Микроглия
| Микроглия | |
|---|---|
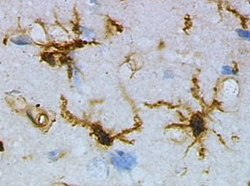 Микроглия в состоянии покоя из коры головного мозга крысы перед черепно-мозговой травмой (окрашивание лектина HRP ) | |
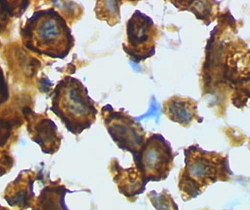 Микроглия/макрофаг – активированная форма коры головного мозга крысы после черепно-мозговой травмы (окрашивание лектина HRP) | |
| Подробности | |
| Предшественник | Примитивные макрофаги, полученные из желточного мешка |
| Система | Центральная нервная система |
| Идентификаторы | |
| МеШ | Д017628 |
| ТД | Х2.00.06.2.00004, Х2.00.06.2.01025 |
| ФМА | 54539 |
| Анатомические термины микроанатомии | |
Микроглия — это тип нейроглии (глиальных клеток), расположенных по всему головному и спинному мозгу . [1] Микроглия составляет около 10-15% клеток головного мозга. [2] Как резидентные макрофаги , они действуют как первая и основная форма активной иммунной защиты в центральной нервной системе (ЦНС). [3] Микроглия возникает в желточном мешке в результате строго регулируемого молекулярного процесса. [4] Эти клетки (и другие нейроглии, включая астроциты ) распределены в больших непересекающихся областях по всей ЦНС. [5] [6] Микроглия является ключевыми клетками в общем поддержании мозга: они постоянно очищают ЦНС от бляшек , поврежденных или ненужных нейронов и синапсов , а также инфекционных агентов. [7] Поскольку эти процессы должны быть эффективными для предотвращения потенциально фатальных повреждений, микроглия чрезвычайно чувствительна даже к небольшим патологическим изменениям в ЦНС. [8] Эта чувствительность частично достигается наличием уникальных калиевых каналов , которые реагируют даже на небольшие изменения внеклеточного калия. [7] Недавние данные показывают, что микроглия также играет ключевую роль в поддержании нормальных функций мозга в здоровых условиях. [9] Микроглия также постоянно контролирует функции нейронов посредством прямых соматических контактов и при необходимости оказывает нейропротекторное действие. [10]
Головной и спинной мозг, составляющие ЦНС, обычно не подвергаются непосредственному воздействию патогенных факторов кровообращения организма из-за ряда эндотелиальных клеток, известных как гематоэнцефалический барьер или ГЭБ. ГЭБ предотвращает попадание большинства инфекций в уязвимую нервную ткань. В случае, когда инфекционные агенты попадают непосредственно в мозг или пересекают гематоэнцефалический барьер, клетки микроглии должны реагировать быстро, чтобы уменьшить воспаление и уничтожить инфекционные агенты до того, как они повредят чувствительную нервную ткань. Из-за нехватки антител из остального организма (немногие антитела достаточно малы, чтобы преодолеть гематоэнцефалический барьер), микроглия должна быть способна распознавать инородные тела, глотать их и действовать как антигенпрезентирующие клетки, активирующие Т-клетки. .
История [ править ]
Возможность просматривать и характеризовать различные нервные клетки, включая микроглию, появилась в 1880 году, когда Франц Ниссль разработал окраску по Нисслю . Франц Ниссль и Уильям Форд Робертсон впервые описали клетки микроглии во время своих гистологических экспериментов. Методы окрашивания клеток в 1880-х годах показали, что микроглия связана с макрофагами . Активация микроглии и образование разветвленных кластеров микроглии впервые были отмечены Виктором Бабешем во время изучения случая бешенства в 1897 году. Бабеш отметил, что клетки были обнаружены при различных вирусных инфекциях головного мозга, но не знал, что это за кластеры микроглии, которые он видел. [11] Испанский ученый Сантьяго Рамон-и-Кахаль определил «третий элемент» (тип клеток), помимо нейронов и астроцитов. [12] Пио дель Рио Ортега , ученик Сантьяго Рамон-и-Кахаля , впервые назвал клетки «микроглией» примерно в 1920 году. Далее он охарактеризовал реакцию микроглии на поражения головного мозга в 1927 году и отметил «фонтаны микроглии», присутствующие в мозолистом теле и других перинатальные области белого вещества в 1932 году. После многих лет исследований Рио Хортегу стали считать «отцом микроглии». [13] [14] В течение длительного периода времени наши знания о микроглии практически не улучшились. Затем, в 1988 году, Хики и Кимура показали, что периваскулярные микроглиальные клетки происходят из костного мозга и экспрессируют высокие уровни белков MHC класса II, используемых для презентации антигена. Это подтвердило постулат Пио Дель Рио-Хортеги о том, что клетки микроглии функционируют аналогично макрофагам , осуществляя фагоцитоз и презентацию антигена . [ нужна ссылка ]
Формы [ править ]

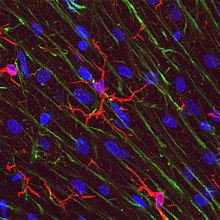
Клетки микроглии чрезвычайно пластичны и претерпевают множество структурных изменений в зависимости от местоположения и системных потребностей. Этот уровень пластичности необходим для выполнения огромного разнообразия функций, которые выполняет микроглия. Способность к трансформации отличает микроглию от макрофагов , которые необходимо регулярно заменять, и дает им возможность в кратчайшие сроки защищать ЦНС, не вызывая иммунологических нарушений. [7] Микроглия принимает специфическую форму или фенотип в ответ на местные условия и химические сигналы, которые они обнаруживают. [15]
Разветвленный [ править ]
Эта форма микроглиальных клеток обычно встречается в определенных местах по всему головному и спинному мозгу при отсутствии инородного материала или умирающих клеток. Эта «покоящаяся» форма микроглии состоит из длинных ветвящихся отростков и небольшого клеточного тела. В отличие от амебоидных форм микроглии, тело клетки разветвленной формы остается на месте, в то время как ее ветви постоянно перемещаются и обозревают окружающую территорию. Ветви очень чувствительны к небольшим изменениям физиологического состояния и требуют очень специфических условий культивирования для наблюдения in vitro . [15]
В отличие от активированной или амебоидной микроглии, разветвленная микроглия не фагоцитирует клетки и секретирует меньше иммуномолекул (в том числе белков MHC класса I / II ). Микроглия в этом состоянии способна искать и идентифицировать иммунные угрозы, сохраняя при этом гомеостаз в ЦНС. [16] [17] [18] Хотя это считается состоянием покоя, микроглия в этой форме по-прежнему чрезвычайно активна в химическом исследовании окружающей среды. Разветвленная микроглия может трансформироваться в активированную форму в любой момент в ответ на травму или угрозу. [15]
Реактивный (Активированный) [ править ]
Хотя исторически термин «активированная» микроглия используется часто, его следует заменить на «реактивную» микроглию. [19] Действительно, по-видимому, покоящаяся микроглия не лишена активных функций, и термин «активация» вводит в заблуждение, поскольку он имеет тенденцию указывать на поляризацию клеточной реактивности «все или ничего». Маркер Iba1 , уровень экспрессии которого повышен в реактивной микроглии, часто используется для визуализации этих клеток. [20]
Нефагоцитарный [ править ]
Это состояние на самом деле является частью ступенчатой реакции, когда микроглия переходит от разветвленной формы к полностью активной фагоцитарной форме. Микроглия может быть активирована различными факторами, включая: провоспалительные цитокины , факторы некроза клеток , липополисахарид и изменения внеклеточного калия (признаки разрыва клеток). После активации клетки претерпевают несколько ключевых морфологических изменений, включая утолщение и ретракцию ветвей, поглощение белков MHC класса I/II, экспрессию иммуномолекул, секрецию цитотоксических факторов, секрецию рекрутирующих молекул и секрецию провоспалительных сигнальных молекул (в результате в каскаде сигналов, способствующих воспалению). Активированная нефагоцитарная микроглия обычно проявляется в виде «кустистых», «палочек» или небольших амебоидов в зависимости от того, насколько далеко они находятся в континууме разветвленной или полной фагоцитарной трансформации. Кроме того, микроглия также подвергается быстрой пролиферации с целью увеличения их количества. Со строго морфологической точки зрения, изменение формы микроглии вдоль континуума связано с изменением морфологической сложности и может быть оценено количественно с помощью методов фрактального анализа, которые оказались чувствительными даже к тонким, визуально необнаружимым изменениям, связанным с различной морфологией в разных патологических состояниях. . [7] [16] [17] [21]
Фагоцитарный [ править ]
Активированная фагоцитарная микроглия является максимально иммунореактивной формой микроглии. Эти клетки обычно принимают большую амебоидную форму, хотя наблюдаются некоторые различия. Помимо наличия антигенпрезентирующих, цитотоксических и опосредующих воспаление сигналов активированной нефагоцитарной микроглии, они также способны фагоцитировать чужеродные материалы и отображать полученные иммуномолекулы для активации Т-клеток . Фагоцитарные микроглии перемещаются к месту повреждения, поглощают раздражающий материал и выделяют провоспалительные факторы, чтобы стимулировать пролиферацию большего количества клеток и делать то же самое. Активированная фагоцитарная микроглия также взаимодействует с астроцитами и нервными клетками, чтобы как можно быстрее бороться с любой инфекцией или воспалением с минимальным повреждением здоровых клеток мозга. [7] [16]
Амебоид [ править ]
Такая форма позволяет микроглии свободно перемещаться по нервной ткани, что позволяет ей выполнять свою роль клетки-мусорщика. Амебоидная микроглия способна фагоцитировать остатки, но не выполняет ту же антигенпрезентирующую и воспалительную роль, что и активированная микроглия . Амебоидная микроглия особенно распространена во время развития и перестройки мозга, когда необходимо большое количество внеклеточного мусора и апоптотических клеток удалить . Эта форма микроглиальных клеток встречается в основном в перинатальных белого вещества областях мозолистого тела, известных как «Фонтаны микроглии». [7] [17] [22]
Гиттерные ячейки [ править ]
Клетки Гиттера являются конечным результатом фагоцитоза микроглиальных клеток инфекционного материала или клеточного мусора. В конце концов, после поглощения определенного количества материала, фагоцитирующая микроглиальная клетка становится неспособной фагоцитировать какие-либо дальнейшие материалы. Полученная клеточная масса известна как зернистое тельце, названное так из-за его «зернистого» вида. Глядя на ткань, окрашенную для обнаружения гиттерных клеток, патологоанатомы могут визуализировать зажившие участки после инфекции. [23]
Периваскулярный [ править ]
В отличие от других типов микроглии, упомянутых выше, «периваскулярная» микроглия относится к местоположению клетки, а не к ее форме/функции. Периваскулярная микроглия преимущественно локализована в стенках базальной пластинки . заменяются клетками-предшественниками, происходящими из костного мозга Они выполняют нормальные функции микроглии, но, в отличие от нормальной микроглии, регулярно , и экспрессируют антигены MHC класса II независимо от окружающей среды. Периваскулярная микроглия также сильно реагирует на антигены дифференцировки макрофагов. [7] Было показано, что эта микроглия необходима для восстановления сосудистых стенок, как показали эксперименты Риттера и наблюдения за ишемической ретинопатией . Периваскулярная микроглия способствует пролиферации эндотелиальных клеток , позволяя формироваться новым сосудам и восстанавливать поврежденные сосуды. Во время этих процессов миелоидных клеток в клетки микроглии. для выполнения этих задач сильно ускоряется рекрутирование и дифференцировка [24]
Юкставаскулярный [ править ]
Как и периваскулярную микроглию, околососудистую микроглию можно отличить главным образом по расположению. Юкставаскулярная микроглия обнаруживается в непосредственном контакте со стенкой базальной пластинки кровеносных сосудов, но не обнаруживается внутри стенок. В этом положении они могут взаимодействовать как с эндотелиальными клетками, так и с перицитами . [25] Подобно периваскулярным клеткам, они экспрессируют белки MHC класса II даже при низких уровнях активности воспалительных цитокинов . В отличие от периваскулярных клеток, но подобно резидентной микроглии, юкставаскулярная микроглия не демонстрирует быстрого обновления или замены миелоидными клетками-предшественниками на регулярной основе. [7]
Функции [ править ]

Клетки микроглии выполняют множество различных задач в ЦНС, в основном связанных как с иммунным ответом, так и с поддержанием гомеостаза. Ниже приведены некоторые из основных известных функций, выполняемых этими клетками. [ нужна ссылка ]
Уборка [ править ]
Помимо того, что каждая микроглиальная клетка очень чувствительна к небольшим изменениям в окружающей среде, она также регулярно физически обследует свою область. Это действие осуществляется в состояниях амебоида и покоя. Если при движении через установленную область микроглиальная клетка обнаруживает какой-либо инородный материал, поврежденные клетки, апоптотические клетки, нейрофибриллярные клубки , фрагменты ДНК или бляшки , она активирует и фагоцитирует материал или клетку. Таким образом, микроглиальные клетки также действуют как «домохозяйки», убирая случайный клеточный мусор. [16] Во время онтогенетического связывания мозга микроглиальные клетки играют большую роль, регулируя количество нейральных клеток-предшественников и удаляя апоптотические нейроны. Есть также доказательства того, что микроглия может совершенствовать синаптические схемы, поглощая и устраняя синапсы. [26] После развития большинство мертвых или апоптотических клеток обнаруживается в коре головного мозга и подкорковом белом веществе . Это может объяснить, почему большинство амебоидных клеток микроглии обнаруживается в «фонтанах микроглии» в коре головного мозга. [22]
Фагоцитоз [ править ]
Основная роль микроглии — фагоцитоз — заключается в поглощении различных материалов. Поглощенные материалы обычно состоят из клеточного мусора, липидов и апоптотических клеток в невоспаленном состоянии, а также вторгшихся вирусов , бактерий или других инородных материалов в воспаленном состоянии. Как только микроглиальная клетка «наполняется», она прекращает фагоцитарную активность и превращается в относительно нереактивную гиттерную клетку . [27]
Внеклеточная сигналов передача
Большая часть роли микроглиальных клеток в головном мозге заключается в поддержании гомеостаза в неинфицированных областях и стимулировании воспаления в инфицированных или поврежденных тканях. Микроглия достигает этого посредством чрезвычайно сложной серии внеклеточных сигнальных молекул, которые позволяют ей взаимодействовать с другой микроглией, астроцитами , нервами , Т-клетками и миелоидными клетками-предшественниками . Как упоминалось выше, цитокин IFN-γ можно использовать для активации клеток микроглии. Кроме того, после активации IFN-γ микроглия также высвобождает больше IFN-γ во внеклеточное пространство. Это активирует больше микроглии и запускает каскад активации, индуцированный цитокинами, быстро активируя всю близлежащую микроглию. Продуцируемый микроглией TNF-α вызывает апоптоз нервной ткани и усиливает воспаление. IL-8 способствует росту и дифференцировке B-клеток , что позволяет ему помогать микроглии в борьбе с инфекцией. Другой цитокин, IL-1 , ингибирует цитокины IL-10 и TGF-β , которые подавляют презентацию антигена и передачу провоспалительных сигналов. Дополнительный дендритные клетки и Т-клетки рекрутируются в место повреждения посредством микроглиальной продукции хемотаксических молекул, таких как MDC , IL-8 и MIP-3β . Наконец, PGE 2 и другие простаноиды предотвращают хроническое воспаление , ингибируя провоспалительную реакцию микроглии и подавляя реакцию Th1 (Т-хелперных клеток). [16]
Презентация антигена
Как упоминалось выше, резидентная неактивированная микроглия действует как плохая антигенпрезентирующая клетка из-за отсутствия у нее белков MHC класса I/II. После активации они быстро экспрессируют белки MHC класса I/II и быстро становятся эффективными презентаторами антигена. В некоторых случаях микроглия также может быть активирована IFN-γ для представления антигенов , но не функционирует так эффективно, как если бы она подверглась поглощению белков MHC класса I/II. Во время воспаления благодаря специализированным поверхностным маркерам , Т-клетки преодолевают гематоэнцефалический барьер а затем напрямую связываются с микроглией, чтобы получить антигены . После того, как им были представлены антигены, Т-клетки продолжают выполнять различные функции, включая провоспалительную рекрутацию, формирование иммунопамяти, секрецию цитотоксических материалов и прямые атаки на плазматические мембраны чужеродных клеток. [7] [16]
Цитотоксичность [ править ]
Помимо способности уничтожать инфекционные организмы посредством межклеточного контакта посредством фагоцитоза , микроглия также может выделять различные цитотоксические вещества. [28] Микроглия в культуре секретирует большое количество перекиси водорода и оксида азота в процессе, известном как « респираторный взрыв ». Оба этих химических вещества могут напрямую повредить клетки и привести к гибели нейронов. Протеазы, секретируемые микроглией, катаболизируют специфические белки, вызывая прямое повреждение клеток, тогда как цитокины, такие как IL-1, способствуют демиелинизации аксонов нейронов. Наконец, микроглия может повреждать нейроны посредством процессов, опосредованных рецептором NMDA, секретируя глутамат , аспартат и хинолиновую кислоту . Цитотоксическая секреция направлена на уничтожение инфицированных нейронов, вирусов и бактерий, но также может вызвать большое количество побочных нейронных повреждений. В результате хроническая воспалительная реакция может привести к крупномасштабному повреждению нервов, поскольку микроглия разрушает мозг, пытаясь уничтожить вторгшуюся инфекцию. [7] Эдаравон , поглотитель радикалов, предотвращает окислительную нейротоксичность, провоцируемую активированной микроглией. [29]
Синаптическое зачистка [ править ]
Явление, впервые замеченное при поражениях позвоночника Блинзингером и Крейцбергом в 1968 году, заключается в том, что послевоспалительная микроглия удаляет ветви нервов вблизи поврежденных тканей. Это способствует возобновлению роста и переназначению поврежденных нейронных цепей . [7] Также было показано, что микроглия участвует в процессе обрезки синапсов во время развития мозга. [30]
Продвижение ремонта [ править ]
После воспаления микроглия проходит несколько этапов, способствующих возобновлению роста нервной ткани. К ним относятся зачистка синапсов, секреция противовоспалительных цитокинов , рекрутирование нейронов и астроцитов в поврежденную область и образование желтых клеток . Без роста и переназначения микроглиальных клеток в резидентных областях ЦНС они были бы значительно медленнее и практически невозможны во многих сосудистых системах, окружающих мозг и глаза. [7] [24] Недавние исследования подтвердили, что процессы микроглии постоянно контролируют функции нейронов через специализированные соматические соединения и определяют «благополучие» нервных клеток. Посредством этого пути межклеточной коммуникации микроглия способна оказывать сильное нейропротекторное действие, внося значительный вклад в восстановление после травмы головного мозга. [10] Также было показано, что микроглия способствует правильному развитию мозга за счет контакта с незрелыми, развивающимися нейронами. [31]
Развитие [ править ]

Долгое время считалось, что клетки микроглии дифференцируются в костном мозге из гемопоэтических стволовых клеток — предшественников всех клеток крови. Однако недавние исследования показывают, что микроглия зарождается в желточном мешке в течение очень ограниченного эмбрионального периода и заселяет паренхиму мозга, руководствуясь точно организованным молекулярным процессом. [4] Клетки-предшественники желточного мешка требуют активации рецептора колониестимулирующего фактора 1 (CSF1R) для миграции в мозг и дифференцировки в микроглию. [32] Кроме того, наибольший вклад в репопуляцию микроглии основан на ее локальном самообновлении, как в устойчивом состоянии, так и при заболевании, тогда как циркулирующие моноциты также могут вносить вклад в меньшей степени, особенно при заболевании. [4] [33]
Моноциты также могут дифференцироваться в миелоидные дендритные клетки и макрофаги периферических систем. Как и макрофаги в остальном организме, микроглия использует фагоцитарные и цитотоксические механизмы для уничтожения чужеродных материалов. Микроглия и макрофаги способствуют иммунному ответу, действуя как антигенпредставляющие клетки , а также стимулируя воспаление и гомеостатические механизмы внутри организма, секретируя цитокины и другие сигнальные молекулы. [34]
В своей форме с пониженной регуляцией в микроглии отсутствуют белки MHC класса I / MHC класса II , цитокины IFN-γ , антигены CD45 и многие другие поверхностные рецепторы, необходимые для выполнения антигенпрезентирующей, фагоцитарной и цитотоксической функций, которые отличают нормальные макрофаги. Микроглия также отличается от макрофагов тем, что они гораздо более жестко регулируются в пространстве и времени, чтобы поддерживать точный иммунный ответ. [16]
Еще одним отличием микроглии от других клеток, которые дифференцируются из миелоидных клеток-предшественников, является скорость обновления. Макрофаги и дендритные клетки постоянно изнашиваются и заменяются миелоидными клетками-предшественниками , которые дифференцируются в нужный тип. Из-за гематоэнцефалического барьера организму будет довольно сложно постоянно заменять микроглию. Таким образом, вместо того, чтобы постоянно заменяться миелоидными клетками-предшественниками , микроглия сохраняет свой статус-кво в состоянии покоя, а затем, когда они активируются, они быстро пролиферируют, чтобы поддерживать свою численность. Однако исследования костной химеры показали, что в случаях тяжелой инфекции гематоэнцефалический барьер ослабляется, и микроглия заменяется гематогенными клетками костномозгового происхождения, а именно миелоидными клетками-предшественниками и макрофагами. Как только инфекция уменьшается, связь между периферическими и центральными системами восстанавливается, и в период восстановления и возобновления роста присутствует только микроглия. [35]
Старение [ править ]
Во время травмы микроглия подвергается всплеску митотической активности; за этой пролиферацией следует апоптоз , приводящий к уменьшению количества клеток до исходного уровня. [36] Активация микроглии создает нагрузку на анаболические и катаболические механизмы клеток, в результате чего активированная микроглия погибает раньше, чем неактивированные клетки. [36] Чтобы компенсировать потерю микроглии с течением времени, микроглия подвергается митозу, а клетки-предшественники, полученные из костного мозга , мигрируют в мозг через мозговые оболочки и сосуды. [36]
Накопление незначительных повреждений нейронов, возникающее при нормальном старении, может трансформировать микроглию в увеличенные и активированные клетки. [37] Это хроническое возрастное увеличение активации микроглии и экспрессии IL-1 может способствовать увеличению риска болезни Альцгеймера с возрастом, способствуя образованию нейритных бляшек у восприимчивых пациентов. [37] Повреждение ДНК может способствовать возрастной активации микроглии. Другим фактором может быть накопление конечных продуктов гликирования , которые накапливаются с возрастом. [37] Эти белки сильно устойчивы к протеолитическим процессам и способствуют сшивке белков . [37]
Исследования обнаружили дистрофию (дефектное развитие) микроглии человека. «Эти клетки характеризуются аномалиями цитоплазматической структуры, такими как деразмифицированные, атрофические, фрагментированные или необычно извилистые отростки, часто имеющие сфероидальные или луковицеобразные отеки». [36] Частота дистрофической микроглии увеличивается с возрастом. [36] Сообщалось о дегенерации и смерти микроглии в исследованиях прионной болезни , шизофрении и болезни Альцгеймера, что указывает на то, что разрушение микроглии может быть связано с нейродегенеративными заболеваниями. [36] Сложностью этой теории является тот факт, что трудно отличить «активированную» и «дистрофическую» микроглию в мозге человека. [36]
На мышах было показано, что блокада CD22 восстанавливает гомеостатический микроглиальный фагоцитоз в стареющем мозге. [38]

Клиническое значение

Микроглия — это первичные иммунные клетки центральной нервной системы, подобные периферическим макрофагам. Они реагируют на патогены и травмы, изменяя морфологию и мигрируя к месту инфекции/травмы, где уничтожают патогены и удаляют поврежденные клетки. В рамках своей реакции они секретируют цитокины, хемокины, простагландины и активные формы кислорода, которые помогают направлять иммунный ответ. Кроме того, они играют важную роль в разрешении воспалительной реакции за счет выработки противовоспалительных цитокинов. Микроглия также широко изучалась на предмет ее вредной роли при нейродегенеративных заболеваниях, таких как болезнь Альцгеймера , болезнь Паркинсона , рассеянный склероз , а также сердечных заболеваниях, глаукоме , вирусных и бактериальных инфекциях. Появляется все больше доказательств того, что иммунная дисрегуляция способствует патофизиологии обсессивно-компульсивного расстройства (ОКР) , синдрома Туретта и детских аутоиммунных нейропсихиатрических расстройств, связанных со стрептококковыми инфекциями (PANDAS). . [39]
Поскольку микроглия быстро реагирует даже на незначительные изменения в гомеостазе центральной нервной системы, ее можно рассматривать как сенсоры неврологических дисфункций или расстройств. [40] При патологиях головного мозга фенотип микроглии непременно изменяется. [40] Таким образом, анализ микроглии может быть чувствительным инструментом для диагностики и характеристики нарушений центральной нервной системы в любом образце ткани. [40] В частности, следует оценить плотность микроглиальных клеток, форму клеток, характер распределения, различные фенотипы микроглии и взаимодействие с другими типами клеток. [40]
генетика Чувственная
Микроглиальная сенсома — относительно новая биологическая концепция, которая, по-видимому, играет большую роль в развитии нервной системы и нейродегенерации . Сенсома относится к уникальной группе белковых транскриптов, используемых для восприятия лигандов и микробов . Другими словами, сенсома представляет собой гены, необходимые для белков, используемых для восприятия молекул внутри организма. Сенсому можно анализировать с помощью различных методов, включая qPCR , секвенирование РНК , анализ на микрочипах и прямое секвенирование РНК. Гены, включенные в сенсорный код рецепторов и трансмембранных белков на плазматической мембране , более высоко экспрессируются в микроглии по сравнению с нейронами. Он не включает секретируемые белки или трансмембранные белки, специфичные для мембраносвязанных органелл, таких как ядро , митохондрии и эндоплазматическая сеть . [41] Множество идентифицированных сенсорных генов кодируют рецепторы распознавания образов, однако существует большое разнообразие включенных генов. Микроглия имеет тот же смысл, что и другие макрофаги, однако они содержат 22 уникальных гена, 16 из которых используются для взаимодействия с эндогенными лигандами. Эти различия создают уникальный микроглиальный биомаркер, который включает более 40 генов, включая P2ry12 и HEXB . DAP12 ( TYROBP ), по-видимому, играет важную роль во взаимодействии чувствительных белков, действуя как сигнальный адаптер и регуляторный белок. [41]
Регуляция генов внутри чувств должна иметь возможность меняться, чтобы реагировать на потенциальный вред. Микроглия может взять на себя роль нейропротекции или нейротоксичности , чтобы противостоять этим опасностям. [42] По этим причинам предполагается, что сенсом может играть роль в нейродегенерации. Сенсомные гены, активация которых активируется с возрастом, в основном участвуют в распознавании инфекционных микробных лигандов, тогда как гены, активность которых снижается, в основном участвуют в распознавании эндогенных лигандов. [41] Этот анализ предполагает глиальную специфическую регуляцию, способствующую нейропротекции при естественной нейродегенерации. Это контрастирует со сдвигом в сторону нейротоксичности, наблюдаемым при нейродегенеративных заболеваниях.
Сенсомы также могут играть роль в развитии нервной системы. Инфекция головного мозга в раннем возрасте приводит к тому, что микроглия становится сверхчувствительной к более поздним иммунным раздражителям. При воздействии инфекции происходит активация чувствительных генов, участвующих в нейровоспалении , и подавление генов, участвующих в нейропластичности. [43] Однако способность сенсома изменять развитие нервной системы может помочь в борьбе с болезнями. Удаление CX3CL1 , высокоэкспрессированного чувствительного гена, в моделях синдрома Ретта на грызунах привело к улучшению здоровья и увеличению продолжительности жизни. [44] Снижение уровня регуляции Cx 3 cr1 у людей без синдрома Ретта связано с симптомами, сходными с шизофренией . [45] Это говорит о том, что сенсом не только играет роль в различных нарушениях развития, но также требует жесткой регуляции для поддержания безболезненного состояния.
См. также [ править ]
- Нейроиммунная система
- Список типов клеток человека, полученных из зародышевых листков
- Список различных типов клеток в организме взрослого человека
Ссылки [ править ]
- ^ Жинхо Ф., Лим С., Хоффель Г., Лоу Д., Хубер Т. (2013). «Происхождение и дифференцировка микроглии» . Границы клеточной нейронауки . 7:45 . дои : 10.3389/fncel.2013.00045 . ПМЦ 3627983 . ПМИД 23616747 .
- ^ Дос Сантос С.Е., Медейрос М., Порфирио Х., Таварес В., Пессоа Л., Гринберг Л. и др. (июнь 2020 г.). «Сходная плотность клеток микроглии в структурах мозга и у видов млекопитающих: последствия для функции ткани мозга» . Журнал неврологии . 40 (24): 4622–4643. doi : 10.1523/JNEUROSCI.2339-19.2020 . ПМЦ 7294795 . ПМИД 32253358 .
- ^ Филиано А.Дж., Гадани С.П., Кипнис Дж. (август 2015 г.). «Взаимодействие врожденного и адаптивного иммунитета в развитии и функционировании мозга» . Исследования мозга . 1617 : 18–27. дои : 10.1016/j.brainres.2014.07.050 . ПМЦ 4320678 . ПМИД 25110235 .
- ↑ Перейти обратно: Перейти обратно: а б с Дермицакис И., Манту М.Е., Медицкоу С., Трембле М., Петратос С., Зупи Л. и др. (март 2023 г.). «Происхождение и появление микроглии в ЦНС — интересная (привет) история эксцентрической клетки» . Актуальные проблемы молекулярной биологии . 45 (3): 2609–2628. дои : 10.3390/cimb45030171 . ПМЦ 10047736 . ПМИД 36975541 .
- ^ Кройцберг Г.В. (март 1995 г.). «Микроглия – первая линия защиты при патологиях головного мозга». Арцнаймиттель-Форшунг . 45 (3А): 357–360. ПМИД 7763326 .
- ^ Бушонг Э.А., Мартоне М.Э., Джонс Ю.З., Эллисман М.Х. (январь 2002 г.). «Протоплазматические астроциты в лучистом слое CA1 занимают отдельные анатомические домены» . Журнал неврологии . 22 (1): 183–192. doi : 10.1523/JNEUROSCI.22-01-00183.2002 . ПМЦ 6757596 . ПМИД 11756501 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л Герман Дж., Мацумото Ю., Кройцберг Г.В. (март 1995 г.). «Микроглия: внутренняя иммунноэффекторная клетка головного мозга». Исследования мозга. Обзоры исследований мозга . 20 (3): 269–287. дои : 10.1016/0165-0173(94)00015-H . ПМИД 7550361 . S2CID 22708728 .
- ^ Диссинг-Олесен Л., Лэдеби Р., Нильсен Х.Х., Тофт-Хансен Х., Далмау И., Финсен Б. (октябрь 2007 г.). «Вызванная аксональным поражением пролиферация микроглии и образование кластеров микроглии у мышей». Нейронаука . 149 (1): 112–122. doi : 10.1016/j.neuroscience.2007.06.037 . ПМИД 17870248 . S2CID 36995129 .
- ^ Кирдорф и Принц, J Clin Invest. 2017;127(9): 3201–3209. https://doi.org/10.1172/JCI90602 .
- ↑ Перейти обратно: Перейти обратно: а б Череп С., Посфаи Б., Ленарт Н., Фекете Р., Ласло З.И., Леле З. и др. (январь 2020 г.). «Микроглия контролирует и защищает функции нейронов посредством специализированных соматических пуринергических соединений» . Наука . 367 (6477): 528–537. Бибкод : 2020Sci...367..528C . doi : 10.1126/science.aax6752 . ПМИД 31831638 . S2CID 209343260 .
- ^ Бабе В.М. (1892). «Некоторые особенности гистологических поражений бешенства». Анналы Института Пастера (на французском языке). 6 :209–23.
- ^ Сьерра А, де Кастро Ф, Дель Рио-Ортега Х, Рафаэль Иглесиас-Росас Х, Гарроса М, Кеттенманн Х (ноябрь 2016 г.). «Большой взрыв» для современной глиальной биологии: перевод и комментарии к серии статей Пио дель Рио-Хортеги 1919 года по микроглии». Глия . 64 (11) (опубликовано 16 сентября 2016 г.): 1801–1840. дои : 10.1002/glia.23046 . ПМИД 27634048 . S2CID 3712733 .
- ^ дель Рио Ортега П., Пенфилд В. (1892). «Церебральный рубец: реакция нейроглии и микроглии на раны головного мозга». Бюллетень больницы Джонса Хопкинса . 41 : 278–303.
- ^ дель Рио-Ортега Ф (1937). «Микроглия». Цитология и клеточная патология нервной системы : 481–534.
- ↑ Перейти обратно: Перейти обратно: а б с Верхрацкий А, Батт А (2013). Глиальная физиология и патофизиология . Чичестер: Джон Уайли и сыновья. ISBN 978-1118402054 . [ нужна страница ]
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Алоизи Ф (ноябрь 2001 г.). «Иммунная функция микроглии». Глия . 36 (2): 165–179. дои : 10.1002/glia.1106 . ПМИД 11596125 . S2CID 25410282 .
- ↑ Перейти обратно: Перейти обратно: а б с Кристенсен Р.Н., Ха Б.К., Сан Ф., Бреснахан Дж.К., Битти М.С. (июль 2006 г.). «Каинат вызывает быстрое перераспределение актинового цитоскелета в амебоидной микроглии». Журнал нейробиологических исследований . 84 (1): 170–181. дои : 10.1002/мл.20865 . ПМИД 16625662 . S2CID 34491558 .
- ^ Дэвис Э.Дж., Фостер Т.Д., Томас В.Е. (1994). «Клеточные формы и функции микроглии головного мозга». Бюллетень исследований мозга . 34 (1): 73–78. дои : 10.1016/0361-9230(94)90189-9 . ПМИД 8193937 . S2CID 22596219 .
- ^ Эгген Б.Дж., Радж Д., Ханиш Великобритания, Боддеке Х.В. (сентябрь 2013 г.). «Микроглиальный фенотип и адаптация». Журнал нейроиммунной фармакологии . 8 (4): 807–823. дои : 10.1007/s11481-013-9490-4 . ПМИД 23881706 . S2CID 15283939 .
- ^ Лан X, Хан X, Ли Q, Ян QW, Ван J (июль 2017 г.). «Модуляторы активации и поляризации микроглии после внутримозгового кровоизлияния» . Обзоры природы. Неврология . 13 (7): 420–433. дои : 10.1038/nrneurol.2017.69 . ПМЦ 5575938 . ПМИД 28524175 .
- ^ Елинек Х.Ф., Карперен А., Боссомайер Т., Бьюкен А. (1975). «Дифференциация степеней активации микроглии с помощью фрактального анализа» (PDF) . Комплексность Интернэшнл . 12 (18): 1713–7. Архивировано из оригинала (PDF) 17 декабря 2008 г.
- ↑ Перейти обратно: Перейти обратно: а б Феррер И., Бернет Э., Сориано Э., дель Рио Т., Фонсека М. (1990). «Естественная гибель клеток в коре головного мозга крысы и удаление мертвых клеток транзиторными фагоцитами». Нейронаука . 39 (2): 451–458. дои : 10.1016/0306-4522(90)90281-8 . ПМИД 2087266 . S2CID 23457378 .
- ^ Сайгин Д., Табиб Т., Биттар Х.Э., Валензи Э., Сембрат Дж., Чан С.Ю. и др. (2006). «Эпидемиология, клиника и распространение поражения головного мозга крупного рогатого скота при менингоэнцефалите, вызванном бычьим герпесвирусом-5» . Легочное кровообращение (на португальском языке). 10 (1): 123–32. дои : 10.1590/S0100-736X2006000200010 . ПМК 7052475 . ПМИД 32166015 .
- ↑ Перейти обратно: Перейти обратно: а б Риттер М.Р., Банин Э., Морено С.К., Агилар Э., Доррелл М.И., Фридлендер М. (декабрь 2006 г.). «Миелоидные предшественники дифференцируются в микроглию и способствуют восстановлению сосудов на модели ишемической ретинопатии» . Журнал клинических исследований . 116 (12): 3266–3276. дои : 10.1172/JCI29683 . ПМЦ 1636693 . ПМИД 17111048 .
- ^ Моррис Г.П., Фостер К.Г., Кортни Дж.М., Коллинз Дж.М., Кэшион Дж.М., Браун Л.С. и др. (август 2023 г.). «Микроглия напрямую связана с перицитами центральной нервной системы» . Глия . 71 (8): 1847–1869. дои : 10.1002/glia.24371 . ПМЦ 10952742 . ПМИД 36994950 .
- ^ Чунг В.С., Уэльс, Калифорния, Баррес Б.А. , Стивенс Б. (ноябрь 2015 г.). «Вызывает ли глия синаптические и когнитивные нарушения при заболеваниях?» . Природная неврология . 18 (11): 1539–1545. дои : 10.1038/nn.4142 . ПМЦ 4739631 . ПМИД 26505565 .
- ^ Галлоуэй Д.А., Филлипс А.Е., Оуэн Д.Р., Мур К.С. (2019). «Фагоцитоз в мозге: гомеостаз и болезни» . Границы в иммунологии . 10 :790. дои : 10.3389/fimmu.2019.00790 . ПМК 6477030 . ПМИД 31040847 .
- ^ Вольф А., Херб М., Шрамм М., Лангманн Т. (июнь 2020 г.). «Ось TSPO-NOX1 контролирует запускаемый фагоцитами патологический ангиогенез в глазу» . Природные коммуникации . 11 (1): 2709. Бибкод : 2020NatCo..11.2709W . дои : 10.1038/s41467-020-16400-8 . ПМК 7264151 . ПМИД 32483169 .
- ^ Банно М., Мизуно Т., Като Х., Чжан Г., Каванокучи Дж., Ван Дж. и др. (февраль 2005 г.). «Поглотитель радикалов эдаравон предотвращает окислительную нейротоксичность, вызванную пероксинитритом и активированной микроглией». Нейрофармакология . 48 (2): 283–290. doi : 10.1016/j.neuropharm.2004.10.002 . ПМИД 15695167 . S2CID 25545853 .
- ^ Сотерос Б.М., генеральный директор Sia (ноябрь 2021 г.). «Комплемент и зависимое от микроглии устранение синапсов в развитии мозга» . ПРОВОДА Механизмы заболеваний . 14 (3): e1545. дои : 10.1002/wsbm.1545 . ПМК 9066608 . ПМИД 34738335 .
- ^ Череп С., Шварц А.Д., Посфаи Б., Ласло З.И., Келлермайер А., Корней З. и др. (сентябрь 2022 г.). «Микроглиальный контроль развития нейронов через соматические пуринергические соединения» . Отчеты по ячейкам . 40 (12): 111369. doi : 10.1016/j.celrep.2022.111369 . ПМЦ 9513806 . ПМИД 36130488 . S2CID 252416407 .
- ^ Стэнли Э.Р., Читу В. (июнь 2014 г.). «Передача сигналов рецептора CSF-1 в миелоидных клетках» . Перспективы Колд-Спринг-Харбор в биологии . 6 (6): а021857. doi : 10.1101/cshperspect.a021857 . ПМК 4031967 . ПМИД 24890514 .
- ^ Жинху Ф., Принц М. (июль 2015 г.). «Происхождение микроглии: современные концепции и прошлые споры» . Перспективы Колд-Спринг-Харбор в биологии . 7 (8): а020537. doi : 10.1101/cshperspect.a020537 . ПМЦ 4526747 . ПМИД 26134003 .
- ^ Чжу Х., Ван З., Юй Дж., Ян Х., Хэ Ф., Лю З. и др. (июль 2019 г.). «Роль и механизмы цитокинов при вторичном повреждении головного мозга после внутримозгового кровоизлияния». Прогресс нейробиологии . 178 : 101610. doi : 10.1016/j.pneurobio.2019.03.003 . ПМИД 30923023 . S2CID 85495400 .
- ^ Герман Дж (1996). «Микроглия: сенсор угроз в нервной системе?». Исследования в области вирусологии . 147 (2–3): 79–88. дои : 10.1016/0923-2516(96)80220-2 . ПМИД 8901425 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Стрейт WJ (сентябрь 2006 г.). «Старение микроглии: есть ли у иммунной системы мозга срок годности?». Тенденции в нейронауках . 29 (9): 506–510. doi : 10.1016/j.tins.2006.07.001 . ПМИД 16859761 . S2CID 8874596 .
- ↑ Перейти обратно: Перейти обратно: а б с д Мрак Р.Э., Гриффин В.С. (март 2005 г.). «Глия и их цитокины при прогрессировании нейродегенерации». Нейробиология старения . 26 (3): 349–354. doi : 10.1016/j.neurobiolaging.2004.05.010 . ПМИД 15639313 . S2CID 33152515 .
- ^ Pluvinage JV, Хейни М.С., Смит Б.А., Сан Дж., Ирам Т., Бонанно Л. и др. (апрель 2019 г.). «Блокада CD22 восстанавливает гомеостатический микроглиальный фагоцитоз в стареющем мозге» . Природа . 568 (7751): 187–192. Бибкод : 2019Natur.568..187P . дои : 10.1038/s41586-019-1088-4 . ПМК 6574119 . ПМИД 30944478 .
- ^ Фрик Л., Питтенджер С (2016). «Микроглиальная дисрегуляция при ОКР, синдроме Туретта и PANDAS» . Журнал иммунологических исследований . 2016 : 8606057. doi : 10.1155/2016/8606057 . ПМК 5174185 . ПМИД 28053994 .
- ↑ Перейти обратно: Перейти обратно: а б с д Швабенланд М., Брюк В., Приллер Дж., Стадельманн К., Лассманн Х., Принц М. (декабрь 2021 г.). «Анализ фенотипов микроглии при нейропатологиях: практическое руководство» . Акта Нейропатологика . 142 (6) (опубликовано в октябре 2021 г.): 923–936. дои : 10.1007/s00401-021-02370-8 . ПМЦ 8498770 . ПМИД 34623511 .
 В эту статью включен текст, доступный по лицензии CC BY 4.0 .
В эту статью включен текст, доступный по лицензии CC BY 4.0 . - ↑ Перейти обратно: Перейти обратно: а б с Хикман С.Э., Кингери Н.Д., Осуми Т.К., Боровски М.Л., Ван Л.К., Минс Т.К., Эль Хури Дж. (декабрь 2013 г.). «Чувствительность микроглии, выявленная с помощью прямого секвенирования РНК» . Природная неврология . 16 (12): 1896–1905. дои : 10.1038/nn.3554 . ПМЦ 3840123 . ПМИД 24162652 .
- ^ Блок, М.Л., Зекка, Л. и Хонг, Дж.С. Нейротоксичность, опосредованная микроглией: раскрытие молекулярных механизмов. Нат. Преподобный Нейроски. 8, 57–69 (2007).
- ^ Джи П., Шахтшнейдер К.М., Шук Л.Б., Уокер Ф.Р., Джонсон Р.В. (май 2016 г.). «Периферическая вирусная инфекция индуцирует чувствительные гены микроглии и усиливает активность микроглиальных клеток в гиппокампе новорожденных поросят» . Мозг, поведение и иммунитет . 54 : 243–251. дои : 10.1016/j.bbi.2016.02.010 . ПМЦ 4828316 . ПМИД 26872419 .
- ^ Хориучи М., Смит Л., Маэдзава И., Джин Л.В. (февраль 2017 г.). «Аблация CX 3 CR1 улучшает двигательные и респираторные дисфункции и улучшает выживаемость мышиной модели синдрома Ретта» . Мозг, поведение и иммунитет . 60 : 106–116. дои : 10.1016/j.bbi.2016.02.014 . ПМК 5531048 . ПМИД 26883520 .
- ^ Бергон А., Бельзо Р., Конт М., Пеллетье Ф., Эрве М., Гардинер Э.Дж. и др. (октябрь 2015 г.). «CX3CR1 нарушается в крови и мозге пациентов с шизофренией» (PDF) . Исследования шизофрении . 168 (1–2): 434–443. дои : 10.1016/j.schres.2015.08.010 . ПМИД 26285829 . S2CID 205073822 .
Дальнейшее чтение [ править ]
- Рок Р.Б., Геккер Г., Ху С., Шэн В.С., Чиран М., Локенсгард Дж.Р., Петерсон П.К. (октябрь 2004 г.). «Роль микроглии при инфекциях центральной нервной системы» . Обзоры клинической микробиологии . 17 (4): 942–64, оглавление. doi : 10.1128/CMR.17.4.942-964.2004 . ПМК 523558 . ПМИД 15489356 .
- Хан X, Ли Q, Лан X, Эль-Муфтий L, Рен Х, Ван Дж (сентябрь 2019 г.). «Истощение микроглии липосомами клодроната повышает уровень провоспалительных цитокинов, вызывает активацию астроцитов и повреждает целостность кровеносных сосудов» . Молекулярная нейробиология . 56 (9): 6184–6196. дои : 10.1007/s12035-019-1502-9 . ПМЦ 6684378 . ПМИД 30734229 .
Внешние ссылки [ править ]
- Информация, видео и ресурсы о микроглии на microglia.info
- Домашняя страница микроглии на microglia.net
- Заползая вам в голову: краткое введение в микроглию — обзор журнала Science Creative Quarterly
- «Иммунитетные мусорщики нацелены на бляшки Альцгеймера» . 6 апреля 2007 г.
- Департамент нейробиологии Викиверситета
- Поиск NIF — микроглиальные клетки через информационную систему нейробиологии
| Базы данных органов управления : Национальные |
|---|