Прогестерон
 | |
 | |
| Имена | |
|---|---|
| Название ИЮПАК | |
| Систематическое название ИЮПАК ( 1S , 3aS , 3bS , 9aR , 9bS ,11aS ) -1-ацетил-9а,11а-диметил-1,2,3,3а,3b,4,5,8,9,9а, 9b,10,11,11a-тетрадекагидро-7H - циклопента[ а ]фенантрен-7-он | |
| Другие имена П4; [1] Прегненедион | |
| Идентификаторы | |
3D model ( JSmol ) | |
| ЧЭБИ | |
| ЧЕМБЛ | |
| ХимическийПаук | |
| Лекарственный Банк | |
| Информационная карта ECHA | 100.000.318 |
| КЕГГ | |
ПабХим CID | |
| НЕКОТОРЫЙ | |
Панель управления CompTox ( EPA ) | |
| Характеристики | |
| С 21 Н 30 О 2 | |
| Молярная масса | 314.469 g/mol |
| Температура плавления | 126 |
| войти P | 4.04 [4] |
| Фармакология | |
| G03DA04 ( ВОЗ ) | |
| Внутрь , местно / чрескожно , вагинально , внутримышечная инъекция , подкожная инъекция , подкожный имплантат. | |
| Фармакокинетика : | |
| ОМП : <10% [5] [6] | |
| • Альбумин : 80% • CBG : 18% • ГСПГ : <1% • Бесплатно: 1–2 % [7] [8] | |
| Печень ( CYP2C19 , CYP3A4 , CYP2C9 , 5α-редуктаза , 3α-HSD , 17α-гидроксилаза , 21-гидроксилаза , 20α-HSD ) [9] [10] | |
| ОМП : 16–18 часов. [5] [6] [11] IM : 22–26 часов [6] [12] СК : 13–18 часов [12] | |
| Реналь | |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |
Прогестерон ( P4 ) — это эндогенный стероид и прогестагена, половой гормон участвующий в менструальном цикле , беременности и эмбриогенезе человека и других биологических видов. [1] [13] Он принадлежит к группе стероидных гормонов, называемых прогестагенами. [13] и является основным прогестагеном в организме. Прогестерон выполняет множество важных функций в организме. Он также является важнейшим метаболическим промежуточным продуктом в производстве других эндогенных стероидов , включая половые гормоны и кортикостероиды , и играет важную роль в функционировании мозга как нейростероид . [14]
Помимо своей роли естественного гормона, прогестерон также используется в качестве лекарства, например, в сочетании с эстрогеном для контрацепции , для снижения риска матки или рака шейки матки , в заместительной гормональной терапии и в феминизирующей гормональной терапии . [15] Впервые он был предписан в 1934 году. [16]
Биологическая активность [ править ]
Прогестерон – самый важный прогестаген в организме. Будучи мощным агонистом ядерного рецептора прогестерона (nPR) (со сродством K D = 1 нМ), возникающее в результате воздействие на рибосомальную транскрипцию играет важную роль в регуляции женского воспроизводства. [13] [17] Кроме того, прогестерон является агонистом недавно открытых мембранных рецепторов прогестерона (mPR). [18] из которых экспрессия оказывает регулирующее воздействие на репродуктивную функцию ( зреление ооцитов , роды и подвижность сперматозоидов ) и рак, хотя для дальнейшего определения роли необходимы дополнительные исследования. [19] Он также действует как лиганд PGRMC1 ( компонент 1 мембраны рецептора прогестерона), который влияет на прогрессирование опухоли , метаболическую регуляцию и контроль жизнеспособности нервных клеток . [20] [21] [22] Кроме того, известно, что прогестерон является антагонистом сигма σ1 - рецептора . [23] [24] отрицательный аллостерический модулятор никотиновых ацетилхолиновых рецепторов , [14] и мощный антагонист минералокортикоидных рецепторов (MR). [25] Прогестерон предотвращает активацию MR, связываясь с этим рецептором со сродством, превышающим даже сродство альдостерона и глюкокортикоидов, таких как кортизол и кортикостерон . [25] и вызывает антиминералокортикоидные эффекты, такие как натрийурез , в физиологических концентрациях. [26] Кроме того, прогестерон связывается и действует как частичный агонист глюкокортикоидного рецептора (ГР), хотя и с очень низкой эффективностью ( EC 50 > в 100 раз меньше по сравнению с кортизолом ). [27] [28]
Прогестерон через свои нейростероидные активные метаболиты такие как 5α-дигидропрогестерон и аллопрегнанолон , действует косвенно как положительный аллостерический рецептора ГАМК А. , модулятор [29]
Прогестерон и некоторые его метаболиты, такие как 5β-дигидропрогестерон , являются агонистами прегнанового X-рецептора (PXR). [30] хотя и слабо ( EC 50 >10 мкМ). [31] Соответственно, прогестерон индуцирует несколько печеночного цитохрома P450 ферментов . [32] такие как CYP3A4 , [33] [34] особенно во время беременности , когда концентрации намного выше, чем обычно. [35] Было обнаружено, что женщины в перименопаузе имеют более высокую активность CYP3A4 по сравнению с мужчинами и женщинами в постменопаузе, и был сделан вывод, что это может быть связано с более высокими уровнями прогестерона, присутствующими у женщин в перименопаузе. [33]
Прогестерон модулирует активность CatSper (катионных каналов спермы). потенциалзависимого Ca 2+ каналы. Поскольку яйцеклетки выделяют прогестерон, сперматозоиды могут использовать прогестерон в качестве сигнала возвращения к яйцеклеткам ( хемотаксис ). В результате было высказано предположение, что вещества, блокирующие сайт связывания прогестерона в каналах CatSper, потенциально могут использоваться в мужской контрацепции . [36] [37]
Биологическая функция [ править ]
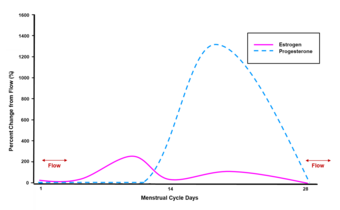
Гормональные взаимодействия [ править ]
Прогестерон обладает рядом физиологических эффектов, которые усиливаются в присутствии эстрогенов . Эстрогены через рецепторы эстрогена (ER) индуцируют или усиливают экспрессию PR . [39] Одним из примеров этого является ткань молочной железы , где эстрогены позволяют прогестерону опосредовать лобулоальвеолярное развитие. [40] [41] [42]
Повышенные уровни прогестерона сильно снижают натрий-удерживающую активность альдостерона, что приводит к натрийурезу и уменьшению объема внеклеточной жидкости. С другой стороны, отмена прогестерона связана с временным увеличением задержки натрия (снижением натрийуреза с увеличением объема внеклеточной жидкости) за счет компенсаторного увеличения продукции альдостерона, который борется с блокадой минералокортикоидных рецепторов ранее повышенным уровень прогестерона. [43]
половая Ранняя дифференциация
Прогестерон играет роль в ранней половой дифференциации человека. [44] Плацентарный прогестерон является сырьем для 5α-дигидротестостерона (ДГТ), вырабатываемого через черный ход, который действует во многих негонадных тканях плода . [45] тогда как недостатки этого пути приводят к недостаточной вирилизации плода мужского пола, что приводит к неполному развитию мужских гениталий. [46] [47] ДГТ — мощный андроген , отвечающий за развитие мужских половых органов, включая пенис и мошонку .
На ранних этапах развития плода недифференцированные гонады могут развиваться либо в семенники, либо в яичники. Наличие Y-хромосомы приводит к развитию семенников. Затем яички вырабатывают тестостерон, который преобразуется в ДГТ с помощью фермента 5α-редуктазы . ДГТ – мощный андроген, отвечающий за маскулинизацию наружных половых органов и развитие предстательной железы. Прогестерон, вырабатываемый плацентой во время беременности, играет роль в половой дифференциации плода, выступая в качестве молекулы-предшественника для синтеза ДГТ через «черный путь». При отсутствии адекватных уровней стероидогенных ферментов во время развития плода «задний путь» синтеза ДГТ может стать недостаточным, что приведет к недостаточной маскулинизации плода мужского пола. Это может привести к развитию неоднозначных гениталий или даже к появлению женских гениталий в некоторых случаях. Таким образом, и ДГТ, и прогестерон играют решающую роль в ранней половой дифференциации плода, при этом прогестерон действует как молекула-предшественник синтеза ДГТ, а ДГТ способствует развитию мужских гениталий. [44]
Репродуктивная система [ править ]

Прогестерон оказывает ключевое воздействие посредством негеномной передачи сигналов на человеческие сперматозоиды, когда они мигрируют по женским путям до того, как происходит оплодотворение , хотя рецептор(ы) пока остаются неидентифицированными. [48] Детальная характеристика событий, происходящих в сперме в ответ на прогестерон, прояснила некоторые события, включая транзиторные изменения внутриклеточного кальция и поддерживающие изменения. [49] медленные колебания кальция, [50] теперь считается, что он, возможно, регулирует моторику. [51] Его производят яичники. [52] Также было показано, что прогестерон оказывает воздействие на сперматозоиды осьминога. [53]
Прогестерон иногда называют «гормоном беременности ». [54] и он выполняет множество ролей, связанных с развитием плода:
- Прогестерон переводит эндометрий в секреторную стадию, чтобы подготовить матку к имплантации. В то же время прогестерон воздействует на вагинальный эпителий и цервикальную слизь , делая ее густой и непроницаемой для сперматозоидов . Прогестерон оказывает антимитогенное действие на эпителиальные клетки эндометрия и, как таковой, смягчает тропные эффекты эстрогена . [55] Если беременность не наступает, уровень прогестерона снизится, что приведет к менструации . Нормальное менструальное кровотечение – это кровотечение отмены прогестерона. Если овуляция не происходит и желтое тело не развивается, уровень прогестерона может быть низким, что приводит к ановуляторному дисфункциональному маточному кровотечению.
- Во время имплантации и беременности прогестерон, по-видимому, снижает материнский иммунный ответ, позволяя принять беременность. [56]
- матки Прогестерон снижает сократимость гладких мышц . [54] Этот эффект способствует профилактике преждевременных родов . [56] Исследования показали, что у женщин, беременных одним плодом, бессимптомных на пренатальной стадии и имеющих высокий риск самопроизвольных преждевременных родов, было обнаружено, что вагинальные препараты прогестерона эффективны для предотвращения спонтанных преждевременных родов. К группе высокого риска самопроизвольных преждевременных родов относятся женщины, у которых шейка матки коротка менее 25 мм или ранее спонтанно рожали преждевременно. Хотя обычно считается, что преждевременные роды происходят на сроке менее 37 недель, эти исследования показали, что вагинальный прогестерон связан с меньшим количеством преждевременных родов на сроке менее 34 недель. [57]
- Падение уровня прогестерона, возможно, является одним из шагов, способствующих началу родов .
- Кроме того, прогестерон подавляет лактацию во время беременности. Падение уровня прогестерона после родов является одним из триггеров выработки молока.
Плод плацентарный прогестерон метаболизирует при производстве стероидов надпочечников . [45]
Грудь [ править ]
развитие Лобулоальвеолярное
Прогестерон играет важную роль в развитии груди у женщин. В сочетании с пролактином он опосредует лобулоальвеолярное созревание молочных желез во время беременности, обеспечивая выработку молока и, следовательно, и грудное вскармливание потомства лактацию после родов (родов). [58] Эстроген индуцирует экспрессию PR в ткани молочной железы и, следовательно, прогестерон зависит от эстрогена, опосредуя лобулоальвеолярное развитие. [40] [41] [42] Было обнаружено, что активатор RANKL является критическим последующим медиатором индуцированного прогестероном лобулоальвеолярного созревания. [59] по RANKL, Мыши, нокаутные демонстрируют почти идентичный фенотип молочных желез с мышами, нокаутными по PR, включая нормальное развитие протоков молочных желез, но полное отсутствие развития лобулоальвеолярных структур. [59]
протоков Развитие
Хотя и в гораздо меньшей степени, чем эстроген, который является основным медиатором развития протоков молочной железы (через ERα ), [60] [61] прогестерон также может в некоторой степени участвовать в развитии протоков молочных желез. [62] У мышей с нокаутом PR или мышей, получавших антагонист PR мифепристон, наблюдается задержка, хотя в остальном нормальное развитие молочных протоков в период полового созревания. [62] Кроме того, мыши, модифицированные для PRA сверхэкспрессии , демонстрируют гиперплазию протоков. [59] а прогестерон индуцирует рост протоков молочной железы мыши. [62] Прогестерон опосредует развитие протоков главным образом посредством индукции экспрессии амфирегулина , , того же фактора роста экспрессию которого в первую очередь индуцирует эстроген, опосредующего развитие протоков. [62] Эти данные на животных позволяют предположить, что, хотя прогестерон и не является необходимым для полного развития протоков молочной железы, он, по-видимому, играет потенцирующую или ускоряющую роль в опосредованном эстрогеном развитии протоков молочной железы. [62]
рака Риск железы молочной
Прогестерон также, по-видимому, участвует в патофизиологии рака молочной железы , хотя его роль и является ли он промотором или ингибитором риска рака молочной железы, полностью не выяснены. [63] [64] Было обнаружено, что большинство прогестинов или синтетических прогестагенов, таких как медроксипрогестерона ацетат , увеличивают риск рака молочной железы у женщин в постменопаузе в сочетании с эстрогеном в качестве компонента гормональной терапии в менопаузе . [65] [64] Комбинация природного перорального прогестерона или атипичного прогестина дидрогестерона с эстрогеном связана с меньшим риском рака молочной железы, чем прогестины плюс эстроген. [66] [67] [68] Однако это может быть просто артефактом низкого уровня прогестерона, вырабатываемого пероральным прогестероном. [63] [69] Необходимы дополнительные исследования роли прогестерона при раке молочной железы. [64]
Здоровье кожи [ править ]
Рецептор эстрогена , как и рецептор прогестерона , обнаружен в коже , в том числе в кератиноцитах и фибробластах . [70] [71] В период менопаузы и в последующий период снижение уровня женских половых гормонов приводит к атрофии , истончению и усилению морщинистости кожи, а также к снижению эластичности , упругости и прочности кожи. [70] [71] ускорение старения кожи и являются результатом снижения содержания коллагена , нарушений морфологии клеток , Эти изменения кожи представляют собой эпидермиса уменьшения основного вещества между волокнами кожи , а также уменьшения капилляров и кровотока . [70] [71] Кожа также становится более сухой во время менопаузы, что связано с уменьшением гидратации кожи и поверхностных липидов (выработка кожного сала). [70] Наряду с хронологическим старением и фотостарением дефицит эстрогенов в климактерическом периоде является одним из трех основных факторов, преимущественно влияющих на старение кожи. [70]
Заместительная гормональная терапия, состоящая из системного лечения эстрогеном отдельно или в сочетании с прогестагеном, оказывает хорошо документированное и значительное благоприятное воздействие на кожу женщин в постменопаузе. [70] [71] Эти преимущества включают увеличение содержания коллагена в коже, увеличение толщины и эластичности кожи, а также увлажнение кожи и поверхностных липидов. [70] [71] Было обнаружено, что местный эстроген оказывает аналогичное благотворное воздействие на кожу. [70] Кроме того, исследование показало, что местный крем с 2% прогестерона значительно повышает эластичность и упругость кожи, а также заметно уменьшает морщины у женщин в пери- и постменопаузе. [71] С другой стороны, гидратация кожи и поверхностные липиды существенно не изменились при местном применении прогестерона. [71]
Эти данные свидетельствуют о том, что прогестерон, как и эстроген, также оказывает благотворное воздействие на кожу и может независимо защищать ее от старения. [71]
Сексуальность [ править ]
Либидо [ править ]
Прогестерон и его нейростероидный активный метаболит аллопрегнанолон, по-видимому, играют важную роль в либидо у женщин. [72]
Гомосексуализм [ править ]
Доктор Диана Флейшман из Портсмутского университета и ее коллеги изучали связь между прогестероном и сексуальным поведением у 92 женщин. Их исследование, опубликованное в « Архивах сексуального поведения», показало, что женщины с более высоким уровнем прогестерона набрали более высокие баллы в анкете, измеряющей гомоэротическую мотивацию. Они также обнаружили, что мужчины с высоким уровнем прогестерона с большей вероятностью имели более высокие показатели гомоэротической мотивации после аффилиативного прайминга по сравнению с мужчинами с низким уровнем прогестерона. [73] [74] [75] [76]
Нервная система [ править ]
Прогестерон, как и прегненолон и дегидроэпиандростерон (ДГЭА), принадлежит к важной группе эндогенных стероидов, называемых нейростероидами . Он может метаболизироваться во всех отделах центральной нервной системы . [77]
Нейростероиды являются нейромодуляторами , обладают нейропротекторным , нейрогенным действием и регулируют нейротрансмиссию и миелинизацию . [78] Эффекты прогестерона как нейростероида опосредованы преимущественно через его взаимодействие с неядерными PR, а именно с mPR и PGRMC1 , а также с некоторыми другими рецепторами, такими как рецепторы σ1 и nACh. [79]
Повреждение головного мозга [ править ]
Предыдущие исследования показали, что прогестерон поддерживает нормальное развитие нейронов головного мозга и оказывает защитное действие на поврежденную ткань мозга. На животных моделях наблюдалось, что у самок снижена восприимчивость к черепно-мозговым травмам , и было высказано предположение, что этот защитный эффект вызван повышенным уровнем циркулирующих эстрогена и прогестерона у самок. [80]
Предлагаемый механизм [ править ]
Механизм защитного действия прогестерона может заключаться в уменьшении воспаления, возникающего после травмы головного мозга и кровоизлияния. [81] [82]
Считается, что повреждения, вызванные черепно-мозговой травмой, частично вызваны массовой деполяризацией, приводящей к эксайтотоксичности . Одним из способов, с помощью которого прогестерон помогает частично облегчить эксайтотоксичность, является блокирование потенциал-зависимых кальциевых каналов , которые запускают высвобождение нейромедиаторов . [83] Это происходит путем манипулирования сигнальными путями факторов транскрипции, участвующих в этом высвобождении. Другой метод снижения эксайтотоксичности — активация ГАМК А , широко распространенного тормозного рецептора нейротрансмиттера. [84]
Также было показано, что прогестерон предотвращает апоптоз нейронов, частое последствие травмы головного мозга. [85] Это происходит путем ингибирования ферментов, участвующих в пути апоптоза, особенно в отношении митохондрий, таких как активированная каспаза 3 и цитохром с .
Прогестерон не только помогает предотвратить дальнейшее повреждение, но также способствует нейрорегенерации . [86] Одним из серьезных последствий черепно-мозговой травмы является отек. Исследования на животных показывают, что лечение прогестероном приводит к снижению уровня отеков за счет увеличения концентрации макрофагов и микроглии, направляемых в поврежденную ткань. [83] [87] Это наблюдалось в виде снижения утечки через гематоэнцефалический барьер при вторичном восстановлении у крыс, получавших прогестерон. Кроме того, было обнаружено, что прогестерон обладает антиоксидантными свойствами, снижая концентрацию свободных радикалов кислорода быстрее, чем без него. [84] Есть также доказательства того, что добавление прогестерона также может помочь повторно миелинизировать поврежденные аксоны, в результате травмы, восстанавливая часть утраченной проводимости нервного сигнала. [84] Другой способ, которым прогестерон способствует регенерации, включает увеличение циркуляции эндотелиальных клеток-предшественников в головном мозге. [88] Это помогает новой сосудистой сети разрастаться вокруг рубцовой ткани, что помогает восстановить область повреждения.
Наркомания [ править ]
Прогестерон усиливает функцию рецепторов серотонина в головном мозге, поэтому избыток или дефицит прогестерона может привести к серьезным нейрохимическим проблемам. Это объясняет, почему некоторые люди прибегают к веществам, повышающим активность серотонина , таким как никотин , алкоголь и каннабис , когда уровень прогестерона падает ниже оптимального уровня. [89]
- Половые различия в уровнях гормонов могут побудить женщин реагировать на никотин иначе, чем мужчин. Когда женщины претерпевают циклические изменения или различные гормональные переходные фазы (менопауза, беременность, подростковый возраст), у них происходят изменения уровня прогестерона. [90] Таким образом, женщины имеют повышенную биологическую уязвимость к усиливающему эффекту никотина по сравнению с мужчинами, и для противодействия этой повышенной уязвимости можно использовать прогестерон. Эта информация подтверждает идею о том, что прогестерон может влиять на поведение. [89]
- Подобно никотину, кокаин также увеличивает высвобождение дофамина в мозге. Нейромедиатор участвует в работе центра вознаграждения и является одним из основных нейромедиаторов, участвующих в злоупотреблении психоактивными веществами и зависимости. Исследование, проведенное среди потребителей кокаина, показало, что прогестерон уменьшает тягу и чувство возбуждения от кокаина. Таким образом, прогестерон был предложен в качестве средства, снижающего тягу к кокаину за счет снижения дофаминергических свойств препарата. [91]
Социальный [ править ]
В исследовании 2012 года, проведенном Амстердамским университетом среди 120 женщин, лютеиновая фаза у женщин (более высокий уровень прогестерона и повышение уровня эстрогена) коррелировала с более низким уровнем соревновательного поведения в сценариях азартных игр и математических соревнований, в то время как их предменструальная фаза (резко снижающаяся) уровень прогестерона и снижение уровня эстрогена) коррелировало с более высоким уровнем конкурентного поведения. [92]
Другие эффекты [ править ]
- Прогестерон также играет роль в эластичности кожи и прочности костей, в дыхании , в нервной ткани и в женской сексуальности , а присутствие рецепторов прогестерона в определенных мышечных и жировых тканях может указывать на роль в половых диморфных пропорциях. их [93] [ ссылка, нарушающая права? ]
- Говорят, что во время беременности прогестерон уменьшает раздражительность матки. [94]
- Во время беременности прогестерон способствует подавлению иммунных реакций матери на антигены плода, что предотвращает отторжение плода. [94]
- Прогестерон повышает уровень эпидермального фактора роста-1 (EGF-1), фактора, который часто используется для индукции пролиферации и поддержания культур стволовых клеток . [95]
- Прогестерон повышает внутреннюю температуру (термогенную функцию) во время овуляции. [96] [97]
- Прогестерон уменьшает спазмы и расслабляет гладкие мышцы . Бронхи расширены, выделение слизи регулируется. (PR широко присутствуют в подслизистой ткани .)
- Прогестерон действует как противовоспалительное средство и регулирует иммунный ответ .
- Прогестерон снижает активность желчного пузыря . [98]
- Прогестерон нормализует свертываемость крови и тонус сосудов, уровень цинка и меди , в клетках уровень кислорода и использование жировых запасов для получения энергии.
- Прогестерон может повлиять на здоровье десен, увеличивая риск гингивита (воспаления десен). [99]
- Прогестерон, по-видимому, предотвращает рак эндометрия (поражающий слизистую оболочку матки), регулируя действие эстрогена.
- Прогестерон играет важную роль в передаче сигналов о высвобождении инсулина и функции поджелудочной железы и может влиять на предрасположенность к диабету или гестационному диабету. [100] [101]
- Было обнаружено, что уровень прогестерона в крови ниже у женщин с более высоким весом и более высоким ИМТ среди тех, кто забеременел в результате экстракорпорального оплодотворения. [102]
- Современные данные показывают, что микронизированный прогестерон, который химически идентичен прогестерону, вырабатываемому в женском организме, в сочетании с эстрогеном при менопаузальной гормональной терапии, по-видимому, не оказывает существенного влияния на венозную тромбоэмболию (сгустки крови в венах) и ишемический инсульт (недостаток крови). приток к мозгу из-за закупорки кровеносного сосуда, снабжающего мозг). Однако необходимо провести дополнительные исследования, чтобы выяснить, изменяет ли микронизированный прогестерон отдельно или в комбинированной гормональной терапии в период менопаузы риск инфарктов миокарда (сердечных приступов). [103]
- Пока не проводилось никаких исследований по влиянию микронизированного прогестерона на выпадение волос в результате менопаузы. [104]
- Несмотря на предложения по использованию гормональной терапии для предотвращения потери мышечной массы у женщин в постменопаузе (50 лет и старше), менопаузальная гормональная терапия, включающая либо только эстроген, либо эстроген и прогестерон, не способствует сохранению мышечной массы. [105] Менопаузальная гормональная терапия также не приводит к снижению массы тела, снижению ИМТ или изменению метаболизма глюкозы. [106]
Биохимия [ править ]
Биосинтез [ править ]
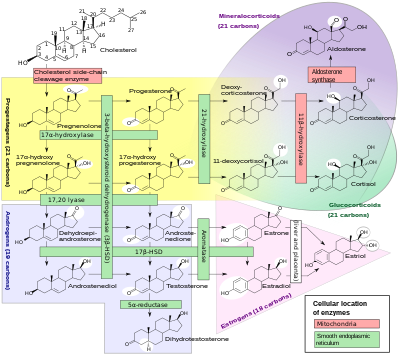
У млекопитающих прогестерон, как и все другие стероидные гормоны , синтезируется из прегненолона , который сам является производным холестерина .
Холестерин подвергается двойному окислению с образованием 22R - гидроксихолестерина , а затем 20α,22R - дигидроксихолестерина . Этот вицинальный диол затем дополнительно окисляется с потерей боковой цепи, начиная с положения C22, с образованием прегненолона. Эту реакцию катализирует цитохром P450scc .
The conversion of pregnenolone to progesterone takes place in two steps. First, the 3β-hydroxyl group is oxidized to a keto group and second, the double bond is moved to C4, from C5 through a keto/enol tautomerization reaction.[108] This reaction is catalyzed by 3β-hydroxysteroid dehydrogenase/δ5-4-isomerase.
Progesterone in turn is the precursor of the mineralocorticoid aldosterone, and after conversion to 17α-hydroxyprogesterone, of cortisol and androstenedione. Androstenedione can be converted to testosterone, estrone, and estradiol, highlighting the critical role of progesterone in testosterone synthesis.
Pregnenolone and progesterone can also be synthesized by yeast.[109]
Approximately 25 mg of progesterone is secreted from the ovaries per day in women, while the adrenal glands produce about 2 mg of progesterone per day.[110]
Distribution[edit]
Progesterone binds extensively to plasma proteins, including albumin (50–54%) and transcortin (43–48%).[111] It has similar affinity for albumin relative to the PR.[17]
Metabolism[edit]
The metabolism of progesterone is rapid and extensive and occurs mainly in the liver,[112][113][114] though enzymes that metabolize progesterone are also expressed widely in the brain, skin, and various other extrahepatic tissues.[77][115] Progesterone has an elimination half-life of only approximately 5 minutes in circulation.[112] The metabolism of progesterone is complex, and it may form as many as 35 different unconjugated metabolites when it is ingested orally.[114][116] Progesterone is highly susceptible to enzymatic reduction via reductases and hydroxysteroid dehydrogenases due to its double bond (between the C4 and C5 positions) and its two ketones (at the C3 and C20 positions).[114]
The major metabolic pathway of progesterone is reduction by 5α-reductase[77] and 5β-reductase into the dihydrogenated 5α-dihydroprogesterone and 5β-dihydroprogesterone, respectively.[113][114][117][118] This is followed by the further reduction of these metabolites via 3α-hydroxysteroid dehydrogenase and 3β-hydroxysteroid dehydrogenase into the tetrahydrogenated allopregnanolone, pregnanolone, isopregnanolone, and epipregnanolone.[119][113][114][117] Subsequently, 20α-hydroxysteroid dehydrogenase and 20β-hydroxysteroid dehydrogenase reduce these metabolites to form the corresponding hexahydrogenated pregnanediols (eight different isomers in total),[113][118] which are then conjugated via glucuronidation and/or sulfation, released from the liver into circulation, and excreted by the kidneys into the urine.[112][114] The major metabolite of progesterone in the urine is the 3α,5β,20α isomer of pregnanediol glucuronide, which has been found to constitute 15 to 30% of an injection of progesterone.[17][120] Other metabolites of progesterone formed by the enzymes in this pathway include 3α-dihydroprogesterone, 3β-dihydroprogesterone, 20α-dihydroprogesterone, and 20β-dihydroprogesterone, as well as various combination products of the enzymes aside from those already mentioned.[17][114][120][121] Progesterone can also first be hydroxylated (see below) and then reduced.[114] Endogenous progesterone is metabolized approximately 50% into 5α-dihydroprogesterone in the corpus luteum, 35% into 3β-dihydroprogesterone in the liver, and 10% into 20α-dihydroprogesterone.[122]
Relatively small portions of progesterone are hydroxylated via 17α-hydroxylase (CYP17A1) and 21-hydroxylase (CYP21A2) into 17α-hydroxyprogesterone and 11-deoxycorticosterone (21-hydroxyprogesterone), respectively,[116] and pregnanetriols are formed secondarily to 17α-hydroxylation.[123][124] Even smaller amounts of progesterone may be also hydroxylated via 11β-hydroxylase (CYP11B1) and to a lesser extent via aldosterone synthase (CYP11B2) into 11β-hydroxyprogesterone.[125][126][44] In addition, progesterone can be hydroxylated in the liver by other cytochrome P450 enzymes which are not steroid-specific.[127] 6β-Hydroxylation, which is catalyzed mainly by CYP3A4, is the major transformation, and is responsible for approximately 70% of cytochrome P450-mediated progesterone metabolism.[127] Other routes include 6α-, 16α-, and 16β-hydroxylation.[114] However, treatment of women with ketoconazole, a strong CYP3A4 inhibitor, had minimal effects on progesterone levels, producing only a slight and non-significant increase, and this suggests that cytochrome P450 enzymes play only a small role in progesterone metabolism.[128]
Metabolism of progesterone in humans[129] |

Levels[edit]

In women, progesterone levels are relatively low during the preovulatory phase of the menstrual cycle, rise after ovulation, and are elevated during the luteal phase, as shown in the diagram above. Progesterone levels tend to be less than 2 ng/mL prior to ovulation and greater than 5 ng/mL after ovulation. If pregnancy occurs, human chorionic gonadotropin is released, maintaining the corpus luteum and allowing it to maintain levels of progesterone. Between 7 and 9 weeks, the placenta begins to produce progesterone in place of the corpus luteum in a process called the luteal-placental shift.[131]
After the luteal-placental shift, progesterone levels start to rise further and may reach 100 to 200 ng/mL at term. Whether a decrease in progesterone levels is critical for the initiation of labor has been argued and may be species-specific. After delivery of the placenta and during lactation, progesterone levels are very low.
Progesterone levels are low in children and postmenopausal women.[132] Adult males have levels similar to those in women during the follicular phase of the menstrual cycle.
| Group | P4 production | P4 levels | ||
|---|---|---|---|---|
| Prepubertal children | ND | 0.06–0.5 ng/mL | ||
| Pubertal girls Tanner stage I (childhood) Tanner stage II (ages 8–12) Tanner stage III (ages 10–13) Tanner stage IV (ages 11–14) Tanner stage V (ages 12–15) Follicular phase (days 1–14) Luteal phase (days 15–28) | ND ND ND ND ND ND | 0.22 (<0.10–0.32) ng/mL 0.30 (0.10–0.51) ng/mL 0.36 (0.10–0.75) ng/mL 1.75 (<0.10–25.0) ng/mL 0.35 (0.13–0.75) ng/mL 2.0–25.0 ng/mL | ||
| Premenopausal women Follicular phase (days 1–14) Luteal phase (days 15–28) Oral contraceptive (anovulatory) | 0.75–5.4 mg/day 15–50 mg/day ND | 0.02–1.2 ng/mL 4–30 ng/mL 0.1–0.3 ng/mL | ||
| Postmenopausal women Oophorectomized women Oophorectomized and adrenalectomized women | ND 1.2 mg/day <0.3 mg/day | 0.03–0.3 ng/mL 0.39 ng/mL ND | ||
| Pregnant women First trimester (weeks 1–12) Second trimester (weeks 13–26) Third trimester (weeks 27–40) Postpartum (at 24 hours) | 55 mg/day 92–100 mg/day 190–563 mg/day ND | 9–75 ng/mL 17–146 ng/mL 55–255 ng/mL 19 ng/mL | ||
| Men | 0.75–3 mg/day | 0.1–0.3 ng/mL | ||
| Notes: Mean levels are given as a single value and ranges are given after in parentheses. Sources: [129][133][134][135][136][137][138][139][140] | ||||
Ranges[edit]
Blood test results should always be interpreted using the reference ranges provided by the laboratory that performed the results. Example reference ranges are listed below.
| Person type | Reference range for blood test | ||
|---|---|---|---|
| Lower limit | Upper limit | Unit | |
| Female - menstrual cycle | (see diagram below) | ||
| Female - postmenopausal | <0.2[141] | 1[141] | ng/mL |
| <0.6[142] | 3[142] | nmol/L | |
| Female on oral contraceptives | 0.34[141] | 0.92[141] | ng/mL |
| 1.1[142] | 2.9[142] | nmol/L | |
| Males ≥16 years | 0.27[141] | 0.9[141] | ng/mL |
| 0.86[142] | 2.9[142] | nmol/L | |
| Female or male 1–9 years | 0.1[141] | 4.1[141] or 4.5[141] | ng/mL |
| 0.3[142] | 13[142] | nmol/L | |

Источники [ править ]
Животное [ править ]
Прогестерон вырабатывается в больших количествах в яичниках (желтым телом ) от начала полового созревания до менопаузы , а также в меньших количествах вырабатывается надпочечниками после начала адренархе как у мужчин, так и у женщин. В меньшей степени прогестерон вырабатывается в нервной ткани , особенно в головном мозге, а в жировой (жировой) ткани также .
Во время беременности прогестерон вырабатывается во все больших количествах яичниками и плацентой . Сначала источником является желтое тело, которое было «спасено» наличием хорионического гонадотропина человека (ХГЧ) из зачатия. Однако после 8-й недели производство прогестерона перемещается в плаценту. Плацента использует материнский холестерин в качестве исходного субстрата, и большая часть вырабатываемого прогестерона попадает в кровоток матери, но некоторая часть попадает в кровообращение плода и используется в качестве субстрата для кортикостероидов плода. В срок плацента вырабатывает около 250 мг прогестерона в день.
Дополнительным животным источником прогестерона являются молочные продукты. После употребления молочных продуктов уровень биодоступного прогестерона повышается. [144]
Растения [ править ]
По крайней мере, в одном растении Juglans regia был обнаружен прогестерон. [145] прогестеронподобные стероиды обнаружены Кроме того, в Dioscorea mexicana . Dioscorea mexicana — растение семейства ямсовых , произрастающее в Мексике . [146] Он содержит стероид диосгенин , который извлекается из растения и превращается в прогестерон. [147] Диосгенин и прогестерон также обнаружены в других видах Dioscorea , а также в других растениях, не являющихся близкими родственниками, таких как пажитник .
Еще одно растение, содержащее вещества, легко превращаемые в прогестерон, — это Dioscorea pseudojaponica, произрастающая на Тайване . Исследования показали, что тайваньский ямс содержит сапонины — стероиды, которые могут превращаться в диосгенин, а затем в прогестерон. [148]
Многие другие виды Dioscorea семейства ямсовых содержат стероидные вещества, из которых может вырабатываться прогестерон. Среди наиболее примечательных из них — Dioscorea villosa и Dioscorea polygonoides . Одно исследование показало, что Dioscorea villosa содержит 3,5% диосгенина. [149] Dioscorea Polygonoides Было обнаружено, что содержит 2,64% диосгенина, как показано методом газовой хроматографии-масс-спектрометрии . [150] Многие виды Dioscorea , происходящие из семейства ямсовых, растут в странах с тропическим и субтропическим климатом. [151]
Медицинское использование
Прогестерон используется в качестве лекарства . Он используется в сочетании с эстрогенами, в гормональной терапии симптомов менопаузы главным образом , и низкого уровня половых гормонов у женщин. [116] [152] Его также можно использовать отдельно для лечения симптомов менопаузы. Исследования показали, что трансдермальный прогестерон (кожный пластырь) и пероральный микронизированный прогестерон являются эффективными средствами лечения определенных симптомов менопаузы, таких как приливы жара и ночная потливость, которые иначе называются вазомоторными симптомами или ВМС. [153]
Он также используется у женщин для поддержания беременности и фертильности , а также для лечения гинекологических заболеваний . [154] [155] [156] [157] Было показано, что прогестерон предотвращает выкидыш у женщин с 1) вагинальным кровотечением на ранних этапах текущей беременности и 2) выкидышем в анамнезе. [158] Прогестерон можно принимать перорально , через влагалище , инъекцией в мышцы или жир и другими способами . [116]
Химия [ править ]

Прогестерон — это природный прегнановый стероид , также известный как прегн-4-ен-3,20-дион. [159] [160] Он имеет двойную связь ( 4-ен ) между положениями C4 и C5 и две кетоновые группы (3,20- дион ), одну в положении C3, а другую в положении C20. [159] [160]
Синтез [ править ]
Прогестерон в промышленных масштабах производится полусинтезом. Используются два основных пути: один из диосгенина ямса , впервые использованный Маркером в 1940 году, и другой, основанный на соевых фитостеролах, масштабы применения которых были увеличены в 1970-х годах. Сообщалось также о дополнительных (не обязательно экономичных) полусинтезах прогестерона, начиная с различных стероидов. Например, кортизон можно одновременно дезоксигенировать в положениях C-17 и C-21 путем обработки йодтриметилсиланом в хлороформе с образованием 11-кетопрогестерона (кетогестина), который, в свою очередь, может быть восстановлен в положении-11 с получением прогестерона. [161]
Полусинтез маркера [ править ]
Экономичный полусинтез прогестерона из растительного стероида диосгенина, выделенного из ямса, был разработан Расселом Маркером в 1940 году для фармацевтической компании Parke-Davis . [162] Этот синтез известен как деградация маркера .

Промежуточный продукт 16-ДФА важен для синтеза многих других важных с медицинской точки зрения стероидов. Очень похожий подход позволяет получить 16-ДФА из соланина . [163]
Полусинтез сои [ править ]
Прогестерон также можно производить из стигмастерола , содержащегося в соевом масле . см . Перси Джулиан .

Полный синтез [ править ]

О полном синтезе прогестерона сообщил в 1971 году У.С. Джонсон . [169] Синтез начинается с реакции фосфониевой соли 7 с фениллитием с образованием илида фосфония 8 . Илид 8 реагирует с альдегидом с образованием алкена 9 . Кетальные , который , защитные группы 9 в свою очередь , гидролизуются с образованием дикетона 10 циклизуется с образованием циклопентенона 11 . Кетон 11 реагирует с метиллитием с образованием третичного спирта 12 , который, в свою очередь, обрабатывается кислотой с образованием третичного катиона 13 . Ключевой стадией синтеза является π-катионная циклизация 13 , при которой одновременно образуются B-, C- и D-кольца стероида с образованием 14 . Этот этап напоминает реакцию катионной циклизации, используемую при биосинтезе стероидов, и поэтому его называют биомиметическим . На следующем этапе енола ортоэфир гидролизуется с образованием кетона 15 . Затем циклопентеновое А-кольцо раскрывается путем окисления озоном с образованием 16 . Наконец, дикетон 17 подвергается внутримолекулярной альдольной конденсации при обработке водным гидроксидом калия с образованием прогестерона. [169]
История [ править ]
Джордж Корнер и Уиллард М. Аллен открыли гормональное действие прогестерона в 1929 году. [17] [170] [171] [172] животных был выделен практически чистый кристаллический материал с высокой прогестеронной активностью К 1931–1932 гг. из желтого тела , а к 1934 г. был очищен и получен чистый кристаллический прогестерон и химическая структура прогестерона. определена [17] [171] Этого добился Адольф Бутенандт из Химического института в технического университета Данциге , который извлек это новое соединение из нескольких тысяч литров мочи . [173]
Позже в том же году был завершен химический синтез прогестерона из стигмастерола и прегнандиола . [171] [174] До этого момента прогестерон, известный под общим названием гормон желтого тела, назывался несколькими группами под разными названиями, включая корпорин, лютеин, лютеостерон и прогестин. [17] [175] В 1935 году, во время Второй Международной конференции по стандартизации половых гормонов в Лондоне, Англия , между группами был достигнут компромисс и было создано название прогестерон (гестационный стероидный кетон). [17] [176]
Ветеринарное использование [ править ]
Использование тестов на прогестерон в собаководстве для определения овуляции становится все более широко распространенным. Существует несколько тестов, но наиболее надежным тестом является анализ крови, взятый ветеринаром и отправленный в лабораторию для обработки. Результаты обычно можно получить через 24–72 часа. Обоснованием использования тестов на прогестерон является то, что увеличение количества гонадотропинов начинается в непосредственной близости от предовуляторного всплеска гонадотропинов и продолжается во время овуляции и течки. Когда уровень прогестерона достигает определенного уровня, это может сигнализировать о стадии течки у самки. Предсказание даты рождения будущего помета может быть очень точным, если известна дата овуляции. В большинстве случаев щенки рожают на день или два из 9 недель беременности. Однако после спаривания невозможно определить беременность с помощью тестов на прогестерон. Это связано с тем, что у собак уровень прогестерона остается повышенным на протяжении всего периода течки. [177]
Цены [ править ]
Цены на прогестерон могут варьироваться в зависимости от местоположения, страхового покрытия, купонов на скидку, количества, дефицита, производителей, брендовых или генерических версий, различных аптек и так далее. На данный момент 30 капсул по 100 мг непатентованной версии Prometrium от CVS Pharmacy стоят около 40 долларов США без каких-либо скидок или страховки. Фирменная версия «Прогестерон» стоит около 450 долларов за 30 капсул без каких-либо скидок или страховки. [178] Для сравнения, Walgreens предлагает 30 капсул по 100 мг в непатентованной версии за 51 доллар без страховки или купонов. Торговая марка стоит около 431 доллара за 30 капсул по 100 мг. [179]
Ссылки [ править ]
 В эту статью включен текст , доступный по лицензии CC BY-SA 3.0 .
В эту статью включен текст , доступный по лицензии CC BY-SA 3.0 .
- ^ Jump up to: Перейти обратно: а б Джеймсон Дж.Л., Де Гроот LJ (25 февраля 2015 г.). Эндокринология: электронная книга для взрослых и детей . Elsevier Науки о здоровье. п. 2179. ИСБН 978-0-323-32195-2 . Архивировано из оригинала 14 января 2023 года . Проверено 29 октября 2017 г.
- ^ Адлер Н., Пфафф Д., Гой Р.В. (6 декабря 2012 г.). Справочник по поведенческой нейробиологии, том 7. Воспроизведение (1-е изд.). Нью-Йорк: Пленум Пресс. п. 189. ИСБН 978-1-4684-4834-4 . Архивировано из оригинала 14 января 2023 года . Проверено 4 июля 2015 г.
- ^ «прогестерон (CHEBI:17026)» . ЧЭБИ . Европейская лаборатория молекулярной биологии-EBI. Архивировано из оригинала 20 марта 2016 года . Проверено 4 июля 2015 г.
- ^ «Прогестерон_мсдс» . Архивировано из оригинала 12 февраля 2021 года . Проверено 19 апреля 2018 г.
- ^ Jump up to: Перейти обратно: а б Станчик ФЗ (сентябрь 2002 г.). «Фармакокинетика и эффективность прогестинов, используемых для заместительной гормональной терапии и контрацепции». Обзоры по эндокринным и метаболическим расстройствам . 3 (3): 211–224. дои : 10.1023/А:1020072325818 . ПМИД 12215716 . S2CID 27018468 .
- ^ Jump up to: Перейти обратно: а б с Саймон Дж.А., Робинсон Д.Э., Эндрюс М.К., Хильдебранд-младший, Рокки М.Л., Блейк Р.Э., Ходген Г.Д. (июль 1993 г.). «Абсорбция перорального микронизированного прогестерона: влияние еды, пропорциональность дозы и сравнение с внутримышечным прогестероном» . Фертильность и бесплодие . 60 (1): 26–33. дои : 10.1016/S0015-0282(16)56031-2 . ПМИД 8513955 .
- ^ Фриц М.А., Сперофф Л. (28 марта 2012 г.). Клиническая гинекологическая эндокринология и бесплодие . Липпинкотт Уильямс и Уилкинс. стр. 44–. ISBN 978-1-4511-4847-3 .
- ^ Маршалл В.Дж., Бангерт С.К. (2008). Клиническая химия . Elsevier Науки о здоровье. стр. 192–. ISBN 978-0-7234-3455-9 . Архивировано из оригинала 14 января 2023 года . Проверено 5 октября 2016 г.
- ^ Ямадзаки Х., Симада Т. (октябрь 1997 г.). «Гидроклирование прогестерона и тестостерона цитохромами P450 2C19, 2C9 и 3A4 в микросомах печени человека». Архив биохимии и биофизики . 346 (1): 161–169. дои : 10.1006/abbi.1997.0302 . ПМИД 9328296 .
- ^ Маккей Г.А., Уолтерс М.Р. (6 февраля 2013 г.). Конспект лекций: Клиническая фармакология и терапия . Джон Уайли и сыновья. п. 33. ISBN 978-1-118-34489-7 . Архивировано из оригинала 14 января 2023 года . Проверено 27 июня 2015 г.
- ^ Зутши В., Ратор А.М., Шарма К. (1 января 2005 г.). Гормоны в акушерстве и гинекологии . Издательство Jaypee Brothers. п. 74. ИСБН 978-81-8061-427-9 . [ постоянная мертвая ссылка ]
- ^ Jump up to: Перейти обратно: а б Кометти Б (ноябрь 2015 г.). «Фармацевтическая и клиническая разработка новой формы прогестерона» . Acta Obstetricia et Gynecologica Scandinavica . 94 (Приложение 161): 28–37. дои : 10.1111/aogs.12765 . ПМИД 26342177 . S2CID 31974637 .
- ^ Jump up to: Перейти обратно: а б с King TL, Brucker MC (25 октября 2010 г.). Фармакология для женского здоровья . Издательство Джонс и Бартлетт. стр. 372–373. ISBN 978-1-4496-5800-7 .
- ^ Jump up to: Перейти обратно: а б Болье Э., Шумахер М. (2000). «Прогестерон как нейроактивный нейростероид, с особым упором на влияние прогестерона на миелинизацию». Стероиды . 65 (10–11): 605–612. дои : 10.1016/s0039-128x(00)00173-2 . ПМИД 11108866 . S2CID 14952168 .
- ^ Приор JC (апрель 2019 г.). «Прогестерон важен для терапии женщин-трансгендеров: доказательства пользы прогестерона у цисженщин» . Журнал клинической эндокринологии и метаболизма . 104 (4): 1181–1186. дои : 10.1210/jc.2018-01777 . ПМИД 30608551 . S2CID 58620122 .
Накоплены доказательства того, что нормальный прогестерон (и овуляция), а также физиологический уровень эстрадиола необходимы во время предклимактерических менструальных циклов цис-женщин для текущей фертильности и долгосрочного здоровья; трансгендерным женщинам может потребоваться терапия прогестероном и аналогичные потенциальные физиологические преимущества.
- ^ Фишер Дж., Ганеллин С.Р. (2006). Открытие аналоговых лекарств . Джон Уайли и сыновья. п. 47Х. ISBN 9783527607495 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час Йосимович Ж.Б. (11 ноября 2013 г.). Гинекологическая эндокринология . Springer Science & Business Media. стр. 9, 25–29. ISBN 978-1-4613-2157-6 . Архивировано из оригинала 14 января 2023 года . Проверено 1 февраля 2016 года .
- ^ Томас П., Панг Ю (2012). «Мембранные рецепторы прогестерона: данные о нейропротекторных, нейростероидных сигнальных и нейроэндокринных функциях в нейрональных клетках» . Нейроэндокринология . 96 (2): 162–171. дои : 10.1159/000339822 . ПМК 3489003 . ПМИД 22687885 .
- ^ Валадес-Космес П., Васкес-Мартинес Э.Р., Сербон М., Камачо-Арройо I (октябрь 2016 г.). «Мембранные рецепторы прогестерона при репродукции и раке». Молекулярная и клеточная эндокринология . 434 : 166–175. дои : 10.1016/j.mce.2016.06.027 . ПМИД 27368976 . S2CID 3826650 .
- ^ Мейер С., Шмид Р., Шмидинг К., Фалькенштейн Э., Велинг М. (февраль 1998 г.). «Характеристика мембранных белков с высоким сродством, связывающих прогестерон, с помощью антипептидной антисыворотки». Стероиды . 63 (2): 111–116. дои : 10.1016/s0039-128x(97)00143-8 . ПМИД 9516722 . S2CID 40096058 .
- ^ Кабе Ю, Ханда Х, Суэмацу М (июль 2018 г.). «Функция и структурная регуляция мембранного белка PGRMC1, реагирующего на монооксид углерода (CO)» . Журнал клинической биохимии и питания . 63 (1): 12–17. дои : 10.3164/jcbn.17-132 . ПМК 6064819 . ПМИД 30087538 .
- ^ Рю К.С., Кляйн К., Зангер У.М. (27 марта 2017 г.). «Мембранассоциированные рецепторы прогестерона: беспорядочные белки с плейотропными функциями - фокус на взаимодействиях с цитохромами P450» . Границы в фармакологии . 8 : 159. дои : 10.3389/fphar.2017.00159 . ПМЦ 5366339 . ПМИД 28396637 .
- ^ Морис Т., Урани А., Фан В.Л., Ромье П. (ноябрь 2001 г.). «Взаимодействие нейроактивных стероидов и функции рецептора сигма1: поведенческие последствия и терапевтические возможности». Исследования мозга. Обзоры исследований мозга . 37 (1–3): 116–132. дои : 10.1016/s0165-0173(01)00112-6 . ПМИД 11744080 . S2CID 44931783 .
- ^ Йоханнессен М., Фонтанилья Д., Мавлютов Т., Руохо А.Е., Джексон М.Б. (февраль 2011 г.). «Антагонистическое действие прогестерона на σ-рецепторы при модуляции потенциалзависимых натриевых каналов» . Американский журнал физиологии. Клеточная физиология . 300 (2): C328–C337. doi : 10.1152/ajpcell.00383.2010 . ПМК 3043630 . ПМИД 21084640 .
- ^ Jump up to: Перейти обратно: а б Рупрехт Р., Реул Дж.М., ван Стинсель Б., Шпенглер Д., Седер М., Бернинг Б. и др. (октябрь 1993 г.). «Фармакологическая и функциональная характеристика лигандов минералокортикоидных и глюкокортикоидных рецепторов человека». Европейский журнал фармакологии . 247 (2): 145–154. дои : 10.1016/0922-4106(93)90072-H . ПМИД 8282004 .
- ^ Элджер В., Бейер С., Поллоу К., Гарфилд Р., Ши С.К., Хиллиш А. (ноябрь 2003 г.). «Концепция и фармакодинамический профиль дроспиренона». Стероиды . 68 (10–13): 891–905. doi : 10.1016/j.steroids.2003.08.008 . ПМИД 14667981 . S2CID 41756726 .
- ^ Аттарди Б.Дж., Железник А., Симхан Х., Чиао Дж.П., Мэттисон Д.Р., Каритис С.Н. (декабрь 2007 г.). «Сравнение связывания прогестерона и глюкокортикоидных рецепторов и стимуляции экспрессии генов прогестероном, 17-альфа-гидроксипрогестерон-капроатом и родственными прогестинами» . Американский журнал акушерства и гинекологии . 197 (6): 599.e1–599.e7. дои : 10.1016/j.ajog.2007.05.024 . ПМК 2278032 . ПМИД 18060946 .
- ^ Лей К., Чен Л., Георгиу Е.С., Сооранна С.Р., Ханджани С., Бросенс Дж.Дж. и др. (2012). «Прогестерон действует через ядерный глюкокортикоидный рецептор, подавляя IL-1β-индуцированную экспрессию ЦОГ-2 в клетках миометрия человека» . ПЛОС ОДИН . 7 (11): e50167. Бибкод : 2012PLoSO...750167L . дои : 10.1371/journal.pone.0050167 . ПМК 3509141 . ПМИД 23209664 .
- ^ Пол С.М., Перди Р.Х. (март 1992 г.). «Нейроактивные стероиды» . Журнал ФАСЭБ . 6 (6): 2311–2322. дои : 10.1096/fasebj.6.6.1347506 . ПМИД 1347506 . S2CID 221753076 .
- ^ Кливер С.А., Гудвин Б., Уилсон Т.М. (октябрь 2002 г.). «Ядерный рецептор прегнана X: ключевой регулятор метаболизма ксенобиотиков» . Эндокринные обзоры . 23 (5): 687–702. дои : 10.1210/er.2001-0038 . ПМИД 12372848 .
- ^ Леманн Дж.М., Макки Д.Д., Уотсон М.А., Уилсон Т.М., Мур Дж.Т., Кливер С.А. (сентябрь 1998 г.). «Осиротный ядерный рецептор человека PXR активируется соединениями, которые регулируют экспрессию гена CYP3A4 и вызывают лекарственное взаимодействие» . Журнал клинических исследований . 102 (5): 1016–1023. дои : 10.1172/JCI3703 . ПМК 508967 . ПМИД 9727070 .
- ^ Минвелл Н.А. (8 декабря 2014 г.). Тактика в современном дизайне лекарств . Спрингер. стр. 161–. ISBN 978-3-642-55041-6 . Архивировано из оригинала 14 января 2023 года . Проверено 1 февраля 2016 года .
- ^ Jump up to: Перейти обратно: а б Легато М.Ю., Билезикян Дж.П. (2004). Принципы гендерной медицины . Профессиональное издательство Персидского залива. стр. 146–. ISBN 978-0-12-440906-4 . Архивировано из оригинала 14 января 2023 года . Проверено 1 февраля 2016 года .
- ^ Уильямс Д.А. (24 января 2012 г.). «Метаболизм лекарств» . В Лемке Т.Л., Уильямс Д.А. (ред.). Принципы медицинской химии Фоя . Липпинкотт Уильямс и Уилкинс. п. 164. ИСБН 978-1-60913-345-0 .
- ^ Эстрогены — достижения в исследованиях и применении: издание 2013 г.: ScholarlyBrief . Научные издания. 21 июня 2013 г. стр. 4–. ISBN 978-1-4816-7550-5 . Архивировано из оригинала 14 января 2023 года . Проверено 1 февраля 2016 года .
- ^ Стрюнкер Т., Гудвин Н., Бренкер С., Кашикар Н.Д., Вейанд И., Зайферт Р., Каупп У.Б. (март 2011 г.). «Канал CatSper опосредует индуцированный прогестероном приток Ca2+ в сперму человека». Природа . 471 (7338): 382–386. Бибкод : 2011Natur.471..382S . дои : 10.1038/nature09769 . ПМИД 21412338 . S2CID 4431334 .
- ^ Лишко П.В., Бочкина И.Л., Киричок Ю. (март 2011 г.). «Прогестерон активирует основной Ca2+-канал человеческой спермы». Природа . 471 (7338): 387–391. Бибкод : 2011Natur.471..387L . дои : 10.1038/nature09767 . ПМИД 21412339 . S2CID 4340309 .
- ^ Приор JC (2020). «Женская репродуктивная система как сбалансированное действие эстрадиола и прогестерона — революционная концепция, меняющая парадигму женского здоровья» . Открытие лекарств сегодня: модели заболеваний . 32 (Часть Б): 31–40. дои : 10.1016/j.ddmod.2020.11.005 .
- ^ Кастнер П., Краст А., Теркотт Б., Стропп У., Тора Л., Гронемейер Х., Шамбон П. (май 1990 г.). «Два различных промотора, регулируемых эстрогеном, генерируют транскрипты, кодирующие две функционально разные формы человеческого рецептора прогестерона А и В» . Журнал ЭМБО . 9 (5): 1603–1614. дои : 10.1002/j.1460-2075.1990.tb08280.x . ПМК 551856 . ПМИД 2328727 .
- ^ Jump up to: Перейти обратно: а б Клайн Дж. М., Wood CE (1 января 2006 г.). Халлам С.З., Осуч-младший (ред.). «Гормональное воздействие на молочную железу приматов в постменопаузе» . Болезнь молочной железы . 24 . IOS Press: 59–70. дои : 10.3233/bd-2006-24105 . ISBN 978-1-58603-653-9 . ПМИД 16917139 . Архивировано из оригинала 27 ноября 2023 года . Проверено 2 августа 2023 г.
- ^ Jump up to: Перейти обратно: а б Джонсон Л.Р. (2003). Основная медицинская физиология . Академическая пресса. п. 770. ИСБН 978-0-12-387584-6 . Архивировано из оригинала 14 января 2023 года . Проверено 1 февраля 2016 года .
- ^ Jump up to: Перейти обратно: а б Коад Дж., Данстолл М. (2011). Анатомия и физиология для акушерок, с онлайн-доступом Pageburst, 3: Анатомия и физиология для акушерок . Elsevier Науки о здоровье. п. 413. ИСБН 978-0-7020-3489-3 .
- ^ Ландау Р.Л., Бергенсталь Д.М., Лугибиль К., Кашт М.Е. (октябрь 1955 г.). «Метаболические эффекты прогестерона у человека». Журнал клинической эндокринологии и метаболизма . 15 (10): 1194–1215. doi : 10.1210/jcem-15-10-1194 . ПМИД 13263410 .
- ^ Jump up to: Перейти обратно: а б с Масютин М., Ядав М. (2023). «Альтернативные пути андрогенов» . Викижурнал медицины . 10 : X. doi : 10.15347/WJM/2023.003 . S2CID 257943362 .
- ^ Jump up to: Перейти обратно: а б О'Шонесси П.Дж., Антиньяк Дж.П., Ле Бизек Б., Морван М.Л., Свечников К., Сёдер О. и др. (февраль 2019 г.). «Альтернативная (закулисная) продукция андрогенов и маскулинизация у человеческого плода» . ПЛОС Биология . 17 (2): e3000002. дои : 10.1371/journal.pbio.3000002 . ПМК 6375548 . ПМИД 30763313 .
- ^ Флюк CE, Пандей А.В. (май 2014 г.). «Стероидогенез яичек - новые гены и пути». Анналы эндокринологии . 75 (2): 40–47. дои : 10.1016/j.ando.2014.03.002 . ПМИД 24793988 .
- ^ Захманн М. (февраль 1996 г.). «Призматические случаи: дефицит 17,20-десмолазы (17,20-лиазы)» . Журнал клинической эндокринологии и метаболизма . 81 (2): 457–459. дои : 10.1210/jcem.81.2.8636249 . ПМИД 8636249 .
- ^ Коррейя Дж. Н., Коннер С. Дж., Киркман-Браун Дж. К. (май 2007 г.). «Негеномное действие стероидов на сперматозоиды человека. «Постоянное щекотание от нагруженной среды» ». Семинары по репродуктивной медицине . 25 (3): 208–219. дои : 10.1055/s-2007-973433 . ПМИД 17447210 . S2CID 260318879 .
- ^ Киркман-Браун Дж.С., Брэй С., Стюарт П.М., Барратт К.Л., Publicover SJ (июнь 2000 г.). «Двухфазное повышение [Ca(2+)](i) в отдельных сперматозоидах человека, подвергшихся воздействию прогестерона» . Биология развития . 222 (2): 326–335. дои : 10.1006/dbio.2000.9729 . ПМИД 10837122 .
- ^ Киркман-Браун Дж. К., Барратт К. Л., Publicover SJ (март 2004 г.). «Медленные колебания кальция в сперматозоидах человека» . Биохимический журнал . 378 (Часть 3): 827–832. дои : 10.1042/BJ20031368 . ПМК 1223996 . ПМИД 14606954 .
- ^ Harper CV, Barratt CL, Publicover SJ (октябрь 2004 г.). «Стимуляция сперматозоидов человека градиентами прогестерона для имитации подхода к ооциту. Индукция [Ca (2+)] (i) колебаний и циклических переходов при биении жгутиков» . Журнал биологической химии . 279 (44): 46315–46325. дои : 10.1074/jbc.M401194200 . ПМИД 15322137 .
- ^ Мариб Э (2013). Анатомия и физиология . Бенджамин-Каммингс. п. 903. ИСБН 9780321887603 .
- ^ Тости Э, Ди Космо А, Куомо А, Ди Кристо С, Граньяньелло Дж (май 2001 г.). «Прогестерон вызывает активацию сперматозоидов Octopus vulgaris». Молекулярное воспроизводство и развитие . 59 (1): 97–105. дои : 10.1002/mrd.1011 . ПМИД 11335951 . S2CID 28390608 .
- ^ Jump up to: Перейти обратно: а б Боуэн Р. (6 августа 2000 г.). «Плацентарные гормоны» . Архивировано из оригинала 17 мая 2007 года . Проверено 12 марта 2008 г.
- ^ Патель Б., Эльгеро С., Такоре С., Дахуд В., Бедаиви М., Мезиано С. (2014). «Роль изоформ ядерного рецептора прогестерона в патофизиологии матки» . Обновление репродукции человека . 21 (2): 155–173. дои : 10.1093/humupd/dmu056 . ПМЦ 4366574 . ПМИД 25406186 .
- ^ Jump up to: Перейти обратно: а б Ди Ренцо Г.К., Джардина И., Клеричи Г., Брилло Э., Герли С. (июль 2016 г.). «Прогестерон при нормальной и патологической беременности». Гормональная молекулярная биология и клинические исследования . 27 (1): 35–48. doi : 10.1515/hmbci-2016-0038 . ПМИД 27662646 . S2CID 32239449 .
- ^ Care A, Невитт С.Дж., Медли Н., Донеган С., Гуд Л., Хэмпсон Л. и др. (февраль 2022 г.). «Вмешательства по предотвращению спонтанных преждевременных родов у женщин с одноплодной беременностью, находящихся в группе высокого риска: систематический обзор и сетевой метаанализ» . БМЖ . 376 : e064547. дои : 10.1136/bmj-2021-064547 . ПМЦ 8845039 . ПМИД 35168930 .
- ^ Масиас Х., Хинк Л. (2012). «Развитие молочной железы» . Междисциплинарные обзоры Wiley. Биология развития . 1 (4): 533–557. дои : 10.1002/wdev.35 . ПМЦ 3404495 . ПМИД 22844349 .
- ^ Jump up to: Перейти обратно: а б с Хилтон Х.Н., Грэм Дж.Д., Кларк С.Л. (сентябрь 2015 г.). «Мини-обзор: Регуляция прогестерона в нормальной молочной железе человека и при раке молочной железы: история двух сценариев?» . Молекулярная эндокринология . 29 (9): 1230–1242. дои : 10.1210/me.2015-1152 . ПМЦ 5414684 . ПМИД 26266959 .
- ^ Барбьери Р.Л. (13 сентября 2013 г.). «Грудь» . У Штрауса Дж. Ф., Барбьери Р. Л. (ред.). Репродуктивная эндокринология Йена и Яффе . Elsevier Науки о здоровье. стр. 236–. ISBN 978-1-4557-2758-2 . Архивировано из оригинала 14 января 2023 года . Проверено 1 февраля 2016 года .
- ^ Скейлинг А.Л., Просниц Э.Р., Хэтэуэй Х.Дж. (июнь 2014 г.). «GPER опосредует индуцированную эстрогеном передачу сигналов и пролиферацию в эпителиальных клетках молочной железы человека, а также в нормальной и злокачественной молочной железе» . Гормоны и рак . 5 (3): 146–160. дои : 10.1007/s12672-014-0174-1 . ПМК 4091989 . ПМИД 24718936 .
- ^ Jump up to: Перейти обратно: а б с д и Ауперли, доктор медицинских наук, Лейппрандт-младший, Беннетт Дж. М., Шварц Р. К., Хаслам С. З. (май 2013 г.). «Амфирегулин опосредует индуцированное прогестероном развитие протоков молочной железы в период полового созревания» . Исследование рака молочной железы . 15 (3): С44. дои : 10.1186/bcr3431 . ПМЦ 3738150 . ПМИД 23705924 .
- ^ Jump up to: Перейти обратно: а б Куль Х., Шнайдер Х.П. (август 2013 г.). «Прогестерон - промоутер или ингибитор рака молочной железы». Климактерический . 16 (Приложение 1): 54–68. дои : 10.3109/13697137.2013.768806 . ПМИД 23336704 . S2CID 20808536 .
- ^ Jump up to: Перейти обратно: а б с Траберт Б., Шерман М.Э., Каннан Н., Станчик Ф.З. (апрель 2020 г.). «Прогестерон и рак молочной железы» . Эндокринные обзоры . 41 (2): 320–344. дои : 10.1210/endrev/bnz001 . ПМК 7156851 . ПМИД 31512725 .
- ^ Совместная группа по гормональным факторам рака молочной железы (сентябрь 2019 г.). «Тип и сроки менопаузальной гормональной терапии и риск рака молочной железы: метаанализ индивидуальных участников мировых эпидемиологических данных» . Ланцет . 394 (10204): 1159–1168. дои : 10.1016/S0140-6736(19)31709-X . ПМК 6891893 . ПМИД 31474332 .
- ^ Стют П., Вильдт Л., Нойлен Дж. (апрель 2018 г.). «Влияние микронизированного прогестерона на риск рака молочной железы: систематический обзор» (PDF) . Климактерический . 21 (2): 111–122. дои : 10.1080/13697137.2017.1421925 . ПМИД 29384406 . S2CID 3642971 . Архивировано (PDF) из оригинала 2 февраля 2024 года . Проверено 2 февраля 2024 г.
- ^ Аси Н., Мохаммед К., Хайдур К., Джионфриддо М.Р., Варгас О.Л., Прокоп Л.Дж. и др. (июль 2016 г.). «Прогестерон против синтетических прогестинов и риск рака молочной железы: систематический обзор и метаанализ» . Систематические обзоры . 5 (1): 121. дои : 10.1186/s13643-016-0294-5 . ПМЦ 4960754 . ПМИД 27456847 .
- ^ Гомпель А, Плю-Бюро G (август 2018 г.). «Прогестерон, прогестины и грудь в лечении менопаузы». Климактерический . 21 (4): 326–332. дои : 10.1080/13697137.2018.1476483 . ПМИД 29852797 . S2CID 46922084 .
- ^ Дэйви Д.А. (октябрь 2018 г.). «Менопаузальная гормональная терапия: лучшее и безопасное будущее». Климактерический . 21 (5): 454–461. дои : 10.1080/13697137.2018.1439915 . ПМИД 29526116 . S2CID 3850275 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час Рейн-Феннинг, штат Нью-Джерси, член парламента от Бринката, Маскат-Барон Y (2003). «Старение кожи и менопауза: последствия для лечения». Американский журнал клинической дерматологии . 4 (6): 371–378. дои : 10.2165/00128071-200304060-00001 . ПМИД 12762829 . S2CID 20392538 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час Хольцер Г., Риглер Э., Хёнигсманн Х., Фарохния С., Шмидт Дж.Б. (сентябрь 2005 г.). «Влияние и побочные эффекты 2% крема с прогестероном на кожу женщин в пери- и постменопаузе: результаты двойного слепого, контролируемого, рандомизированного исследования». Британский журнал дерматологии . 153 (3): 626–634. дои : 10.1111/j.1365-2133.2005.06685.x . ПМИД 16120154 . S2CID 6077829 .
- ^ Король СР (9 ноября 2012 г.). Нейростероиды и нервная система . Springer Science & Business Media. стр. 44–46. ISBN 978-1-4614-5559-2 .
- ^ Флейшман Д.С., Фесслер Д.М., Чолакианс А.Е. (июль 2015 г.). «Проверка гипотезы принадлежности гомоэротической мотивации у людей: эффекты прогестерона и прайминга» . Архив сексуального поведения . 44 (5): 1395–1404. дои : 10.1007/s10508-014-0436-6 . ПМИД 25420899 . S2CID 9864224 . Архивировано из оригинала 23 сентября 2020 года . Проверено 2 августа 2023 г.
- ^ «Гомосексуальность может помочь нам сблизиться» . Новости УОП . Архивировано из оригинала 2 июля 2019 года . Проверено 2 июля 2019 г.
- ^ «Исследование предполагает, что гомосексуальные мысли являются неотъемлемой частью человеческой эволюции» . Телеграф . 25 ноября 2014 г. Архивировано из оригинала 15 февраля 2018 г. Проверено 4 апреля 2018 г.
- ^ «Новое исследование определяет эволюционную основу гомосексуализма» . ХаффПост . 26 ноября 2014 г. Архивировано из оригинала 27 ноября 2023 г. . Проверено 21 октября 2023 г.
- ^ Jump up to: Перейти обратно: а б с Ханукоглу I, Караволас Х.Дж., Гой Р.В. (апрель 1977 г.). «Метаболизм прогестерона в шишковидной железе, стволе мозга, таламусе и мозолистом теле самок крыс» . Исследования мозга . 125 (2): 313–324. дои : 10.1016/0006-8993(77)90624-2 . ПМИД 558037 . S2CID 35814845 . Архивировано из оригинала 5 марта 2021 года . Проверено 28 июня 2019 г.
- ^ Шумахер М., Геннун Р., Роберт Ф., Карелли С., Гаго Н., Гумари А. и др. (июнь 2004 г.). «Локальный синтез и двойное действие прогестерона в нервной системе: нейропротекция и миелинизация». Исследование гормона роста и IGF . 14 (Приложение А): S18–S33. дои : 10.1016/j.ghir.2004.03.007 . ПМИД 15135772 .
- ^ Сингх М., Су С., Нг С. (сентябрь 2013 г.). «Негеномные механизмы действия прогестерона в мозге» . Границы в неврологии . 7 : 159. дои : 10.3389/fnins.2013.00159 . ПМЦ 3776940 . ПМИД 24065876 .
- ^ Крыша RL, Зал ED (май 2000 г.). «Гендерные различия при острой травме ЦНС и инсульте: нейропротекторное действие эстрогена и прогестерона». Журнал нейротравмы . 17 (5): 367–388. дои : 10.1089/neu.2000.17.367 . ПМИД 10833057 .
- ^ Пан Д.С., Лю В.Г., Ян С.Ф., Цао Ф. (октябрь 2007 г.). «Тормозящее действие прогестерона на факторы воспаления после экспериментальной черепно-мозговой травмы». Биомедицинские и экологические науки . 20 (5): 432–438. ПМИД 18188998 .
- ^ Цзян С., Цзо Ф., Ван Й., Ван Дж., Ян З., Лу Х. и др. (июнь 2016 г.). «Прогестерон оказывает нейропротекторное действие и улучшает долгосрочный неврологический исход после внутримозгового кровоизлияния у мышей среднего возраста» . Нейробиология старения . 42 : 13–24. doi : 10.1016/j.neurobiolaging.2016.02.029 . ПМК 4857017 . ПМИД 27143417 .
- ^ Jump up to: Перейти обратно: а б Луома Дж.И., Стерн К.М., Мермельштейн П.Г. (август 2012 г.). «Ингибирование прогестероном передачи сигналов кальция в нейронах лежит в основе аспектов опосредованной прогестероном нейропротекции» . Журнал биохимии стероидов и молекулярной биологии . 131 (1–2): 30–36. дои : 10.1016/j.jsbmb.2011.11.002 . ПМК 3303940 . ПМИД 22101209 .
- ^ Jump up to: Перейти обратно: а б с Штейн Д.Г. (март 2008 г.). «Прогестерон оказывает нейропротекторное действие после черепно-мозговой травмы» . Обзоры исследований мозга . 57 (2): 386–397. дои : 10.1016/j.brainresrev.2007.06.012 . ПМЦ 2699575 . ПМИД 17826842 .
- ^ Эспиноза Т.Р., Райт Д.В. (2011). «Роль прогестерона при черепно-мозговой травме» . Журнал реабилитации после травм головы . 26 (6): 497–499. дои : 10.1097/HTR.0b013e31823088fa . ПМК 6025750 . ПМИД 22088981 .
- ^ Цзян С., Цзо Ф., Ван Ю, Лу Х., Ян Ц, Ван Дж. (январь 2016 г.). «Прогестерон изменяет экспрессию VEGF и BDNF и способствует нейрогенезу после ишемического инсульта» . Молекулярная нейробиология . 54 (1): 571–581. дои : 10.1007/s12035-015-9651-y . ПМЦ 4938789 . ПМИД 26746666 .
- ^ Херсон П.С., Кернер И.П., Херн П.Д. (май 2009 г.). «Секс, половые стероиды и черепно-мозговые травмы» . Семинары по репродуктивной медицине . 27 (3): 229–239. дои : 10.1055/s-0029-1216276 . ПМЦ 2675922 . ПМИД 19401954 .
- ^ Ли З, Ван Б, Кан З, Чжан Б, Ян З, Чен Дж и др. (январь 2012 г.). «Прогестерон увеличивает количество циркулирующих эндотелиальных клеток-предшественников и индуцирует регенерацию нервов после черепно-мозговой травмы у старых крыс» . Журнал нейротравмы . 29 (2): 343–353. дои : 10.1089/neu.2011.1807 . ПМЦ 3261789 . ПМИД 21534727 .
- ^ Jump up to: Перейти обратно: а б Линч В.Дж., Софуоглу М. (декабрь 2010 г.). «Роль прогестерона в никотиновой зависимости: данные от начала до рецидива» . Экспериментальная и клиническая психофармакология . 18 (6): 451–461. дои : 10.1037/a0021265 . ПМЦ 3638762 . ПМИД 21186920 .
- ^ Косгроув К.П., Эстерлис И., Макки С.А., Буа Ф., Сейбил Дж.П., Мазур СМ и др. (апрель 2012 г.). «Половые различия в наличии β2*-никотиновых рецепторов ацетилхолина у недавно воздержавшихся от курения табака» . Архив общей психиатрии . 69 (4): 418–427. doi : 10.1001/archgenpsychiatry.2011.1465 . ПМЦ 3508698 . ПМИД 22474108 .
- ^ Мелло Н.К., Кнудсон И.М., Келли М., Файвел П.А., Мендельсон Дж.Х. (октябрь 2011 г.). «Влияние прогестерона и тестостерона на самостоятельное употребление кокаина и дискриминацию кокаина самками макак-резус» . Нейропсихофармакология . 36 (11): 2187–2199. дои : 10.1038/нпп.2011.130 . ПМК 3176575 . ПМИД 21796112 .
- ^ Бузер Т (1 июня 2012 г.). «Влияние менструального цикла и гормональных контрацептивов на конкурентоспособность» (PDF) . Журнал экономического поведения и организации . Гендерные различия в неприятии риска и конкуренции. 83 (1): 1–10. дои : 10.1016/j.jebo.2011.06.006 . ISSN 0167-2681 . Архивировано (PDF) из оригинала 2 февраля 2024 года . Проверено 2 февраля 2024 г.
- ^ Шрирам Д. (2007). Медицинская химия . Нью-Дели: Dorling Kindersley India Pvt. ООО с. 432. ИСБН 978-81-317-0031-0 .
- ^ Jump up to: Перейти обратно: а б Блэкберн С (14 апреля 2014 г.). Физиология матери, плода и новорожденного . Elsevier Науки о здоровье. стр. 92–. ISBN 978-0-323-29296-2 .
- ^ Фавр Э.Дж., Ланге, Калифорния (январь 2007 г.). «Прогестероновые рецепторы активируют Wnt-1, вызывая трансактивацию рецептора эпидермального фактора роста и c-Src-зависимую устойчивую активацию митоген-активируемой протеинкиназы Erk1/2 в клетках рака молочной железы» . Молекулярная и клеточная биология . 27 (2): 466–480. дои : 10.1128/MCB.01539-06 . ПМК 1800800 . ПМИД 17074804 .
- ^ Носек ТМ. «Раздел 5/5ч9/с5ч9_13» . Основы физиологии человека . Архивировано из оригинала 24 марта 2016 года.
- ^ Ротшильд I (1969), Салханик Х.А., Кипнис Д.М., Виле Р.Л. (ред.), «Физиологическая основа эффекта прогестерона, повышающего температуру» , Метаболические эффекты гонадных гормонов и противозачаточных стероидов , Бостон, Массачусетс: Springer US, стр. 668–675, номер домена : 10.1007/978-1-4684-1782-1_49 , ISBN. 978-1-4684-1782-1 , заархивировано из оригинала 29 августа 2021 года , получено 22 марта 2021 года.
- ^ Хоулд Ф.С., Фрид ГМ, Фазекас А.Г., Трембле С., Мерсеро В.А. (декабрь 1988 г.). «Прогестероновые рецепторы регулируют моторику желчного пузыря». Журнал хирургических исследований . 45 (6): 505–512. дои : 10.1016/0022-4804(88)90137-0 . ПМИД 3184927 .
- ^ «Гормоны и здоровье полости рта» . ВебМД . Архивировано из оригинала 18 июня 2016 года . Проверено 22 июля 2013 г.
- ^ Пикард Ф., Ванатабе М., Шунджанс К., Лайдон Дж., О'Мэлли Б.В., Оверкс Дж. (ноябрь 2002 г.). «Мыши с нокаутом рецептора прогестерона имеют улучшенный гомеостаз глюкозы вследствие пролиферации бета-клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (24): 15644–15648. дои : 10.1073/pnas.202612199 . ПМЦ 137770 . ПМИД 12438645 .
- ^ Брэништяну Д.Д., Матье С. (март 2003 г.). «Прогестерон при гестационном сахарном диабете: виноват или не виновен?». Тенденции в эндокринологии и обмене веществ . 14 (2): 54–56. дои : 10.1016/S1043-2760(03)00003-1 . ПМИД 12591170 . S2CID 38209977 .
- ^ Уайнотт Р.М., Саммерс К.М., Якубиак М., Ван Вурхис Б.Дж., Мехия Р.Б. (июнь 2021 г.). «Влияние веса и индекса массы тела на уровень прогестерона в сыворотке и частоту живорождения в криоконсервированных циклах экстракорпорального оплодотворения» . Отчеты Ф&С . 2 (2): 195–200. дои : 10.1016/j.xfre.2021.02.005 . ПМЦ 8267385 . ПМИД 34278354 .
- ^ Кеммле Л.М., Штадлер А., Янка Х., фон Вольф М., Штюте П. (август 2022 г.). «Влияние микронизированного прогестерона на сердечно-сосудистые события - систематический обзор». Климактерический . 25 (4): 327–336. дои : 10.1080/13697137.2021.2022644 . ПМИД 35112635 . S2CID 246487187 .
- ^ Гассер С., Хайдемейер К., фон Вольф М., Штуте П. (июнь 2021 г.). «Влияние прогестерона на кожу и волосы в период менопаузы – всесторонний обзор». Климактерический . 24 (3): 229–235. дои : 10.1080/13697137.2020.1838476 . ПМИД 33527841 . S2CID 231757325 .
- ^ Джавед А.А., Мэйхью А.Дж., Ши А.К., Райна П. (август 2019 г.). «Связь между гормональной терапией и мышечной массой у женщин в постменопаузе: систематический обзор и метаанализ» . Открытая сеть JAMA . 2 (8): e1910154. doi : 10.1001/jamanetworkopen.2019.10154 . ПМК 6716293 . ПМИД 31461147 .
- ^ Кокоз А., Грюттер С., Стют П. (апрель 2019 г.). «Влияние микронизированного прогестерона на массу тела, индекс массы тела и метаболизм глюкозы: систематический обзор». Климактерический . 22 (2): 148–161. дои : 10.1080/13697137.2018.1514003 . ПМИД 30477366 . S2CID 53782622 .
- ^ Хэггстрем М., Ричфилд Д. (2014). «Схема путей стероидогенеза человека» . Викижурнал медицины . 1 (1). дои : 10.15347/wjm/2014.005 . ISSN 2002-4436 .
- ^ Бьюик П.М. (2002). Лекарственные натуральные продукты: биосинтетический подход . Нью-Йорк: Уайли. п. 244. ИСБН 0-471-49641-3 .
- ^ Дюпор К., Спаньоли Р., Дегрис Э., Помпон Д. (февраль 1998 г.). «Самостоятельный биосинтез прегненолона и прогестерона в модифицированных дрожжах». Природная биотехнология . 16 (2): 186–189. дои : 10.1038/nbt0298-186 . ПМИД 9487528 . S2CID 852617 .
- ^ Завод Р.М. (24 января 2012 г.). «Женское здоровье» . В Лемке Т.Л., Уильямс Д.А. (ред.). Принципы медицинской химии Фоя . Липпинкотт Уильямс и Уилкинс. стр. 1397–. ISBN 978-1-60913-345-0 . Архивировано из оригинала 14 января 2023 года . Проверено 19 июля 2018 г.
- ^ Прогестерон — Drugs.com , заархивировано из оригинала 27 марта 2019 г. , получено 23 августа 2015 г.
- ^ Jump up to: Перейти обратно: а б с Фальконе Т., Херд WW (2007). Клиническая репродуктивная медицина и хирургия . Elsevier Науки о здоровье. стр. 22–. ISBN 978-0-323-03309-1 . Архивировано из оригинала 10 января 2023 года . Проверено 6 ноября 2016 г.
- ^ Jump up to: Перейти обратно: а б с д Cupps PT (20 февраля 1991 г.). Размножение домашних животных . Эльзевир. стр. 101–. ISBN 978-0-08-057109-6 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я Станчик ФЗ (ноябрь 2003 г.). «Не все прогестины одинаковы». Стероиды . 68 (10–13): 879–890. doi : 10.1016/j.steroids.2003.08.003 . ПМИД 14667980 . S2CID 44601264 .
- ^ Дауд Ф.Дж., Джонсон Б., Мариотти А. (3 сентября 2016 г.). Фармакология и терапия в стоматологии . Elsevier Науки о здоровье. стр. 448–. ISBN 978-0-323-44595-5 .
- ^ Jump up to: Перейти обратно: а б с д Куль Х. (август 2005 г.). «Фармакология эстрогенов и прогестагенов: влияние различных путей введения». Климактерический . 8 (Приложение 1): 3–63. дои : 10.1080/13697130500148875 . ПМИД 16112947 . S2CID 24616324 .
- ^ Jump up to: Перейти обратно: а б Завод ТМ, Железник А.Я. (15 ноября 2014 г.). Физиология репродукции Кнобила и Нила . Академическая пресса. стр. 304–. ISBN 978-0-12-397769-4 .
- ^ Jump up to: Перейти обратно: а б Санторо Н.Ф., Нил-Перри Дж. (11 сентября 2010 г.). Аменорея: клиническое руководство на основе конкретных случаев . Springer Science & Business Media. стр. 13–. ISBN 978-1-60327-864-5 . Архивировано из оригинала 14 января 2023 года . Проверено 6 ноября 2016 г.
- ^ Редди Д.С. (2010). «Нейростероиды». Половые различия в человеческом мозге, их основы и последствия . Прогресс в исследованиях мозга. Том. 186. Эльзевир. стр. 113–37. дои : 10.1016/B978-0-444-53630-3.00008-7 . ISBN 9780444536303 . ПМК 3139029 . ПМИД 21094889 .
- ^ Jump up to: Перейти обратно: а б Болье Э., Келли П.А. (30 ноября 1990 г.). Гормоны: от молекул к болезни . Springer Science & Business Media. стр. 401–. ISBN 978-0-412-02791-8 .
- ^ Беранич Н., Гобек С., Рижнер Т.Л. (май 2011 г.). «Прогестины как ингибиторы 20-кетостероидредуктазы человека, AKR1C1 и AKR1C3». Химико-биологические взаимодействия . 191 (1–3): 227–233. Бибкод : 2011CBI...191..227B . дои : 10.1016/j.cbi.2010.12.012 . ПМИД 21182831 .
- ^ Андерсон Г.Д., Одегард П.С. (октябрь 2004 г.). «Фармакокинетика эстрогена и прогестерона при хронической болезни почек». Достижения в области хронической болезни почек . 11 (4): 357–360. дои : 10.1053/j.ackd.2004.07.001 . ПМИД 15492972 .
- ^ Гринблатт Дж. М., Броган К. (27 апреля 2016 г.). Интегративная терапия депрессии: новое определение моделей оценки, лечения и профилактики . ЦРК Пресс. стр. 201–. ISBN 978-1-4987-0230-0 .
- ^ Грэм С. (2 декабря 2012 г.). Репродуктивная биология человекообразных обезьян: сравнительные и биомедицинские перспективы . Эльзевир. стр. 179–. ISBN 978-0-323-14971-6 . Архивировано из оригинала 14 января 2023 года . Проверено 6 ноября 2016 г.
- ^ Струшкевич Н., Гилеп А.А., Шен Л., Эроусмит Ч.Х., Эдвардс А.М., Усанов С.А., Парк Х.В. (февраль 2013 г.). «Структурные данные о специфичности субстрата альдостеронсинтазы и целевом ингибировании» . Молекулярная эндокринология . 27 (2): 315–324. дои : 10.1210/me.2012-1287 . ПМЦ 5417327 . ПМИД 23322723 .
- ^ ван Ройен Д., Гент Р., Барнард Л., Сварт А.С. (апрель 2018 г.). «Метаболизм 11β-гидроксипрогестерона и 11-кетопрогестерона in vitro в 11-кетодигидротестостерон по черному пути». Журнал биохимии стероидов и молекулярной биологии . 178 : 203–212. дои : 10.1016/j.jsbmb.2017.12.014 . ПМИД 29277707 . S2CID 3700135 .
- ^ Jump up to: Перейти обратно: а б де Азеведо Пиччинато С (2008). Регуляция метаболизма стероидов и печеночного транскриптома эстрадиолом и прогестероном . стр. 24–25. ISBN 978-1-109-04632-8 . [ постоянная мертвая ссылка ]
- ^ Акалин С. (январь 1991 г.). «Эффекты кетоконазола у гирсутичных женщин». Акта Эндокринологика . 124 (1): 19–22. дои : 10.1530/acta.0.1240019 . ПМИД 1825737 . S2CID 9831739 .
- ^ Jump up to: Перейти обратно: а б Офрер МБ, Бенсон Х (июнь 1976 г.). «Прогестерон: обзор и последние достижения». Журнал фармацевтических наук . 65 (6): 783–800. дои : 10.1002/jps.2600650602 . ПМИД 945344 .
- ^ Стрикер Р., Эберхарт Р., Шевайеллер М.К., Куинн Ф.А., Бишоф П., Стрикер Р. (2006). «Установление подробных эталонных значений лютеинизирующего гормона, фолликулостимулирующего гормона, эстрадиола и прогестерона в разные фазы менструального цикла на анализаторе Abbott ARCHITECT». Клиническая химия и лабораторная медицина . 44 (7): 883–887. дои : 10.1515/CCLM.2006.160 . ПМИД 16776638 . S2CID 524952 .
- ^ Чапо А.И., Пулккинен М.О., Вист WG (март 1973 г.). «Эффекты лютеэктомии и заместительной терапии прогестероном у беременных на ранних сроках». Американский журнал акушерства и гинекологии . 115 (6): 759–765. дои : 10.1016/0002-9378(73)90517-6 . ПМИД 4688578 .
- ^ Клинический центр НИЗ (16 августа 2004 г.). «Исторические эталонные диапазоны прогестерона» . Национальные институты здравоохранения США. Архивировано из оригинала 9 января 2009 года . Проверено 12 марта 2008 г.
- ^ Чернецкий CC, Бергер Б.Дж. (31 октября 2012 г.). Лабораторные исследования и диагностические процедуры - Электронная книга . Elsevier Науки о здоровье. стр. 908–. ISBN 978-1-4557-4502-9 . Архивировано из оригинала 27 февраля 2024 года . Проверено 23 августа 2023 г.
- ^ Беккер К.Л. (2001). Принципы и практика эндокринологии и обмена веществ . Липпинкотт Уильямс и Уилкинс. стр. 889, 940. ISBN. 978-0-7817-1750-2 . Архивировано из оригинала 27 февраля 2024 года . Проверено 23 августа 2023 г.
- ^ Йосимович Ж.Б. (11 ноября 2013 г.). Гинекологическая эндокринология . Springer Science & Business Media. стр. 9, 25–29, 139. ISBN. 978-1-4613-2157-6 . Архивировано из оригинала 14 января 2023 года . Проверено 1 февраля 2016 года .
- ^ ван Кип П., Утян В. (6 декабря 2012 г.). Предменструальный синдром: материалы семинара, проведенного во время Шестого Международного конгресса психосоматической акушерства и гинекологии, Берлин, сентябрь 1980 г. Springer Science & Business Media. стр. 51–52. ISBN 978-94-011-6255-5 . Архивировано из оригинала 14 января 2023 года . Проверено 1 февраля 2016 года .
- ^ Штраус Дж. Ф., Барбьери Р.Л. (2009). Репродуктивная эндокринология Йена и Яффе: физиология, патофизиология и клиническое ведение . Elsevier Науки о здоровье. стр. 807–. ISBN 978-1-4160-4907-4 . Архивировано из оригинала 10 января 2023 года . Проверено 23 августа 2023 г.
- ^ Баджадж Л., Берман С. (1 января 2011 г.). Принятие педиатрических решений по Берману . Elsevier Науки о здоровье. стр. 160–. ISBN 978-0-323-05405-8 . Архивировано из оригинала 11 января 2023 года . Проверено 23 августа 2023 г.
- ^ Лауритцен С (1988). «Природные и синтетические половые гормоны – биологические основы и принципы медицинского лечения». У Германа П.Г. Шнайдера, Кристиана Лауритцена, Эберхарда Нишлага (ред.). репродукции человека ( Фонды и клиника на немецком языке). Вальтер де Грюйтер. стр. 229–306. ISBN 978-3110109689 . OCLC 35483492 . Архивировано из оригинала 1 октября 2023 года . Проверено 23 августа 2023 г.
- ^ Литтл, AB, и Биллиар, РБ (1983). Прогестагены. В эндокринологии беременности, 3-е издание (стр. 92–111). Харпер и Роу Филадельфия. https://scholar.google.com/scholar?cluster=2512291948467467634 Архивировано 22 февраля 2022 г. в Wayback Machine.
- ^ Jump up to: Перейти обратно: а б с д и ж г час я Референтные диапазоны прогестерона , данные проведены в Клиническом центре Национального института здравоохранения, Бетесда, Мэриленд, 3 февраля 2009 г.
- ^ Jump up to: Перейти обратно: а б с д и ж г час Преобразовано из значений массы с использованием молярной массы 314,46 г/моль.
- ^ Хэггстрем М (2014). «Референтные диапазоны эстрадиола, прогестерона, лютеинизирующего гормона и фолликулостимулирующего гормона во время менструального цикла» . Викижурнал медицины . 1 (1). дои : 10.15347/wjm/2014.001 . ISSN 2002-4436 . S2CID 88035135 .
- ^ Гудсон III WH, Хандагама П., Мур II Д.Х., Дэйрки С. (13 декабря 2007 г.). «Молочные продукты являются источником пищевого прогестерона» . 30-й ежегодный симпозиум по раку молочной железы в Сан-Антонио. стр. Аннотация № 2028. Архивировано из оригинала 20 сентября 2008 года . Проверено 12 марта 2008 г.
- ^ Паули Г.Ф., Фризен Дж.Б., Гёдеке Т., Фарнсворт Н.Р., Глодный Б. (март 2010 г.). «Появление прогестерона и родственных ему животных стероидов у двух высших растений». Журнал натуральных продуктов . 73 (3): 338–345. дои : 10.1021/np9007415 . ПМИД 20108949 . S2CID 26467578 .
- ^ Эпплцвейг Н. (май 1969 г.). "Стероиды". Химическая неделя . 104 : 57–72. ПМИД 12255132 .
- ^ Ногучи Э., Фудзивара Ю., Мацусита С., Икеда Т., Оно М., Нохара Т. (сентябрь 2006 г.). «Метаболизм томатных стероидных гликозидов в организме человека» . Химический и фармацевтический вестник . 54 (9): 1312–1314. дои : 10.1248/cpb.54.1312 . ПМИД 16946542 .
- ^ Ян DJ, Лу TJ, Хван Л.С. (октябрь 2003 г.). «Выделение и идентификация стероидных сапонинов в тайваньском сорте ямса (Dioscorea pseudojaponica Yamamoto)» (PDF) . Журнал сельскохозяйственной и пищевой химии . 51 (22): 6438–6444. дои : 10.1021/jf030390j . ПМИД 14558759 . Архивировано (PDF) из оригинала 3 августа 2022 года . Проверено 2 апреля 2022 г.
- ^ «Окончательный отчет об измененной оценке безопасности экстракта корня Dioscorea villosa (дикий ямс)» . Международный журнал токсикологии . 23 (Приложение 2): 49–54. 2004. дои : 10.1080/10915810490499055 . ПМИД 15513824 . S2CID 962216 .
- ^ Ниньо Дж., Хименес Д.А., Москера О.М., Корреа Ю.М. (2007). «Количественное определение диосгенина с помощью ВЭЖХ в коллекции клубней Dioscorea Polygonoides из колумбийской флоры» . Журнал Бразильского химического общества . 18 (5): 1073–1076. дои : 10.1590/S0103-50532007000500030 . S2CID 95193700 .
- ^ Мёда Т., Нагай Т., Нагасима Т. (2005). «Свойства крахмалов в клубнях ямса (Dioscorea spp.)». Текущие темы пищевой науки и технологий : 105–114. ISBN 81-308-0003-9 .
- ^ Весп Л.М., Deutsch MB (март 2017 г.). «Варианты гормонального и хирургического лечения трансгендерных женщин и лиц трансженского спектра». Психиатрические клиники Северной Америки . 40 (1): 99–111. дои : 10.1016/j.psc.2016.10.006 . ПМИД 28159148 .
- ^ Долицкий С.Н., Кордейро Митчелл CN, Штадлер СС, Сегарс Дж.Х. (ноябрь 2020 г.). «Эффективность лечения только прогестином для лечения симптомов менопаузы: систематический обзор». Менопауза . 28 (2): 217–224. дои : 10.1097/GME.0000000000001676 . ПМИД 33109992 . S2CID 225100434 .
- ^ Руан X, Муек АО (ноябрь 2014 г.). «Системная терапия прогестероном — пероральная, вагинальная, инъекционная и даже трансдермальная?». Матуритас . 79 (3): 248–255. дои : 10.1016/j.maturitas.2014.07.009 . ПМИД 25113944 .
- ^ Филикори М (ноябрь 2015 г.). «Клиническая роль и применение прогестерона в репродуктивной медицине: обзор» . Acta Obstetricia et Gynecologica Scandinavica . 94 (Приложение 161): 3–7. дои : 10.1111/aogs.12791 . ПМИД 26443945 .
- ^ Чампалья В., Когниньи GE (ноябрь 2015 г.). «Клиническое применение прогестерона при бесплодии и вспомогательных репродуктивных технологиях» . Acta Obstetricia et Gynecologica Scandinavica . 94 (Приложение 161): 17–27. дои : 10.1111/aogs.12770 . ПМИД 26345161 . S2CID 40753277 .
- ^ Чой С.Дж. (сентябрь 2017 г.). «Использование дополнительной терапии прогестероном для предотвращения преждевременных родов: обзор литературы» . Наука акушерства и гинекологии . 60 (5): 405–420. дои : 10.5468/ogs.2017.60.5.405 . ПМК 5621069 . ПМИД 28989916 .
- ^ Кумарасами А., Харб Х.М., Девалл А.Дж., Чид В., Робертс Т.Е., Горанитис I и др. (июнь 2020 г.). «Прогестерон для предотвращения выкидыша у женщин с кровотечением на ранних сроках беременности: РКИ PRISM» . Оценка технологий здравоохранения . 24 (33): 1–70. дои : 10.3310/hta24330 . ПМК 7355406 . ПМИД 32609084 .
- ^ Jump up to: Перейти обратно: а б Элкс Дж. (14 ноября 2014 г.). Словарь лекарств: Химические данные: Химические данные, структуры и библиография . Спрингер. стр. 1024–. ISBN 978-1-4757-2085-3 .
- ^ Jump up to: Перейти обратно: а б Index Nominum 2000: Международный каталог лекарств . Тейлор и Фрэнсис. Январь 2000 г., стр. 880–. ISBN 978-3-88763-075-1 .
- ^ Нумадзава М., Нагаока М., Кунитама Ю. (сентябрь 1986 г.). «Региоспецифическая дезоксигенация дигидроксиацетонового фрагмента у C-17 кортикостероидов йодтриметилсиланом» . Химический и фармацевтический вестник . 34 (9): 3722–3726. дои : 10.1248/cpb.34.3722 . ПМИД 3815593 .
- ^ Jump up to: Перейти обратно: а б Маркер Р.Э., Крюгер Дж. (1940). «Стерины. CXII. Сапогенины. XLI. Получение триллина и его превращение в прогестерон». Дж. Ам. хим. Соц . 62 (12): 3349–3350. дои : 10.1021/ja01869a023 .
- ^ Госвами А., Котоки Р., Растоги Р.К., Гош А.К. (1 мая 2003 г.). «Эффективный процесс получения 16-дегидропрегненолона ацетата в одном котле». Исследования и разработки органических процессов . 7 (3): 306–308. дои : 10.1021/op0200625 .
- ^ Хейл Ф.В. (1950). «Прогестерон из 3-ацетоксибиснор-5-холенальдегида и 3-кетобиснор-4-холенальдегида». Журнал Американского химического общества . 72 (6): 2617–2619. дои : 10.1021/ja01162a076 .
- ^ Сломп Г (1958). «Озонолиз. II. 1 Влияние пиридина на озонолиз 4,22-стигмастадиен-3-она 2». Журнал Американского химического общества . 80 (4): 915–921. дои : 10.1021/ja01537a041 .
- ^ Сундарараман П., Джерасси К. (октябрь 1977 г.). «Удобный синтез прогестерона из стигмастерола». Журнал органической химии . 42 (22): 3633–3634. дои : 10.1021/jo00442a044 . ПМИД 915584 .
- ^ «Стенограммы Новы: забытый гений» . PBS.org. 6 февраля 2007 г. Архивировано из оригинала 11 октября 2018 г. . Проверено 8 сентября 2017 г.
- ^ «Гиганты прошлого» . Lipidlibrary.aocs.org. Архивировано из оригинала 15 апреля 2012 года.
- ^ Jump up to: Перейти обратно: а б с Джонсон WS, Грейвесток MB, Маккерри BE (август 1971 г.). «Участие ацетиленовой связи в биогенетически подобных циклизациях олефинов. II. Синтез dl-прогестерона». Журнал Американского химического общества . 93 (17): 4332–4334. дои : 10.1021/ja00746a062 . ПМИД 5131151 .
- ^ Угловой GW, Аллен В.М. (1 марта 1929 г.). «Физиология желтого тела» . Американский журнал физиологии. Устаревший контент . 88 (2): 326–339. дои : 10.1152/ajplegacy.1929.88.2.326 . ISSN 0002-9513 . Архивировано из оригинала 12 августа 2021 года . Проверено 12 августа 2021 г.
- ^ Jump up to: Перейти обратно: а б с Коутиньо Э.М., Сигал С.Дж. (1999). Менструация устарела? . Издательство Оксфордского университета. стр. 31–. ISBN 978-0-19-513021-8 . Архивировано из оригинала 14 января 2023 года . Проверено 5 октября 2016 г.
- ^ Уокер А. (7 марта 2008 г.). Менструальный цикл . Рутледж. стр. 49–. ISBN 978-1-134-71411-7 .
- ^ Пиосик Р. (2003). «Адольф Бутенандт и его работа в Гданьском политехническом университете». ЧЕМКОН . 10 (3): 135–138. дои : 10.1002/ckon.200390038 .
- ^ Гинзбург Б. (6 декабря 2012 г.). Предменструальный синдром: этические и юридические последствия с биомедицинской точки зрения . Springer Science & Business Media. стр. 274–. ISBN 978-1-4684-5275-4 . Архивировано из оригинала 14 января 2023 года . Проверено 5 октября 2016 г.
- ^ Роллстон HD (1936). Эндокринные органы в здоровье и болезни: исторический обзор . Издательство Оксфордского университета, Х. Милфорд. п. 406. Архивировано из оригинала 14 января 2023 года . Проверено 5 октября 2016 г.
- ^ Аллен В.М. (октябрь 1970 г.). «Прогестерон: как появилось название?». Южный медицинский журнал . 63 (10): 1151–1155. дои : 10.1097/00007611-197010000-00012 . ПМИД 4922128 . S2CID 35867375 .
- ^ Рефсал К. (февраль 2009 г.). «Интерпретация результатов сывороточного прогестерона для управления разведением собак» (PDF) . Webcd.endo.ref . Архивировано из оригинала 29 августа 2021 года . Проверено 26 февраля 2018 г.
- ^ «Цены на прогестерон, купоны и советы по экономии — GoodRx» . www.goodrx.com . Архивировано из оригинала 30 марта 2023 года . Проверено 1 августа 2023 г.
- ^ «Цены на прогестерон, купоны и советы по экономии — GoodRx» . www.goodrx.com . Архивировано из оригинала 30 марта 2023 года . Проверено 1 августа 2023 г.
Внешние ссылки [ править ]
- Прогестерон МС Спектр
- Прогестерон Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
- Кимбалл Дж.В. (27 мая 2007 г.). «Прогестерон» . Страницы биологии Кимбалла . Архивировано из оригинала 18 июня 2008 года . Проверено 18 июня 2008 г.
- Прогестерон
- Ингибиторы 5α-редуктазы
- Ингибиторы 11β-гидроксистероиддегидрогеназы
- Производные алкена
- Антиминералокортикоиды
- Дикетоны
- Положительные аллостерические модуляторы ГАМКА-рецепторов
- Глюкокортикоиды
- Антагонисты глициновых рецепторов
- Нейростероиды
- Гепатотоксины
- Гормоны гипоталамус-гипофиз-надпочечниковой системы.
- Гормоны оси гипоталамус-гипофиз-гонады
- Гормоны гипоталамо-гипофизарно-пролактиновой оси
- Гормоны яичников
- Гормоны плаценты
- Гормоны надпочечниковой коры
- Гормоны головного мозга
- Гормоны беременной женщины
- Женская эндокринная система человека
- Человеческие гормоны
- Агонисты рецепторов прегнана X
- Преньянес
- Прогестагены
- Релизеры пролактина
- Половые гормоны
- Антагонисты сигмы
- Стероидные гормоны
- Полный синтез