Дегидроэпиандростерон
 | |
 | |
| Имена | |
|---|---|
| Название ИЮПАК 3β-Гидроксиандрост-5-ен-17-он | |
| Систематическое название ИЮПАК (3a S ,3b R ,7 S ,9a R ,9b S ,11a S )-7-Гидрокси-9а,11а-диметил-2,3,3а,3b,4,6,7,8,9,9а, 9b,10,11,11a-тетрадекагидро-1H - циклопента[ а ]фенантрен-1-он | |
| Другие имена Андростенолон; Прастерон; Андрост-5-ен-3β-ол-17-он; 5,6-дидегидроэпиандростерон; [1] Дегидроизоэпиандростерон | |
| Идентификаторы | |
3D model ( JSmol ) | |
| ЧЭБИ | |
| ЧЕМБЛ | |
| ХимическийПаук | |
| Лекарственный Банк | |
| Информационная карта ECHA | 100.000.160 |
ПабХим CID | |
| НЕКОТОРЫЙ | |
Панель управления CompTox ( EPA ) | |
| Характеристики | |
| С 19 Н 28 О 2 | |
| Молярная масса | 288.424 g/mol |
| Температура плавления | 148.5 |
| Фармакология | |
| QA14AA07 ( ВОЗ ) G03EA03 ( ВОЗ ) (комбинация с эстрогеном ) | |
| Внутрь , вагинально ( вставка ), внутримышечная инъекция (в виде прастерона энантата ), инъекция (в виде прастерона сульфата натрия ) | |
| Фармакокинетика : | |
| 50% [2] | |
| Печеночный [2] | |
| ДГЭА : 25 минут [3] ДГЭА-С : 11 часов [3] | |
| Моча | |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |
Дегидроэпиандростерон ( ДГЭА ), также известный как андростенолон , является эндогенным предшественником стероидных гормонов . [4] Это один из наиболее распространенных стероидов, циркулирующих в организме человека. [5] ДГЭА вырабатывается в надпочечниках , [6] и половые железы мозг. [7] Он действует как метаболический промежуточный продукт в биосинтезе андрогенов других и эстрогенов половых стероидов как в половых железах, так и в различных тканях . [4] [8] [9] Тем не менее, DHEA также обладает множеством потенциальных биологических эффектов, связываясь с множеством ядерных и клеточных поверхностных рецепторов . [10] и действует как нейростероид и модулятор рецепторов нейротрофических факторов . [11]
В Соединенных Штатах ДГЭА продается как безрецептурная добавка , а также лекарство под названием прастерон .
Биологическая функция
[ редактировать ]В качестве андрогена
[ редактировать ]ДГЭА и другие андрогены надпочечников, такие как андростендион , хотя и относительно слабые андрогены, ответственны за андрогенные эффекты адренархе , такие как ранний рост волос на лобке и подмышками по-взрослому , запах тела , повышенная жирность волос и кожи и легкие прыщи . [12] [13] [14] ДГЭА усиливается локально за счет превращения в тестостерон и дигидротестостерон (ДГТ) в коже и волосяных фолликулах . [4] У женщин с синдромом полной нечувствительности к андрогенам (CAIS), которые имеют нефункциональный андрогенный рецептор (AR) и невосприимчивы к андрогенному воздействию DHEA и других андрогенов, отсутствуют или только редкие/скудные волосы на лобке и подмышках, а также волосы на теле в целом. , демонстрируя роль ДГЭА и других андрогенов в развитии волос на теле как при адренархе, так и при пубархе . [15] [16] [17] [18]
В качестве эстрогена
[ редактировать ]ДГЭА – слабый эстроген . [4] [10] [19] Кроме того, он трансформируется в мощные эстрогены, такие как эстрадиол, в определенных тканях, таких как влагалище , и тем самым оказывает эстрогенное действие на такие ткани. [4]
Как нейростероид
[ редактировать ]Как нейростероид и нейротрофин , ДГЭА оказывает важное воздействие на центральную нервную систему . [20] [21] [22]
Биологическая активность
[ редактировать ]Гормональная активность
[ редактировать ]Андрогенный рецептор
[ редактировать ]Хотя он действует как эндогенный предшественник более мощных андрогенов, таких как тестостерон и ДГТ, было обнаружено, что ДГЭА обладает некоторой степенью андрогенной сам по себе низким сродством (K i = 1 мкМ). активности, действуя как слабый частичный агонист с андрогенный рецептор (АР). Однако его внутренняя активность в отношении рецептора довольно слаба, и из-за конкуренции за связывание с полными агонистами , такими как тестостерон, он может фактически вести себя скорее как антагонист в зависимости от циркулирующих уровней тестостерона и дигидротестостерона (ДГТ), и, следовательно, , как антиандроген . Однако его сродство к рецептору очень низкое и по этой причине вряд ли будет иметь большое значение в нормальных обстоятельствах. [19] [23]
Эстрогеновые рецепторы
[ редактировать ]Помимо сродства к андрогенным рецепторам, DHEA также связывается с ERα и ERβ рецепторами эстрогена (и активирует их) со значениями K i 1,1 мкМ и 0,5 мкМ соответственно и значениями EC 50 >1 мкМ и 200 нМ соответственно. Хотя было обнаружено, что он является частичным агонистом ERα с максимальной эффективностью 30–70%, концентрации, необходимые для такой степени активации, делают маловероятным, что активность DHEA в отношении этого рецептора является физиологически значимой. Примечательно, однако, что DHEA действует как полный агонист ERβ с максимальным ответом, аналогичным или немного превышающим таковой эстрадиола , а его уровни в кровообращении и местных тканях человеческого тела достаточно высоки, чтобы активировать рецептор в той же степени. как это наблюдается при уровнях циркулирующего эстрадиола, несколько превышающих их максимальные, неовуляторные концентрации ; действительно, в сочетании с эстрадиолом, когда оба препарата находились на уровнях, эквивалентных их физиологическим концентрациям, общая активация ERβ удваивалась. [10] [19]
Другие ядерные рецепторы
[ редактировать ]DHEA не связывается и не активирует рецепторы прогестерона , глюкокортикоидов или минералокортикоидов . [19] [24] Другие ядерные рецепторы- мишени DHEA, помимо рецепторов андрогенов и эстрогенов, включают PPARα , PXR и CAR . [25] Однако, хотя DHEA является лигандом PPARα и PXR у грызунов, у человека он отсутствует. [26] Считается, что помимо прямых взаимодействий DHEA регулирует ряд других белков посредством непрямых геномных механизмов, включая ферменты CYP2C11 и 11β-HSD1 , последний из которых необходим для биосинтеза глюкокортикоидов, таких как кортизол , и, как было предложено, участвовать в антиглюкокортикоидном эффекте DHEA – и белка-носителя IGFBP1 . [19] [27]
Нейростероидная активность
[ редактировать ]Рецепторы нейромедиаторов
[ редактировать ]Было обнаружено, что DHEA напрямую действует на несколько нейромедиаторов , в том числе действует как аллостерический модулятор рецептора NMDA , как отрицательный аллостерический модулятор рецептора GABA A рецепторов и как агонист рецептора σ1 - положительный . [28] [25]
Нейротрофиновые рецепторы
[ редактировать ]В 2011 году было сделано удивительное открытие, что DHEA, а также его сульфатный эфир DHEA-S напрямую связываются и активируют TrkA и p75. НТР , рецепторы нейротрофинов, таких как фактор роста нервов (NGF) и нейротрофический фактор головного мозга (BDNF), с высоким сродством. [25] [29] Впоследствии было также обнаружено, что DHEA связывается с TrkB и TrkC с высоким сродством, хотя он активирует только TrkC, а не TrkB. [25] [30] DHEA и DHEA-S связывались с этими рецепторами со сродством в низком наномолярном диапазоне (около 5 нМ), которое, тем не менее, было примерно на два порядка ниже по сравнению с высокоэффективными полипептидными нейротрофинами, такими как NGF (0,01–0,1 нМ). [25] [29] [30] В любом случае, DHEA и DHEA-S циркулируют в концентрациях, необходимых для активации этих рецепторов, и поэтому были идентифицированы как важные эндогенные нейротрофические факторы . [25] [29] С тех пор их назвали «стероидными микронейротрофинами» из-за их низкомолекулярной и стероидной природы по сравнению с их полипептидными аналогами-нейротрофинами. [31] Последующие исследования показали, что ДГЭА и/или ДГЭА-С на самом деле могут быть филогенетически древними «предковыми» лигандами рецепторов нейротрофинов с ранних этапов эволюции нервной системы . [25] [30] Выводы о том, что ДГЭА связывается с рецепторами нейротрофинов и мощно активирует их , могут объяснить положительную связь между снижением уровня циркулирующего ДГЭА с возрастом и возрастными нейродегенеративными заболеваниями . [25] [29]
Белок 2, связанный с микротрубочками
[ редактировать ]что, как и прегненолон , его синтетическое производное 3β-метоксипрегненолон (MAP-4343) и прогестерон Было обнаружено, , DHEA связывается с белком 2, ассоциированным с микротрубочками (MAP2), особенно с подтипом MAP2C (K d = 27 мкМ). [25] Однако неясно, увеличивает ли ДГЭА связывание MAP2 с тубулином , как прегненолон. [25]
СДВГ
[ редактировать ]Некоторые исследования показали, что уровни ДГЭА слишком низкие у людей с СДВГ, а лечение метилфенидатом или бупропионом (стимулирующие препараты) нормализует уровни ДГЭА. [32]
Другая деятельность
[ редактировать ]Ингибитор G6PDH
[ редактировать ]DHEA является неконкурентным ингибитором глюкозо G6PDH Tooltip (K i = 17 мкМ; IC 50 = 18,7 мкМ) и способен снижать NADPH Tooltip и уменьшать НАДФН-зависимое производство свободных радикалов . [33] [34] Считается, что это действие может быть ответственным за большую часть противовоспалительных , антигиперпластических , химиопрофилактических , антигиперлипидемических , антидиабетических и противообестических , а также определенных иммуномодулирующих действий ДГЭА (при наличии некоторых экспериментальных данных, подтверждающих это мнение). [33] [34] [35] [36] Однако также сообщалось, что ингибирование активности G6PDH DHEA in vivo не наблюдалось и что концентрации, необходимые DHEA для ингибирования G6PDH in vitro , очень высоки, что делает возможный вклад ингибирования G6PDH в эффекты DHEA неопределенным. . [34]
Рак
[ редактировать ]Добавки DHEA рекламируются в виде добавок из-за заявленных ими свойств предотвращения рака; нет никаких научных доказательств, подтверждающих эти утверждения. [37]
Разнообразный
[ редактировать ]Было обнаружено, что DHEA конкурентно ингибирует TRPV1 . [28]
Биохимия
[ редактировать ]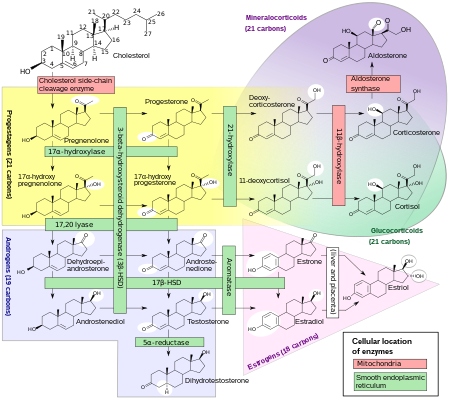
Биосинтез
[ редактировать ]ДГЭА вырабатывается в сетчатой зоне под коры надпочечников контролем адренокортикотропного гормона (АКТГ) и гонадами под контролем гонадотропин-высвобождающего гормона (ГнРГ). [39] [40] Он также вырабатывается в мозге. [41] ДГЭА синтезируется из холестерина с помощью ферментов, расщепляющих боковую цепь холестерина (CYP11A1; P450scc) и 17α-гидроксилазы/17,20-лиазы (CYP17A1), с прегненолоном и 17α-гидроксипрегненолоном в качестве промежуточных продуктов . [42] Он вырабатывается главным образом из коры надпочечников , и только около 10% секретируется половыми железами . [43] [44] [45] Примерно от 50 до 70% циркулирующего ДГЭА возникает в результате десульфатации ДГЭА-С в периферических тканях. [43] Сам ДГЭА-С происходит почти исключительно из коры надпочечников, причем у женщин от 95 до 100% секретируется корой надпочечников. [39] [45]
Увеличение эндогенного производства
[ редактировать ]Известно, что регулярные физические упражнения увеличивают выработку ДГЭА в организме. [46] [47] ограничение калорий увеличивает уровень DHEA у приматов. Также было показано, что [48] Некоторые предполагают, что увеличение эндогенного ДГЭА, вызванное ограничением калорий, частично ответственно за увеличение продолжительности жизни, которое, как известно, связано с ограничением калорий. [49]
Распределение
[ редактировать ]В кровообращении ДГЭА в основном связан с альбумином , а небольшое количество связано с глобулином, связывающим половые гормоны (ГСПГ). [50] [51] Небольшой остаток ДГЭА, не связанный с альбумином или ГСПГ, является несвязанным и находится в свободном обращении. [50]
ДГЭА легко проникает через гематоэнцефалический барьер в центральную нервную систему . [41]
Метаболизм
[ редактировать ]DHEA трансформируется в DHEA-S путем сульфатирования в положении C3β с помощью сульфотрансферазы ферментов SULT2A1 и, в меньшей степени, SULT1E1 . [42] [52] [53] Это происходит естественным путем в коре надпочечников и во время метаболизма первого прохождения в печени и кишечнике при экзогенного ДГЭА. пероральном введении [52] Уровни ДГЭА-С в кровообращении примерно в 250–300 раз выше, чем ДГЭА. [20] ДГЭА-С, в свою очередь, может превращаться обратно в ДГЭА в периферических тканях с помощью стероидсульфатазы (СТС). [54] [55]
Конечный период полувыведения ДГЭА короткий и составляет всего 15–30 минут. [56] Напротив, конечный период полувыведения ДГЭА-С намного дольше – от 7 до 10 часов. [56] Поскольку ДГЭА-С может быть преобразован обратно в ДГЭА, он служит циркулирующим резервуаром для ДГЭА, тем самым продлевая срок действия ДГЭА. [57] [20]
Метаболиты ДГЭА включают ДГЭА-С, 7α-гидрокси-ДГЭА , 7β-гидрокси-ДГЭА , 7-кето-ДГЭА , 7α-гидроксиэпиандростерон и 7β-гидроксиэпиандростерон , а также андростенедиол и андростендион . [8]
Беременность
[ редактировать ]Во время беременности ДГЭА-С метаболизируется в сульфаты 16α-гидрокси-ДГЭА и 15α-гидрокси-ДГЭА в плода печени в качестве промежуточных продуктов при выработке эстрогенов эстриола и эстетрола соответственно. [58]
Уровни
[ редактировать ]До полового созревания у человека уровни ДГЭА и ДГЭА-С повышаются при дифференцировке зоны сетчатой надпочечников коры . [25] Пиковые уровни DHEA и DHEA-S наблюдаются примерно в возрасте 20 лет, после чего следует возрастное снижение на протяжении всей жизни, в конечном итоге возвращающееся к препубертатным концентрациям. [25] Уровни ДГЭА в плазме у взрослых мужчин составляют от 10 до 25 нМ, у женщин в пременопаузе — от 5 до 30 нМ, а у женщин в постменопаузе — от 2 до 20 нМ. [25] И наоборот, уровни DHEA-S на порядок выше при концентрации 1–10 мкМ. [25] Уровни ДГЭА и ДГЭА-С снижаются до более низких наномолярных и микромолярных диапазонов у мужчин и женщин в возрасте от 60 до 80 лет. [25]
Уровни DHEA следующие: [59]
- Взрослые мужчины: 180–1250 нг/дл.
- Взрослые женщины: 130–980 нг/дл.
- Беременные женщины: 135–810 нг/дл.
- Дети препубертатного возраста (<1 года): 26–585 нг/дл.
- Дети препубертатного возраста (1–5 лет): 9–68 нг/дл.
- Дети препубертатного возраста (6–12 лет): 11–186 нг/дл.
- Мальчики-подростки (Таннер II–III): 25–300 нг/дл.
- Девочки-подростки (Таннер II–III): 69–605 нг/дл.
- Мальчики-подростки (Таннер IV–V): 100–400 нг/дл.
- Девочки-подростки (Таннер IV–V): 165–690 нг/дл.
Измерение
[ редактировать ]Поскольку почти весь ДГЭА вырабатывается надпочечниками, измерения уровня ДГЭА-С/ДГЭА в крови полезны для выявления избыточной активности надпочечников, наблюдаемой при раке или гиперплазии надпочечников, включая определенные формы врожденной гиперплазии надпочечников . Женщины с синдромом поликистозных яичников, как правило, имеют повышенный уровень ДГЭА-С. [60]
Химия
[ редактировать ]ДГЭА, также известный как андрост-5-ен-3β-ол-17-он, представляет собой природный андростана стероид и 17-кетостероид . [61] Структурно он тесно связан с андростенедиолом (андрост-5-ен-3β,17β-диол), андростендионом (андрост-4-ен-3,17-дион) и тестостероном (андрост-4-ен-17β-ол-3). -один). [61] ДГЭА является 5- ( 5α δ дегидроаналогом эпиандростерона -андростан-3β-ол-17-он) и также известен как 5-дегидроэпиандростерон или как 5 -эпиандростерон. [61]
Изомеры
[ редактировать ]Термин «дегидроэпиандростерон» неоднозначен с химической точки зрения, поскольку он не включает определенные положения в эпиандростероне, в которых отсутствуют атомы водорода. Сам ДГЭА представляет собой 5,6-дидегидроэпиандростерон или 5-дегидроэпиандростерон. Также существует ряд встречающихся в природе изомеров, которые могут иметь аналогичную активность. Некоторые изомеры ДГЭА представляют собой 1-дегидроэпиандростерон (1-андростерон) и 4-дегидроэпиандростерон . [62] Эти изомеры также технически называются «ДГЭА», поскольку они представляют собой дегидроэпиандростероны, в которых водороды удалены из скелета эпиандростерона .
Дегидроандростерон (ДГК) представляет собой 3α- эпимер ДГЭА и также является эндогенным андрогеном.
История
[ редактировать ]ДГЭА был впервые выделен из мочи человека в 1934 году Адольфом Бутенандтом и Куртом Чернингом. [63]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Девиллерс Дж. (27 апреля 2009 г.). Моделирование эндокринных нарушений . ЦРК Пресс. стр. 339–. ISBN 978-1-4200-7636-3 .
- ^ Перейти обратно: а б Капп М.Дж., Трейси Т.С. (10 декабря 2002 г.). Диетические добавки: токсикология и клиническая фармакология . Springer Science & Business Media. стр. 135–. ISBN 978-1-59259-303-3 .
- ^ Перейти обратно: а б Одденс Б.Дж., Вермюлен А. (15 ноября 1996 г.). Андрогены и стареющие мужчины . ЦРК Пресс. стр. 5–. ISBN 978-1-85070-763-9 .
- ^ Перейти обратно: а б с д и Лабри Ф., Луу-Те В., Беланжер А., Лин С.С., Симард Дж., Пеллетье Дж., Лабри С. (ноябрь 2005 г.). «Является ли дегидроэпиандростерон гормоном?» . Дж. Эндокринол . 187 (2): 169–96. дои : 10.1677/joe.1.06264 . ПМИД 16293766 .
- ^ Уильям Ф. Ганонг, доктор медицинских наук, «Обзор медицинской физиологии», 22-е изд., McGraw Hill, 2005, стр. 362.
- ^ Индекс Merck , 13-е издание, 7798.
- ^ Шульман Р.А., Дин С. (2007). Решите задачу с помощью добавок . Нью-Йорк: Rodale, Inc., с. 100. ИСБН 978-1-57954-942-8 .
ДГЭА (дегидроэпиандростерон) — это распространенный гормон, вырабатываемый надпочечниками, половыми железами и мозгом.
- ^ Перейти обратно: а б Мо Кью, Лу С.Ф., Саймон Н.Г. (апрель 2006 г.). «Дегидроэпиандростерон и его метаболиты: дифференциальное влияние на трафик андрогенных рецепторов и транскрипционную активность». Журнал биохимии стероидов и молекулярной биологии . 99 (1): 50–8. дои : 10.1016/j.jsbmb.2005.11.011 . ПМИД 16524719 . S2CID 30489004 .
- ^ Скотт Т. (1996). Краткая энциклопедия биологии . Вальтер де Грюйтер. п. 49 . ISBN 978-3-11-010661-9 . Проверено 25 мая 2012 г.
- ^ Перейти обратно: а б с Уэбб С.Дж., Геогеган Т.Э., Проф Р.А., Майкл Миллер К.К. (2006). «Биологическое действие дегидроэпиандростерона затрагивает множество рецепторов» . Обзоры метаболизма лекарств . 38 (1–2): 89–116. дои : 10.1080/03602530600569877 . ПМЦ 2423429 . ПМИД 16684650 .
- ^ Фрисс Э., Шиффельхольц Т., Стеклер Т., Штайгер А. (декабрь 2000 г.). «Дегидроэпиандростерон - нейростероид». Европейский журнал клинических исследований . 30 (Приложение 3): 46–50. дои : 10.1046/j.1365-2362.2000.0300s3046.x . ПМИД 11281367 . S2CID 30733847 .
- ^ Песковиц О.Г., Югстер Э.А. (2004). Детская эндокринология: механизмы, проявления и управление . Липпинкотт Уильямс и Уилкинс. стр. 362–. ISBN 978-0-7817-4059-3 .
- ^ Фима Лифшиц (26 декабря 2006 г.). Детская эндокринология: нарушения роста, надпочечников, половой функции, щитовидной железы, кальция и баланса жидкости . ЦРК Пресс. стр. 289–. ISBN 978-1-4200-4272-6 .
- ^ Салхан С. (1 августа 2011 г.). Учебник гинекологии . JP Medical Ltd., стр. 94–. ISBN 978-93-5025-369-4 .
- ^ Лавери Дж. П., Санфилиппо Дж. С. (6 декабря 2012 г.). Детская и подростковая акушерство и гинекология . Springer Science & Business Media. стр. 45–. ISBN 978-1-4612-5064-7 .
- ^ Нуссбаум Р.Л., Макиннес Р.Р., Уиллард Х.Ф. (28 апреля 2015 г.). Томпсон и Томпсон Генетика в медицине . Elsevier Науки о здоровье. стр. 102–. ISBN 978-0-323-39206-8 .
- ^ Сетчелл, М.Э., Хадсон, штат Коннектикут (4 апреля 2013 г.). Учебник Шоу по оперативной гинекологии . Elsevier Науки о здоровье. стр. 129–. ISBN 978-81-312-3481-5 .
- ^ Биссоннетт Б., Даленс Б. (20 июля 2006 г.). Синдромы: быстрое распознавание и периоперационные последствия . МакГроу Хилл Профессионал. п. 184. ИСБН 978-0-07-135455-4 .
- ^ Перейти обратно: а б с д и Чен Ф., Кнехт К., Бирзин Э., Фишер Дж., Уилкинсон Х., Мойена М., Морено К.Т., Шмидт А., Харада С., Фридман Л.П., Решка А.А. (ноябрь 2005 г.). «Прямые функции агониста/антагониста дегидроэпиандростерона» . Эндокринология . 146 (11): 4568–76. дои : 10.1210/en.2005-0368 . ПМИД 15994348 .
- ^ Перейти обратно: а б с Вейцман А (1 февраля 2008 г.). Нейроактивные стероиды в функции мозга, поведении и нервно-психических расстройствах: новые стратегии исследований и лечения . Springer Science & Business Media. стр. 229–. ISBN 978-1-4020-6854-6 .
- ^ Граванис АГ, Меллон С.Х. (24 июня 2011 г.). Гормоны в нейродегенерации, нейропротекции и нейрогенезе . Джон Уайли и сыновья. стр. 349–. ISBN 978-3-527-63397-5 .
- ^ Половые различия в человеческом мозге, их основы и последствия . Эльзевир. 3 декабря 2010 г. стр. 127–. ISBN 978-0-444-53631-0 .
- ^ Гао В., Бол CE, Далтон Дж. Т. (сентябрь 2005 г.). «Химия и структурная биология андрогенных рецепторов» . Химические обзоры . 105 (9): 3352–70. дои : 10.1021/cr020456u . ПМК 2096617 . ПМИД 16159155 .
- ^ Линдшау С., Кирш Т., Клинге У., Колкхоф П., Петерс И., Фибелер А. (сентябрь 2011 г.). «Индуцированное дегидроэпиандростероном фосфорилирование и транслокация FoxO1 зависят от минералокортикоидного рецептора» . Гипертония . 58 (3): 471–8. doi : 10.1161/ГИПЕРТЕНЗИЯХА.111.171280 . ПМИД 21747041 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Проф Р.А., Кларк Б.Дж., Клинге К.М. (апрель 2016 г.). «Новые механизмы действия ДГЭА» . Журнал молекулярной эндокринологии . 56 (3): Р139–55. дои : 10.1530/JME-16-0013 . ПМИД 26908835 .
- ^ Уотсон Р.Р. (22 июля 2011 г.). ДГЭА в здоровье человека и старении . ЦРК Пресс. стр. 208–. ISBN 978-1-4398-3884-6 .
- ^ Калими М., Шафагой Ю., Лория Р., Пэджетт Д., Регелсон В. (февраль 1994 г.). «Антиглюкокортикоидные эффекты дегидроэпиандростерона (ДГЭА)». Молекулярная и клеточная биохимия . 131 (2): 99–104. дои : 10.1007/BF00925945 . ПМИД 8035785 . S2CID 26893297 .
- ^ Перейти обратно: а б Король СР (9 ноября 2012 г.). Нейростероиды и нервная система . Springer Science & Business Media. стр. 15–16. ISBN 978-1-4614-5559-2 .
- ^ Перейти обратно: а б с д Лазаридис I, Харалампопулос I, Алексаки VI, Авлонитис Н, Педиадитакис I, Эфстатопулос П, Калогеропулу Т, Кастанас Э, Граванис А (апрель 2011 г.). «Нейростероид дегидроэпиандростерон взаимодействует с рецепторами фактора роста нервов (NGF), предотвращая апоптоз нейронов» . ПЛОС Биология . 9 (4): e1001051. дои : 10.1371/journal.pbio.1001051 . ПМК 3082517 . ПМИД 21541365 .
- ^ Перейти обратно: а б с Педиадитакис I, Илиопулос I, Теологидис I, Деливаноглу Н, Маргиорис А.Н., Харалампопулос I, Граванис А (январь 2015 г.). «Дегидроэпиандростерон: предковый лиганд рецепторов нейротрофинов» . Эндокринология . 156 (1): 16–23. дои : 10.1210/en.2014-1596 . ПМИД 25330101 .
- ^ Граванис А., Калогеропулу Т., Панутсакопулу В., Термос К., Неофиту С., Харалампопулос I (октябрь 2012 г.). «Нейростероиды и микронейротрофины передают сигнал через рецепторы NGF, чтобы индуцировать передачу сигналов, способствующих выживанию, в нейрональных клетках». Научная сигнализация . 5 (246): пт8. дои : 10.1126/scisignal.2003387 . ПМИД 23074265 . S2CID 26914550 .
- ^ Ли, MS; Ян, JW; Ко, Ю.Х.; Хан, К.; Ким, Ш.; Ли, MS; Джо, Ш.; Юнг, И.К. (2008). «Влияние метилфенидата и бупропиона на уровни DHEA-S и кортизола в плазме при синдроме дефицита внимания и гиперактивности» . Детская психиатрия и развитие человека . 39 (2): 201–209. дои : 10.1007/s10578-007-0081-6 . ПМИД 17763937 . S2CID 11041447 .
- ^ Перейти обратно: а б Шварц А.Г., Пашко Л.Л. (апрель 2004 г.). «Дегидроэпиандростерон, глюкозо-6-фосфатдегидрогеназа и долголетие». Обзоры исследований старения . 3 (2): 171–87. дои : 10.1016/J.arr.2003.05.001 . ПМИД 15177053 . S2CID 11871872 .
- ^ Перейти обратно: а б с Чолино HP, Макдональд CJ, Yeh GC (июль 2002 г.). «Ингибирование ферментов, активирующих канцерогены, 16альфа-фтор-5-андростен-17-оном». Исследования рака . 62 (13): 3685–90. ПМИД 12097275 .
- ^ Маккормик Д.Л., Джонсон В.Д., Козуб Н.М., Рао К.В., Любет Р.А., Стил В.Е., Босланд М.К. (февраль 2007 г.). «Химиопрофилактика канцерогенеза простаты у крыс с помощью диетического 16альфа-фтор-5-андростен-17-она (флуастерона), минимально андрогенного аналога дегидроэпиандростерона» . Канцерогенез . 28 (2): 398–403. дои : 10.1093/carcin/bgl141 . ПМИД 16952912 .
- ^ Аучи Д., Калер Л., Субраманиан С., Хуанг Й., Фринк Дж., Ридинг С., Оффнер Х. (сентябрь 2007 г.). «Новый биодоступный перорально синтетический андростен ингибирует коллаген-индуцированный артрит у мышей: гормоны андростена как регуляторы регуляторных Т-клеток». Анналы Нью-Йоркской академии наук . 1110 (1): 630–40. Бибкод : 2007NYASA1110..630A . дои : 10.1196/анналы.1423.066 . ПМИД 17911478 . S2CID 32258529 .
- ^ Рассел Дж., Ровер А., ред. (2009). «ДГЭА» . Полное руководство Американского онкологического общества по дополнительным и альтернативным методам лечения рака (2-е изд.). Американское онкологическое общество. стр. 729–733 . ISBN 9780944235713 .
- ^ Хэггстрем, Микаэль; Ричфилд, Дэвид (2014). «Схема путей стероидогенеза человека» . Викижурнал медицины . 1 (1). дои : 10.15347/wjm/2014.005 . ISSN 2002-4436 .
- ^ Перейти обратно: а б Эрккола Р. (2006). Менопауза . Эльзевир. стр. 5–. ISBN 978-0-444-51830-9 .
- ^ Кляйне Б., Россманит WG (11 февраля 2016 г.). Гормоны и эндокринная система: Учебник эндокринологии . Спрингер. стр. 264–265. ISBN 978-3-319-15060-4 .
- ^ Перейти обратно: а б Пиццорно Дж. Э. (2013). Учебник натуральной медицины . Elsevier Науки о здоровье. стр. 711–. ISBN 978-1-4377-2333-5 .
- ^ Перейти обратно: а б Рейни В.Е., Накамура Ю. (февраль 2008 г.). «Регуляция биосинтеза андрогенов надпочечниками» . Журнал биохимии стероидов и молекулярной биологии . 108 (3–5): 281–6. дои : 10.1016/j.jsbmb.2007.09.015 . ПМК 2699571 . ПМИД 17945481 .
- ^ Перейти обратно: а б Адлер РА (14 декабря 2009 г.). Остеопороз: патофизиология и клиническое лечение . Springer Science & Business Media. стр. 387–. ISBN 978-1-934115-19-0 .
- ^ Шилл В.Б., Комхэр Ф.Х., Харгрив Т.Б. (26 августа 2006 г.). Андрология для клинициста . Springer Science & Business Media. стр. 243–. ISBN 978-3-540-33713-3 .
- ^ Перейти обратно: а б Линос Д.А., ван Херден Дж.А. (5 декабря 2005 г.). Надпочечники: аспекты диагностики и хирургического лечения . Springer Science & Business Media. стр. 161–. ISBN 978-3-540-26861-1 .
- ^ Филер Э., Дюше П., Лак Дж. (октябрь 1998 г.). «Влияние количества тренировок на концентрацию кортизола, дегидроэпиандростерона в слюне и на соотношение концентраций дегидроэпиандростерона: кортизола у женщин за 16 недель тренировок». Европейский журнал прикладной физиологии и физиологии труда . 78 (5): 466–71. дои : 10.1007/s004210050447 . ПМИД 9809849 . S2CID 20583279 .
- ^ Коупленд Дж.Л., Конситт Л.А., Трембле М.С. (апрель 2002 г.). «Гормональные реакции на упражнения на выносливость и сопротивление у женщин в возрасте 19-69 лет» . Журналы геронтологии. Серия А, Биологические и медицинские науки . 57 (4): Б158–65. дои : 10.1093/gerona/57.4.B158 . ПМИД 11909881 .
- ^ Мэттисон Дж.А., Лейн М.А., Рот Г.С., Ингрэм Д.К. (2003). «Ограничение калорий у макак-резус» . Экспериментальная геронтология . 38 (1–2): 35–46. дои : 10.1016/S0531-5565(02)00146-8 . ПМИД 12543259 . S2CID 41481691 . .
- ^ Робертс Э. (февраль 1999 г.). «Важность наличия дегидроэпиандростеронсульфата (в крови приматов): более долгая и здоровая жизнь?». Биохимическая фармакология . 57 (4): 329–46. дои : 10.1016/S0006-2952(98)00246-9 . ПМИД 9933021 . .
- ^ Перейти обратно: а б Алески С., Маноли И., Блэкман М.Р. (29 декабря 2004 г.). «Дегидроэпиандростерон (ДГЭА)» . В Коутс П.М., Блэкман М.Р., Крэгг Г.М., Левин М., Мосс Дж., Уайт Дж.Д. (ред.). Энциклопедия пищевых добавок (Печать) . ЦРК Пресс. стр. 169–. ISBN 978-0-8247-5504-1 .
- ^ Беккер К.Л. (2001). Принципы и практика эндокринологии и обмена веществ . Липпинкотт Уильямс и Уилкинс. стр. 712–. ISBN 978-0-7817-1750-2 .
- ^ Перейти обратно: а б Мюллер Дж.В., Гиллиган Л.К., Идковяк Дж., Арлт В., Фостер П.А. (октябрь 2015 г.). «Регулирование действия стероидов путем сульфатации и десульфатации» . Эндокр преп . 36 (5): 526–63. дои : 10.1210/er.2015-1036 . ПМЦ 4591525 . ПМИД 26213785 .
- ^ Лэш Л.Х. (2005). Метаболизм и транспорт лекарств: молекулярные методы и механизмы . Springer Science & Business Media. стр. 353–. ISBN 978-1-59259-832-8 .
- ^ Морфин Р. (2 сентября 2003 г.). ДГЭА и мозг . ЦРК Пресс. стр. 28–. ISBN 978-0-203-30121-0 .
- ^ Карасек М. (2006). Старение и возрастные заболевания: основы . Издательство Нова. стр. 66–. ISBN 978-1-59454-426-2 .
- ^ Перейти обратно: а б Белый BA, Портерфилд SP (2013). Эндокринная и репродуктивная физиология, серия монографий Мосби по физиологии (с онлайн-доступом для студентов), 4: Эндокринная и репродуктивная физиология . Elsevier Науки о здоровье. стр. 164–. ISBN 978-0-323-08704-9 .
- ^ Калими М.Ю., Регельсон В. (2000). Дегидроэпиандростерон (ДГЭА): биохимические, физиологические и клинические аспекты . Вальтер де Грюйтер. стр. 41–. ISBN 978-3-11-016111-3 .
- ^ Збелла, Э.А.; Илекис, Дж.; Скомменья, А.; Бенвенист, Р. (1986). «Конкурентные исследования сульфата дегидроэпиандростерона и сульфата 16 альфа-гидроксидегидроэпиандростерона в культивируемых клетках хориокарциномы человека JEG-3: влияние на секрецию эстрона, 17 бета-эстрадиола и эстриола» . Журнал клинической эндокринологии и метаболизма . 63 (3): 751–757. doi : 10.1210/jcem-63-3-751 . ISSN 0021-972X . ПМИД 2942557 .
- ^ «ДГЭА (дегидроэпиандростерон)» (PDF) . Квест Диагностика . Архивировано из оригинала (PDF) 27 сентября 2020 г.
- ^ Банашевска Б, Спачиньски Р.З., Пелеш М., Павельчик Л. (2003). «Частота повышенного соотношения ЛГ/ФСГ у женщин с синдромом поликистозных яичников с нормо- и гиперинсулинемией». Летопись Медицинской академии в Белостоке . 48 : 131–4. ПМИД 14737959 .
- ^ Перейти обратно: а б с Элкс Дж. (14 ноября 2014 г.). Словарь лекарств: Химические данные: Химические данные, структуры и библиография . Спрингер. стр. 641–. ISBN 978-1-4757-2085-3 .
- ^ Эдит Джозефи; Ф. Радт (1 декабря 2013 г.). Энциклопедия органической химии Elsevier: Серия III: Карбоизоциклические конденсированные соединения . Спрингер. стр. 2608–. ISBN 978-3-662-25863-7 .
- ^ Шварц А.Г., Пашко Л.Л. (2001). «Потенциальное терапевтическое использование дегидроэпиандростерона и структурных аналогов». Технология и терапия диабета . 3 (2): 221–4. дои : 10.1089/152091501300209589 . ПМИД 11478328 .
Дальнейшее чтение
[ редактировать ]- Лабри Ф., Мартель С., Беланжер А., Пеллетье Дж. (апрель 2017 г.). «По данным интракринологии, андрогены у женщин по существу состоят из ДГЭА в каждой периферической ткани». Журнал биохимии стероидов и молекулярной биологии . 168 : 9–18. дои : 10.1016/j.jsbmb.2016.12.007 . ПМИД 28153489 . S2CID 2620899 .