМЦ 391578 . ПМИД 6589627 .
- ^ Томас К.М., Надутамби Д., Зондло, штат Нью-Джерси (февраль 2006 г.). -изомерии амидов «Электронный контроль цис - транс посредством ароматико-пролильного взаимодействия». Журнал Американского химического общества . 128 (7): 2216–2217. дои : 10.1021/ja057901y . ПМИД 16478167 .
- ^ Густафсон К.Л., Петрушка Н.К., Асимгил Х., Ли Х.В., Альбах С., Майкл А.К. и др. (май 2017 г.). «Медленное конформационное переключение в домене трансактивации BMAL1 модулирует циркадные ритмы» . Молекулярная клетка . 66 (4): 447–457.e7. doi : 10.1016/j.molcel.2017.04.011 . ПМЦ 5484534 . ПМИД 28506462 .
- ^ Зиберт К.Дж. «Дымка и пена» . Корнелл АгроТех . Архивировано из оригинала 11 июля 2010 г. Проверено 13 июля 2010 г. По состоянию на 12 июля 2010 г.
- ^ Пазуки А, Асгари Дж, Сохани ММ, Пессаракли М, Афлаки Ф (2015). «Влияние некоторых источников органического азота и антибиотиков на рост каллуса сортов риса Индика». Журнал питания растений . 38 (8): 1231–1240. дои : 10.1080/01904167.2014.983118 . S2CID 84495391 .
- ^ Фогель, Практическая органическая химия, 5-е издание.
Дальнейшее чтение [ править ]
- Бальбах Дж., Шмид FX (2000). «Изомеризация пролина и ее катализ при сворачивании белка». В боли Р.Х. (ред.). Механизмы сворачивания белка (2-е изд.). Издательство Оксфордского университета. стр. 212–249. ISBN 978-0-19-963788-1 . .
