Конкурентное ингибирование

22Ч
5 OH ) служит конкурентным ингибитором метанола ( CH
3 OH ) и этиленгликоль ( (CH 2 OH) 2 ) для фермента алкогольдегидрогеназы в печени , когда он присутствует в больших количествах. По этой причине этанол иногда используется в качестве средства для лечения или предотвращения токсичности после случайного проглатывания этих химикатов.
Конкурентное ингибирование — это прерывание химического пути из-за того, что одно химическое вещество подавляет действие другого, конкурируя с ним за связывание или связывание . любую метаболическую или химическую информационную Этот принцип потенциально может повлиять на систему, но в биохимии и медицине особенно важны несколько классов конкурентного ингибирования , включая конкурентную форму ингибирования ферментов , конкурентную форму антагонизма рецепторов , конкурентную форму антиметаболитной активности, и конкурентная форма отравления (которая может включать любой из перечисленных видов).
Тип ингибирования ферментов
[ редактировать ]При конкурентном ингибировании ферментативного катализа связывание ингибитора предотвращает связывание целевой молекулы фермента, также известной как субстрат. [1] Это достигается путем блокирования каким-либо образом сайта связывания субстрата – активного сайта. V max указывает максимальную скорость реакции, а K m представляет собой количество субстрата, необходимое для достижения половины V max . K m также играет роль в определении склонности субстрата связывать фермент. [2] Конкурентное ингибирование можно преодолеть, добавляя в реакцию больше субстрата, что увеличивает вероятность связывания фермента и субстрата. В результате конкурентное ингибирование изменяет только K m , оставляя V max прежним. [3] Это можно продемонстрировать с помощью графиков кинетики ферментов, таких как график Михаэлиса-Ментен или график Лайнуивера-Бёрка . Как только ингибитор связывается с ферментом, наклон будет изменен, поскольку K m либо увеличивается, либо уменьшается по сравнению с исходным K m реакции. [4] [5] [6]
Большинство конкурентных ингибиторов действуют путем обратимого связывания с активным центром фермента. [1] В результате многие источники утверждают, что это определяющая особенность конкурентных ингибиторов. [7] Однако это ошибочное упрощение , поскольку существует множество возможных механизмов, с помощью которых фермент может связывать либо ингибитор, либо субстрат, но никогда то и другое одновременно. [1] Например, аллостерические ингибиторы могут проявлять конкурентное, неконкурентное или неконкурентное ингибирование. [1]
Механизм
[ редактировать ]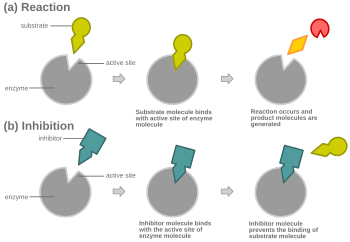
При конкурентном ингибировании ингибитор, напоминающий нормальный субстрат, связывается с ферментом, обычно в активном центре , и предотвращает связывание субстрата. [8] В любой момент фермент может быть связан с ингибитором, субстратом или ни с тем, ни с другим, но он не может связываться с ними обоими одновременно. Во время конкурентного ингибирования ингибитор и субстрат конкурируют за активный центр. Активный центр — это область фермента, с которой может связываться определенный белок или субстрат. Таким образом, активный сайт позволит только одному из двух комплексов связываться с сайтом, либо позволяя протекать реакции, либо вызывая ее. При конкурентном ингибировании ингибитор напоминает субстрат, занимая его место и связываясь с активным центром фермента. Увеличение концентрации субстрата уменьшит «конкуренцию» за правильное связывание субстрата с активным центром и позволит протекать реакции. [3] Когда концентрация субстрата превышает концентрацию конкурентного ингибитора, более вероятно, что субстрат вступит в контакт с активным центром фермента, чем с ингибитором.
Конкурентные ингибиторы обычно используются для производства фармацевтических препаратов. [3] Например, метотрексат — химиотерапевтический препарат, действующий как конкурентный ингибитор. Он структурно подобен коферменту фолату , который связывается с ферментом дигидрофолатредуктазой . [3] Этот фермент участвует в синтезе ДНК и РНК, и когда метотрексат связывает фермент, он делает его неактивным, так что он не может синтезировать ДНК и РНК. [3] Таким образом, раковые клетки не могут расти и делиться. Другой пример: простагландин вырабатывается в больших количествах в ответ на боль и может вызвать воспаление. Незаменимые жирные кислоты образуют простагландины; когда это было обнаружено, оказалось, что на самом деле это очень хорошие ингибиторы простагландинов. Эти ингибиторы жирных кислот использовались в качестве лекарств для облегчения боли, поскольку они могут действовать как субстрат, связываться с ферментом и блокировать простагландины. [9]
Примером конкурентного подавления, не связанного с лекарствами, является предотвращение потемнения фруктов и овощей. Например, тирозиназа , фермент грибов, обычно связывается с субстратом, монофенолами , и образует коричневые о-хиноны. [10] Конкурентные субстраты, такие как 4-замещенные бензальдегиды грибов, конкурируют с субстратом, снижая количество связывающихся монофенолов. Эти ингибирующие соединения, добавленные в продукт, сохраняют его свежесть в течение более длительного периода времени, уменьшая связывание монофенолов, вызывающих потемнение. [10] Это позволяет повысить качество продукции и увеличить срок ее хранения.
Конкурентное ингибирование может быть обратимым и необратимым. Если это обратимое ингибирование , то действие ингибитора можно преодолеть за счет увеличения концентрации субстрата. [8] Если оно необратимо, единственный способ преодолеть его — это производить больше мишени (и, как правило, разрушать и/или выводить из организма необратимо ингибированную мишень).
Практически в каждом случае конкурентные ингибиторы связываются в том же сайте связывания (активном сайте), что и субстрат, но связывание в том же сайте не является обязательным. Конкурентный ингибитор может связываться с аллостерическим сайтом свободного фермента и предотвращать связывание субстрата, если он не связывается с аллостерическим сайтом при связывании субстрата. Например, стрихнин действует как аллостерический ингибитор глициновых рецепторов в спинном мозге и стволе головного мозга млекопитающих. Глицин является основным постсинаптическим тормозным нейромедиатором со специфическим рецепторным участком. Стрихнин связывается с альтернативным сайтом, который снижает сродство глицинового рецептора к глицину, что приводит к судорогам из-за уменьшения ингибирования глицином. [11]
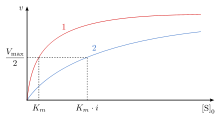
При конкурентном торможении максимальная скорость ( ) реакции не изменяется, тогда как кажущееся сродство субстрата к месту связывания снижается ( константа диссоциации, по-видимому, увеличивается). Изменение ( константа Михаэлиса-Ментен ) параллельна изменению , поскольку одно увеличивается, другое должно уменьшаться. Когда конкурентный ингибитор связывается с ферментом, увеличивается. Это означает, что аффинность связывания фермента снижается, но ее можно преодолеть, увеличив концентрацию субстрата. [12] Любую данную концентрацию конкурентного ингибитора можно преодолеть за счет увеличения концентрации субстрата. В этом случае субстрат уменьшит доступность ингибитора для связывания и, таким образом, вытеснит ингибитор в связывании с ферментом. [12]




Биологические примеры
[ редактировать ]После случайного приема загрязненного опиоидного препарата десметилпродина нейротоксический МФТП эффект 1-метил-4-фенил-1,2,3,6-тетрагидропиридина ( ) был обнаружен . МФТП способен преодолевать гематоэнцефалический барьер и проникать в кислые лизосомы . [13] МФТП биологически активируется МАО-В, изоферментом моноаминоксидазы (МАО), который в основном концентрируется при неврологических расстройствах и заболеваниях. [14] Позже было обнаружено, что МФТП вызывает симптомы, схожие с симптомами болезни Паркинсона . Клетки центральной нервной системы (астроциты) содержат МАО-В, который окисляет МФТП до 1-метил-4-фенилпиридиния (МФП+), который является токсичным. [13] MPP+ в конечном итоге попадает во внеклеточную жидкость с помощью переносчика дофамина , что в конечном итоге вызывает симптомы болезни Паркинсона. Однако конкурентное ингибирование фермента МАО-В или переносчика дофамина защищает от окисления MPTP до MPP+. Несколько соединений были протестированы на способность ингибировать окисление MPTP до MPP+, включая метиленовый синий , 5-нитроиндазол , норхарман , 9-метилнорхарман и менадион . [14] Они продемонстрировали снижение нейротоксичности, вызываемой MPTP.

Сульфамидные препараты также действуют как конкурентные ингибиторы. Например, сульфаниламид конкурентно связывается с ферментом в активном центре дигидроптероатсинтазы (DHPS), имитируя субстрат парааминобензойную кислоту (ПАБА). [15] Это предотвращает связывание самого субстрата, что останавливает выработку фолиевой кислоты, важного питательного вещества. Бактерии должны синтезировать фолиевую кислоту, поскольку у них нет ее переносчика. Без фолиевой кислоты бактерии не могут расти и делиться. Таким образом, благодаря конкурентному ингибированию сульфаниламидных препаратов они являются отличными антибактериальными средствами.Пример конкурентного ингибирования был продемонстрирован экспериментально для фермента сукцинатдегидрогеназы, катализирующего окисление сукцината до фумарата в цикле Кребса . Малонат является конкурентным ингибитором сукцинатдегидрогеназы. Связывание сукцинатдегидрогеназы с субстратом сукцинатом конкурентно ингибируется. Это происходит потому, что по химическому составу малонат похож на сукцинат. Способность малоната ингибировать связывание фермента и субстрата основана на соотношении малоната и сукцината. Малонат связывается с активным центром сукцинатдегидрогеназы, а сукцинат не может. Таким образом, он подавляет реакцию. [16]
Уравнение
[ редактировать ]Модель Михаэлиса-Ментен может стать бесценным инструментом для понимания кинетики ферментов. Согласно этой модели, график скорости реакции (V 0 ), связанной с концентрацией [S] субстрата, может затем использоваться для определения таких значений, как V max , начальная скорость и K m (V max /2 или сродство фермент-субстратный комплекс). [4]
Конкурентное ингибирование увеличивает кажущееся значение константы Михаэлиса-Ментен , , такой, что начальная скорость реакции, , определяется


![{\displaystyle V_{0}={\frac {V_{\max }\,[S]}{K_{m}^{\text{app}}+[S]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/7e6ea985b759c13c54c08b2e1a1f7b292c549beb)
где , - константа диссоциации ингибитора и – концентрация ингибитора.
![{\displaystyle K_{m}^{\text{app}}=K_{m}(1+[I]/K_{i})}](https://wikimedia.org/api/rest_v1/media/math/render/svg/e013bb3d416acd07c0c55ee196d46e9ede190a63)

![{\displaystyle [I]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/5962300a54e8ce8b5761dac9a5fbbca450c2ce0f)
остается прежним, поскольку присутствие ингибитора можно преодолеть за счет более высоких концентраций субстрата. , концентрация субстрата, необходимая для достижения , увеличивается при наличии конкурентного ингибитора. Это связано с тем, что концентрация субстрата, необходимая для достижения с ингибитором превышает концентрацию субстрата, необходимую для достижения без ингибитора.

Вывод
[ редактировать ]В простейшем случае односубстратного фермента, подчиняющегося кинетике Михаэлиса–Ментен, типичная схема
![{\displaystyle {\what {E + S <=>[k_1][k_{-1}] ES ->[k_2] E + P}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/aa34fa0b8ef16e2c4886b79b41fd6a84e83a68fd)
модифицируется для включения связывания ингибитора со свободным ферментом:
![{\displaystyle {\ce {EI + S <=>[k_{-3}][k_3] E + S + I <=>[k_1][k_{-1}] ES + I ->[k_2] E + П + Я}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/9c358fddd797429f33b9c778a4bf4f669e8000b0)
Обратите внимание, что ингибитор не связывается с комплексом ES, а субстрат не связывается с комплексом EI. Обычно предполагается, что такое поведение указывает на связывание обоих соединений в одном и том же сайте, но это не является строго необходимым. Как и при выводе уравнения Михаэлиса-Ментен, предположим, что система находится в стационарном состоянии, т. е. концентрация каждого вида ферментов не меняется.
![{\displaystyle {\frac {d[{\ce {E}}]}{dt}}={\frac {d[{\ce {ES}}]}{dt}}={\frac {d[{ \ce {EI}}]}{dt}}=0.}](https://wikimedia.org/api/rest_v1/media/math/render/svg/7d5d8b4f228bd04aff5488c2efbc9f11f6a27a4b)
Кроме того, известная общая концентрация фермента равна , а скорость измеряется в условиях, когда концентрации субстрата и ингибитора существенно не изменяются и накопилось незначительное количество продукта.
![{\displaystyle [{\ce {E}}]_{0}=[{\ce {E}}]+[{\ce {ES}}]+[{\ce {EI}}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/ee7863f535867e7f0b9e3619c9d5431b8d14851a)
Таким образом, мы можем составить систему уравнений:
| ( 1 ) |
| ( 2 ) |
![{\displaystyle {\frac {d[{\ce {E}}]}{dt}}=0=-k_ {1}[{\ce {E}}][{\ce {S}}]+k_ {-1}[{\what {ES}}]+k_{2}[{\what {ES}}]-k_{3}[{\what {E}}][{\what {I}}] +k_{-3}[{\что {EI}}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/524f829178a7d543a7f1dfe0d3836d3f2cd9beca)
| ( 3 ) |
![{\displaystyle {\frac {d[{\ce {ES}}]}{dt}}=0=k_{1}[{\ce {E}}][{\ce {S}}]-k_{ -1}[{\ce {ES}}]-k_{2}[{\ce {ES}}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/deee1b4eb1dae3e5957260a6e625632daa130b8c)
| ( 4 ) |
![{\displaystyle {\frac {d[{\ce {EI}}]}{dt}}=0=k_{3}[{\ce {E}}][{\ce {I}}]-k_{ -3}[ЭИ]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/632d3667fd0a9b85d3e0572d171e0ee8b67e8668)
где и известны. Начальная скорость определяется как , поэтому нам нужно определить неизвестное с точки зрения известного и .
![{\displaystyle {\что {[S], [I]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/1ec1dbaad807d3af650d3805d798ed15120624bd)
![{\displaystyle {\что {[E]_0}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/9a654ad516b5d2547fd9980bb7590464441d6c38)
![{\displaystyle V_{0}=d[{\ce {P}}]/dt=k_{2}[{\ce {ES}}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/0cee1a793ade4b98a68c506615766985cb243899)
![{\displaystyle {\ce {[ES]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/a96c38a14c944f4ae51601cd8670fd2bf8e12331)
Из уравнения ( 3 ) мы можем определить E через ES , переставив его в
![{\displaystyle k_{1}[{\ce {E}}][{\ce {S}}]=(k_{-1}+k_{2})[{\ce {ES}}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/940c092e01999c100f4d5fff6650b39dff53de02)
Деление на дает
![{\displaystyle k_{1}[{\ce {S}}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/28690106a4f72a9371157eefe3e178d952cb36f0)
![{\displaystyle [{\ce {E}}]={\frac {(k_{-1}+k_{2})[{\ce {ES}}]}{k_{1}[{\ce {S }}]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/41cd11072115b7e97377d2dc62bd80a0e9460cd5)
Как и при выводе уравнения Михаэлиса–Ментен, член можно заменить макроскопической константой скорости :

| ( 5 ) |
![{\displaystyle [{\ce {E}}]={\frac {K_{m}[{\ce {ES}}]}{\ce {[S]}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/5caf94a9df39bcbeae36e01bfcd1f48e27626ce9)
Подставив уравнение ( 5 ) в уравнение ( 4 ), получим
![{\displaystyle 0={\frac {k_{3}[{\ce {I}}]K_{m}[{\ce {ES}}]}{\ce {[S]}}}-k_{- 3}[{\что {EI}}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/3612eb7c921915246bb9dd47363904140d563374)
Переставляя, мы находим, что
![{\displaystyle [{\ce {EI}}]={\frac {K_{m}k_{3}[{\ce {I}}][{\ce {ES}}]}{k_{-3} [{\что {S}}]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/b5e8681d305f3fa55e808a5c06fad3748f06e131)
На этом этапе мы можем определить константу диссоциации ингибитора как , давая

| ( 6 ) |
![{\displaystyle [{\ce {EI}}]={\frac {K_{m}[{\ce {I}}][{\ce {ES}}]}{K_{i}[{\ce { С}}]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/7b5b6d93be687b516fc668d4934ac27e5972ae30)
На этом этапе подставьте уравнение ( 5 ) и уравнение ( 6 ) в уравнение ( 1 ):
![{\displaystyle [{\ce {E}}]_{0}={\frac {K_{m}[{\ce {ES}}]}{\ce {[S]}}}+[{\ce {ES}}]+{\frac {K_{m}[{\ce {I}}][{\ce {ES}}]}{K_{i}[{\ce {S}}]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/68c1417ec7ba5d7255a28927ed40932af9323e01)
Переставляя решение для ES, находим
![{\displaystyle [{\ce {E}}]_{0}=[{\ce {ES}}]\left({\frac {K_{m}}{\ce {[S]}}}+1 +{\frac {K_{m}[{\ce {I}}]}{K_{i}[{\ce {S}}]}}\right)=[{\ce {ES}}]{\ дробь {K_{m}K_{i}+K_{i}[{\ce {S}}]+K_{m}[{\ce {I}}]}{K_{i}[{\ce {S }}]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/2691e0b26e6bee9fe37253403c892909a07add45)
| ( 7 ) |
![{\displaystyle [{\ce {ES}}]={\frac {K_{i}[{\ce {S}}][{\ce {E}}]_{0}}{K_{m}K_ {i}+K_{i}[{\ce {S}}]+K_{m}[{\ce {I}}]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/b57aff644fb93ec40ef3663d1463f25ea158869e)
Возвращаясь к нашему выражению для , теперь у нас есть:
![{\displaystyle V_{0}=k_{2}[{\ce {ES}}]={\frac {k_{2}K_{i}[{\ce {S}}][{\ce {E} }]_{0}}{K_{m}K_{i}+K_{i}[{\ce {S}}]+K_{m}[{\ce {I}}]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/a7548c901ebd2e13620f6c9c6c30510ad5365859)
![{\displaystyle V_{0}={\frac {k_{2}[{\ce {E}}]_{0}[{\ce {S}}]}{K_{m}+[{\ce { S}}]+K_{m}{\frac {[{\ce {I}}]}{K_{i}}}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/83cb5b38fc5e1ea8d7df09f1f8fec2d8f47db635)
Поскольку скорость максимальна, когда весь фермент связан в виде фермент-субстратного комплекса, .Замена и объединение терминов в конечном итоге дает традиционную форму:
![{\displaystyle V_{\max }=k_{2}[{\ce {E}}]_{0}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/4ea107b2c44540b990daab7f21c8a35fc76f6476)
| ( 8 ) |
![{\displaystyle V_{0}={\frac {V_{\max }[{\ce {S}}]}{K_{m}\left(1+{\frac {[{\ce {I}}] }{K_{i}}}\right)+[{\ce {S}}]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/09a841f4777fa07f79bcec43de7264e95ef8f57c)
Для расчета концентрации конкурентного ингибитора что дает дробь скорости где :
![{\displaystyle {\что {[I]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/da716928de5f662f6ff1a1fc9ec0bde65f3fe2c2)


| ( 9 ) |
![{\displaystyle [{\ce {I}}]=\left({\frac {1}{f_{V{_{0}}}}}-1\right)K_{i}\left(1+{ \frac {[{\ce {S}}]}{K_{m}}}\right)}](https://wikimedia.org/api/rest_v1/media/math/render/svg/ccc99438e12f1dfdd4bdbdd0b21637c4003c470f)
Примечания и ссылки
[ редактировать ]- ^ Jump up to: а б с д «Виды торможения» . Центр трансляционной терапии НИЗ. Архивировано из оригинала 8 сентября 2011 года . Проверено 2 апреля 2012 г.
- ^ Лодиш Х., Берк А., Зипурски С.Л., Мацудайра П., Балтимор Д., Дарнелл Дж. (2000). «Функциональный дизайн белков» . Молекулярно-клеточная биология (4-е изд.).
- ^ Jump up to: а б с д и Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). «Ферменты могут быть ингибированы определенными молекулами» . Биохимия (5-е изд.).
- ^ Jump up to: а б Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). «Модель Михаэлиса-Ментен объясняет кинетические свойства многих ферментов» . Биохимия (5-е изд.).
- ^ Иди С.Г. (1942). «Ингибирование холинэстеразы физостигмином и простигмином» . Журнал биологической химии . 146 : 85–93. дои : 10.1016/S0021-9258(18)72452-6 .
- ^ Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). «Приложение: Vmax и KM можно определить с помощью графиков двойной взаимности» . Биохимия (5-е изд.).
- ^ Офардт К. «Виртуальная химическая книга» . Элмхерстский колледж. Архивировано из оригинала 17 октября 2015 года . Проверено 1 сентября 2015 г.
- ^ Jump up to: а б «Карта: биохимия бесплатно и легко (Ахерн и Раджагопал)» . Свободные тексты по биологии . 24 декабря 2014 года . Проверено 2 ноября 2017 г.
- ^ Флауэр Р.Дж. (март 1974 г.). «Препараты, ингибирующие биосинтез простагландинов» . Фармакологические обзоры . 26 (1): 33–67. ПМИД 4208101 .
- ^ Jump up to: а б Хименес М., Часарра С., Эскрибано Х., Кабанес Х., Гарсия-Кармона Ф. (август 2001 г.). «Конкурентное ингибирование грибной тирозиназы 4-замещенными бензальдегидами». Журнал сельскохозяйственной и пищевой химии . 49 (8): 4060–4063. дои : 10.1021/jf010194h . ПМИД 11513710 .
- ^ Дик РМ (2011). «Глава 2. Фармакодинамика: изучение действия лекарств». В Уэллетте Р., Джойс Дж. А. (ред.). Фармакология для медсестры-анестезиолога . Джонс и Бартлетт Обучение. ISBN 978-0-7637-8607-6 .
- ^ Jump up to: а б Voet D, Voet JG, Пратт CW (29 февраля 2016 г.). Основы биохимии: жизнь на молекулярном уровне (Пятое изд.). Хобокен, Нью-Джерси. ISBN 9781118918401 . OCLC 910538334 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Jump up to: а б Сиан Дж., Юдим М.Б., Ридерер П., Герлах М. (1999). «Синдром Паркинсона, индуцированный MPTP» . Базовая нейрохимия: молекулярные, клеточные и медицинские аспекты. 6-е издание .
- ^ Jump up to: а б Эрраис Т., Гильен Х. (август 2011 г.). «Ингибирование биоактивации нейротоксина МФТП антиоксидантами, окислительно-восстановительными агентами и ингибиторами моноаминоксидазы». Пищевая и химическая токсикология . 49 (8): 1773–1781. дои : 10.1016/j.fct.2011.04.026 . hdl : 10261/63126 . ПМИД 21554916 .
- ^ «Как действуют сульфамидные препараты» . Национальные институты здравоохранения (NIH) . 15 мая 2015 года . Проверено 2 ноября 2017 г.
- ^ Поттер В.Р., Дюбуа К.П. (март 1943 г.). «Исследование механизма транспорта водорода в тканях животных» . Журнал общей физиологии . 26 (4): 391–404. дои : 10.1085/jgp.26.4.391 . ПМК 2142566 . ПМИД 19873352 .
См. также
[ редактировать ]- Регрессия Шильда для ингибирования лигандных рецепторов
- Неконкурентное ингибирование