Дендрит

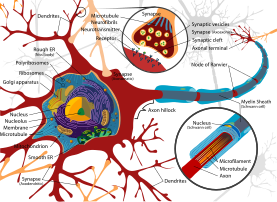
Дендрит полученную от других нервных клеток , (от греческого δένδρον déndron , «дерево») или дендрон — это разветвленное протоплазматическое расширение нервной клетки, которое передает электрохимическую стимуляцию, к телу клетки, или соме , нейрона, из которого выступают дендриты. Электрическая стимуляция передается на дендриты вышестоящими нейронами (обычно через их аксоны ) через синапсы , которые расположены в различных точках дендритного дерева.
Дендриты играют решающую роль в интеграции этих синаптических входов и в определении степени, в которой потенциалы действия производятся нейроном. [1]
Структура и функции [ править ]

Дендриты — один из двух типов протоплазматических выступов, которые выступают из тела клетки нейрона , другой тип — аксон . Аксоны можно отличить от дендритов по нескольким признакам, включая форму, длину и функцию. Дендриты часто сужаются по форме и становятся короче, тогда как аксоны имеют тенденцию сохранять постоянный радиус и могут быть очень длинными. Обычно аксоны передают электрохимические сигналы, а дендриты принимают электрохимические сигналы, хотя у некоторых типов нейронов у определенных видов отсутствуют специализированные аксоны, и они передают сигналы через свои дендриты. [3] Дендриты обеспечивают увеличенную площадь поверхности для приема сигналов от окончаний аксонов других нейронов. [4] Дендрит крупной пирамидной клетки получает сигналы примерно от 30 000 пресинаптических нейронов. [5] Возбуждающие синапсы заканчиваются на дендритных шипиках , крошечных выступах из дендрита с высокой плотностью рецепторов нейромедиаторов . Большинство тормозных синапсов непосредственно контактируют с дендритным стержнем.
Синаптическая активность вызывает локальные изменения электрического потенциала плазматической мембраны дендрита. Это изменение мембранного потенциала будет пассивно распространяться вдоль дендрита, но становится слабее с расстоянием без потенциала действия . Для генерации потенциала действия многие возбуждающие синапсы должны быть активны одновременно, что приводит к сильной деполяризации дендрита и тела клетки ( сомы ). Потенциал действия, который обычно начинается на бугорке аксона , распространяется по всей длине аксона к окончаниям аксона, где он запускает высвобождение нейротрансмиттеров, а также назад в дендрит (ретроградное распространение), обеспечивая важный сигнал для определения времени спайков. -зависимая пластичность (STDP). [4]
Большинство синапсов являются аксодендритами, в них аксон передает сигнал дендриту. Существуют также дендродендритические синапсы, передающие сигналы от одного дендрита к другому. [6] Аутапсом называют синапс , при котором аксон одного нейрона передает сигналы собственному дендриту.
Общая структура дендрита используется для классификации нейронов на мультиполярные , биполярные и униполярные типы. Мультиполярные нейроны состоят из одного аксона и множества дендритных деревьев. Пирамидные клетки представляют собой мультиполярные кортикальные нейроны с пирамидальными телами клеток и крупными дендритами, простирающимися к поверхности коры ( апикальные дендриты ). Биполярные нейроны имеют два основных дендрита на противоположных концах тела клетки. Многие тормозные нейроны имеют такую морфологию. Униполярные нейроны, типичные для насекомых, имеют стебель, отходящий от тела клетки и разделяющийся на две ветви, одна из которых содержит дендриты, а другая - концевые кнопки. У позвоночных сенсорные нейроны, реагирующие на прикосновение или температуру, униполярны. [6] [7] [8] Дендритное ветвление может быть обширным, и в некоторых случаях его достаточно для получения до 100 000 входных сигналов к одному нейрону. [4]
История [ править ]
Термин «дендриты» впервые был использован в 1889 году Вильгельмом Хисом для описания количества более мелких «протоплазматических отростков», прикрепленных к нервной клетке . [9] Немецкому анатому Отто Фридриху Карлу Дейтерсу обычно приписывают открытие аксона, отличив его от дендритов.
Некоторые из первых внутриклеточных записей в нервной системе были сделаны в конце 1930-х годов Кеннетом С. Коулом и Говардом Дж. Кертисом. Швейцарец Рюдольф Альберт фон Кёлликер и немец Роберт Ремак были первыми, кто идентифицировал и охарактеризовал начальный сегмент аксона . Алан Ходжкин и Эндрю Хаксли также использовали гигантский аксон кальмара (1939) и к 1952 году получили полное количественное описание ионной основы потенциала действия, что привело к формулировке модели Ходжкина-Хаксли . За эту работу Ходжкин и Хаксли были совместно удостоены Нобелевской премии в 1963 году. Формулы, описывающие проводимость аксонов, были распространены на позвоночных в уравнениях Франкенхойзера-Хаксли. Луи-Антуан Ранвье был первым, кто описал пробелы или узлы, обнаруженные на аксонах, и за этот вклад эти аксональные особенности теперь обычно называют узлами Ранвье. Сантьяго Рамон-и-Кахаль, испанский анатом, предположил, что аксоны являются выходными компонентами нейронов. [10] Он также предположил, что нейроны представляют собой отдельные клетки, которые сообщаются друг с другом через специализированные соединения или пространства между клетками, теперь известные как синапс. Рамон-и-Кахаль усовершенствовал процесс окрашивания серебром, известный как метод Гольджи, который был разработан его соперником Камилло Гольджи . [11]
дендритов Развитие
На развитие дендритов на дифференцировку могут влиять несколько факторов. К ним относятся модуляция сенсорной информации, загрязнителей окружающей среды, температуры тела и употребления наркотиков. [12] Например, у крыс, выращенных в темноте, было обнаружено уменьшенное количество шипов в пирамидных клетках, расположенных в первичной зрительной коре, и заметное изменение в распределении ветвления дендритов в звездчатых клетках 4-го слоя. [13] Эксперименты, проведенные in vitro и in vivo, показали, что наличие афферентов и входной активности само по себе может модулировать паттерны дифференцировки дендритов. [14]
Мало что известно о процессе, посредством которого дендриты ориентируются in vivo и вынуждены создавать сложную структуру ветвления, уникальную для каждого конкретного класса нейронов. Одной из теорий механизма развития дендритных деревьев является синаптотропная гипотеза. Синаптотропная гипотеза предполагает, что входной сигнал от пресинаптической клетки к постсинаптической (и созревание возбуждающих синаптических входов) в конечном итоге может изменить ход формирования синапсов в дендритных и аксональных ветвях. [15]
Это образование синапсов необходимо для развития нейрональной структуры в функционирующем мозге. Баланс между метаболическими затратами на развитие дендритов и необходимостью закрыть рецептивное поле, по-видимому, определяет размер и форму дендритов. Сложный набор внеклеточных и внутриклеточных сигналов модулирует развитие дендритов, включая факторы транскрипции, взаимодействия рецептор-лиганд, различные сигнальные пути, локальный механизм трансляции, элементы цитоскелета, аванпосты Гольджи и эндосомы. Они способствуют организации дендритов на телах отдельных клеток и размещению этих дендритов в нейронных цепях. Например, было показано, что белок 1, связывающий зипкод β-актина (ZBP1), способствует правильному ветвлению дендритов.
Другие важные факторы транскрипции, участвующие в морфологии дендритов, включают CUT, Abrupt, Collier, Spineless, ACJ6/drifter, CREST, NEUROD1, CREB, NEUROG2 и т. д. Секретируемые белки и рецепторы клеточной поверхности включают нейротрофины и тирозинкиназные рецепторы, BMP7, Wnt/растрёпанный. , EPHB 1–3, Семафорин/плексин-нейропилин, слит-робо, нетрин-измотанный, рилин. Rac, CDC42 и RhoA служат регуляторами цитоскелета, а моторный белок включает KIF5, динеин, LIS1. Было обнаружено, что дендритное разветвление индуцируется в клетках Пуркинье мозжечка Р. веществом [16] Важные секреторные и эндоцитарные пути, контролирующие развитие дендритов, включают DAR3/SAR1, DAR2/Sec23, DAR6/Rab1 и т. д. Все эти молекулы взаимодействуют друг с другом, контролируя морфогенез дендритов, включая приобретение типоспецифического разветвления дендритов, регуляцию размера дендритов и организация дендритов, исходящих от разных нейронов. [1] [17]
Типы дендритных структур [ править ]
Дендритное разветвление, также известное как дендритное ветвление, представляет собой многоэтапный биологический процесс, в ходе которого нейроны формируют новые дендритные деревья и ветви для создания новых синапсов. [1] Дендриты многих организмов имеют разные морфологические закономерности ветвления. Морфология дендритов , такая как плотность ветвей и структура группировки, тесно связана с функцией нейрона. Пороки развития дендритов также тесно коррелируют с нарушением функции нервной системы. [14]
Морфология ветвления может иметь адендритную структуру (не имеющую ветвящейся структуры или не древовидную) или древовидную радиационную структуру. Древовидные структуры разветвления могут быть веретенообразными (когда два дендрита расходятся от противоположных полюсов тела клетки с небольшим количеством ветвей, см. Биполярные нейроны ), сферическими (когда дендриты расходятся частично или во всех направлениях от тела клетки, см. Зернистые клетки мозжечка). ), ламинарный (где дендриты могут либо расходиться плоско, смещенными от тела клетки одним или несколькими стеблями, либо мультиплоскостно, см. горизонтальные клетки сетчатки , ганглиозные клетки сетчатки , амакриновые клетки сетчатки соответственно), цилиндрический (где дендриты расходятся во всех направлениях в цилиндр, дискообразный вид, см. паллидные нейроны ), конический (дендриты расходятся как конус от тела клетки, см. пирамидальные клетки ) или веерный (где дендриты расходятся в виде плоского веера, как в клетках Пуркинье ).
Электрические свойства [ править ]
Структура и разветвление дендритов нейрона, а также наличие и изменение потенциалзависимой ионной проводимости сильно влияют на то, как нейрон интегрирует входные данные от других нейронов. Эта интеграция является как временной, включающей суммирование стимулов, поступающих в быстрой последовательности, так и пространственной, влекущей за собой агрегацию возбуждающих и тормозящих сигналов от отдельных ветвей. [18]
Когда-то считалось, что дендриты просто пассивно передают электрическую стимуляцию. Эта пассивная передача означает, что изменения напряжения, измеряемые в теле клетки, являются результатом активации дистальных синапсов, распространяющих электрический сигнал к телу клетки без помощи потенциалзависимых ионных каналов . Теория пассивного кабеля описывает, как изменения напряжения в определенном месте дендрита передают этот электрический сигнал через систему сближающихся сегментов дендрита разного диаметра, длины и электрических свойств. На основе теории пассивного кабеля можно проследить, как изменения в морфологии дендритов нейрона влияют на мембранное напряжение в теле клетки и, таким образом, как изменения в архитектуре дендритов влияют на общие выходные характеристики нейрона. [19] [20]
Потенциалы действия, инициированные на аксонном холмике, распространяются обратно в дендритное ветвь. Эти потенциалы обратного распространения деполяризуют дендритную мембрану и обеспечивают решающий сигнал для модуляции синапсов и долговременной потенциации . Обратное распространение не является полностью пассивным, а модулируется наличием дендритных потенциалзависимых калиевых каналов . Более того, в некоторых типах нейронов серия потенциалов обратного распространения может индуцировать потенциал действия кальция ( дендритный спайк ) в дендритных зонах инициации. [21] [22]
Пластичность [ править ]
Сами дендриты, по-видимому, способны к пластическим изменениям в течение взрослой жизни животных, в том числе беспозвоночных. [23] Дендриты нейронов имеют различные отсеки, известные как функциональные единицы, которые способны вычислять входящие стимулы. Эти функциональные единицы участвуют в обработке входных данных и состоят из субдоменов дендритов, таких как шипы, ветви или группы ветвей. Следовательно, пластичность, которая приводит к изменениям в структуре дендритов, влияет на коммуникацию и обработку данных в клетке. Во время развития морфология дендритов формируется внутренними программами внутри генома клетки и внешними факторами, такими как сигналы от других клеток. Но во взрослой жизни внешние сигналы становятся более влиятельными и вызывают более значительные изменения в структуре дендритов по сравнению с внутренними сигналами во время развития. У женщин дендритная структура может меняться в результате физиологических состояний, вызванных гормонами в такие периоды, как беременность, лактация и после эстрального цикла. Особенно это заметно в пирамидных клетках области СА1 гиппокампа, где плотность дендритов может варьировать до 30%. [14]
Недавние экспериментальные наблюдения показывают, что адаптация осуществляется в дендритных деревьях нейронов, где время адаптации составляет всего несколько секунд. [24] [25] Некоторые архитектуры машинного обучения, основанные на дендритных деревьях, показали, что они упрощают алгоритм обучения, не влияя на производительность. [26]
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с Урбанска М., Блажейчик М., Яворски Дж. (2008). «Молекулярные основы дендритного разветвления». Acta Neurobiologiae Experimentalis . 68 (2): 264–288. ПМИД 18511961 .
- ^ Перес-Альварес, Альберто; Фири, Бренна С.; О'Тул, Райан Дж.; Ян, Вэй; Арганда-Каррерас, Игнасио; Ламот-Молина, Пол Дж.; Мойерт, Бенджамин; Мор, Мануэль А.; Панзера, Лорен С.; Шульце, Кристиан; Шрайтер, Эрик Р.; Вигерт, Дж. Саймон; Ну и дела, Кристин Э.; Хоппа, Майкл Б.; Эртнер, Томас Г. (18 мая 2020 г.). «Стоп-кадровая визуализация синаптической активности с использованием SynTagMA» . Природные коммуникации . 11 (1): 2464. doi : 10.1038/s41467-020-16315-4 . ISSN 2041-1723 . ПМК 7235013 . ПМИД 32424147 .
- ^ Яу К.В. (декабрь 1976 г.). «Рецептивные поля, геометрия и блок проводимости сенсорных нейронов центральной нервной системы пиявки» . Журнал физиологии . 263 (3): 513–538. дои : 10.1113/jphysicalol.1976.sp011643 . ПМЦ 1307715 . ПМИД 1018277 .
- ^ Jump up to: Перейти обратно: а б с Альбертс Б (2009). Основная клеточная биология (3-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-4129-1 .
- ^ Эяль, Гай; Верхоог, Маттейс Б.; Теста-Сильва, Гильерме; Дейчер, Яир; Бенавидес-Пиччионе, Рут; ДеФелипе, Хавьер; де Кок, Кристиан П.Дж.; Мансвелдер, Хьюберт Д.; Сегев, Идан (29 июня 2018 г.). «Кортикальные пирамидальные нейроны человека: от шипов к шипам через модели» . Границы клеточной нейронауки . 12 : 181. дои : 10.3389/fncel.2018.00181 . ISSN 1662-5102 . ПМК 6034553 . ПМИД 30008663 .
- ^ Jump up to: Перейти обратно: а б Карлсон Н.Р. (2013). Физиология поведения (11-е изд.). Бостон: Пирсон. ISBN 978-0-205-23939-9 .
- ^ Пинель Дж. П. (2011). Биопсихология (8-е изд.). Бостон: Аллин и Бэкон. ISBN 978-0-205-83256-9 .
- ^ Ян Й.Н., Ян ЛИ (май 2010 г.). «Разветвление: механизмы дендритного разветвления» . Обзоры природы. Нейронаука . 11 (5): 316–328. дои : 10.1038/nrn2836 . ПМК 3079328 . ПМИД 20404840 .
- ^ Палец С (1994). Истоки нейробиологии: история исследований функций мозга . Издательство Оксфордского университета. п. 44. ИСБН 9780195146943 . ОСЛК 27151391 .
Нервная клетка с ее непрерывными отростками была описана Отто Фридрихом Карлом Дейтерсом (1834–1863) в работе, завершенной Максом Шульце (1825–1874) в 1865 году, через два года после смерти Дейтерса от брюшного тифа. В этой работе тело клетки было изображено с одним главным «осевым цилиндром» и рядом более мелких «протоплазматических отростков» (см. рис. 3.19). Последние стали известны как «дендриты» — термин, придуманный Вильгельмом Гисом (1831–1904) в 1889 году.
- ^ Дебанн Д., Кампанак Э., Бяловас А., Карлье Э., Алькарас Дж. (апрель 2011 г.). «Физиология аксонов» (PDF) . Физиологические обзоры . 91 (2): 555–602. doi : 10.1152/physrev.00048.2009 . ПМИД 21527732 . Архивировано (PDF) из оригинала 5 мая 2020 г.
- ^ Лопес-Муньос Ф., Бойя Дж., Аламо К. (октябрь 2006 г.). «Теория нейронов, краеугольный камень нейробиологии, к столетию со дня вручения Нобелевской премии Сантьяго Рамону-и-Кахалю». Бюллетень исследований мозга . 70 (4–6): 391–405. дои : 10.1016/j.brainresbull.2006.07.010 . ПМИД 17027775 . S2CID 11273256 .
- ^ МакИвен Б.С. (сентябрь 2010 г.). «Стресс, секс и нейронная адаптация к изменяющейся среде: механизмы ремоделирования нейронов» . Анналы Нью-Йоркской академии наук . 1204 (Дополнение): E38–E59. Бибкод : 2010NYASA1204...38M . дои : 10.1111/j.1749-6632.2010.05568.x . ПМК 2946089 . ПМИД 20840167 .
- ^ Борхес С., Берри М. (июль 1978 г.). «Влияние выращивания в темноте на развитие зрительной коры крыс». Журнал сравнительной неврологии . 180 (2): 277–300. дои : 10.1002/cne.901800207 . ПМИД 659662 . S2CID 42749947 .
- ^ Jump up to: Перейти обратно: а б с Тавосанис Г. (январь 2012 г.). «Дендритная структурная пластичность». Развивающая нейробиология . 72 (1): 73–86. дои : 10.1002/днеу.20951 . ПМИД 21761575 . S2CID 2055017 .
- ^ Клайн Х., Хаас К. (март 2008 г.). «Регуляция развития и пластичности дендритных деревьев с помощью глутаматергических синаптических входов: обзор синаптотрофной гипотезы» . Журнал физиологии . 586 (6): 1509–1517. дои : 10.1113/jphysyol.2007.150029 . ПМК 2375708 . ПМИД 18202093 .
- ^ Балояннис, Ставрос; Коста, Василики; Дереци, Грузия; Мичмизос, Димитриос (1999). « Внутрижелудочковое введение вещества Р увеличивает ветвление дендритов и синаптических поверхностей клеток Пуркинье в мозжечке крысы » . Международный журнал неврологии . 101 (1–4): 89–107 – через Pub Med.
- ^ Перич М., Урбанска А.С., Кравчик П.С., Паробчак К., Яворски Дж. (апрель 2011 г.). «Белок 1, связывающий зипкод, регулирует развитие дендритных ветвей в нейронах гиппокампа» (PDF) . Журнал неврологии . 31 (14): 5271–5285. doi : 10.1523/JNEUROSCI.2387-10.2011 . ПМК 6622686 . ПМИД 21471362 . Архивировано (PDF) из оригинала 22 сентября 2017 г.
- ^ Кандель Э.Р. (2003). Принципы нейронауки (4-е изд.). Кембридж: МакГроу Хилл. ISBN 0-8385-7701-6 .
- ^ Кох С. (1999). Биофизика вычислений: обработка информации в одиночных нейронах . Нью-Йорк [ua]: Оксфордский университет. Нажимать. ISBN 0-19-510491-9 .
- ^ Хойссер М (2008). Дендриты (2-е изд.). Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-856656-4 .
- ^ Гидон, Альберт; Зольник, Тимоти Адам; Фидзинский, Павел; Болдуан, Феликс; Папуци, Афанасия; Пуарази, Панайота; Хольткамп, Мартин; Вида, Имре; Ларкум, Мэтью Эван (03 января 2020 г.). «Дендритные потенциалы действия и вычисления в нейронах коры головного мозга 2/3 слоя человека» . Наука . 367 (6473): 83–87. дои : 10.1126/science.aax6239 . ISSN 0036-8075 . ПМИД 31896716 .
- ^ Ларкум, Мэтью Э.; У, Цзямэн; Дювердин, Сара А.; Гидон, Альберт (2022). «Руководство по дендритным шипам коры млекопитающих in vitro и in vivo» . Нейронаука . 489 : 15–33. doi : 10.1016/j.neuroscience.2022.02.009 . ПМИД 35182699 .
- ^ Мичмизос Д., Кутсураки Э., Аспродини Э., Балояннис С. 2011. Синаптическая пластичность: унифицированная модель для решения некоторых постоянных вопросов. Международный журнал неврологии , 121(6): 289-304. https://www.tandfonline.com/doi/abs/10.3109/00207454.2011.556283
- ^ Ходассман С., Варди Р., Тугендхафт Ю., Голденталь А., Кантер И. (апрель 2022 г.). «Эффективное дендритное обучение как альтернатива гипотезе синаптической пластичности» . Научные отчеты . 12 (1): 6571. Бибкод : 2022NatSR..12.6571H . дои : 10.1038/s41598-022-10466-8 . ПМК 9051213 . ПМИД 35484180 .
- ^ Сарди С., Варди Р., Голдентал А., Шейнин А., Узан Х., Кантер И. (март 2018 г.). «Адаптивные узлы обогащают нелинейное совместное обучение, выходя за рамки традиционной адаптации посредством связей» . Научные отчеты . 8 (1): 5100. Бибкод : 2018НатСР...8.5100С . дои : 10.1038/s41598-018-23471-7 . ПМЦ 5865176 . ПМИД 29572466 .
- ^ Меир Ю, Бен-Ноам I, Цах Ю, Ходассман С, Кантер I (январь 2023 г.). «Обучение древовидной архитектуре превосходит сверточную сеть прямого распространения » Научные отчеты . 13 (1): 962. doi : 10.1038/s41598-023-27986-6 . ПМЦ 9886946 . ПМИД 36717568 .
Внешние ссылки [ править ]
- Гистологическое изображение: 3_09 в Центре медицинских наук Университета Оклахомы - «Слайд 3 Спинной мозг ».
- Дендритное дерево — клеточно-центрированная база данных
- Стереоизображения дендритных деревьев в Kryptopterus электрорецепторных органах