Астроцит
| Астроцит | |
|---|---|
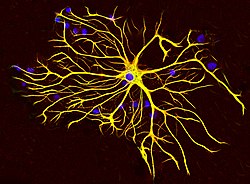 Астроциты из мозга крысы , выращенные в культуре тканей и окрашенные антителами к GFAP (красный) и виментину (зеленый). Оба белка присутствуют в больших количествах в промежуточных нитях этой клетки, поэтому клетка выглядит желтой. Синий материал показывает ДНК, визуализируемую красителем DAPI , и показывает ядро астроцита и других клеток. Изображение предоставлено EnCor Biotechnology Inc. | |
| Подробности | |
| Предшественник | Глиобласт |
| Расположение | Головной и спинной мозг |
| Идентификаторы | |
| латинский | астроцит |
| МеШ | D001253 |
| НейроЛекс ID | сао1394521419 |
| ТД | Х2.00.06.2.00002, Х2.00.06.2.01008 |
| ФМА | 54537 |
| Анатомические термины микроанатомии | |
Астроциты (от древнегреческого ἄστρον , ástron , «звезда» и κύτος , kútos , «полость», «клетка»), также известные под общим названием астроглия , представляют собой характерные звездообразные глиальные клетки в головном и спинном мозге . Они выполняют множество функций, в том числе биохимический контроль эндотелиальных клеток , образующих гематоэнцефалический барьер , [1] обеспечение питательными веществами нервной ткани, поддержание внеклеточного ионного баланса, регуляция мозгового кровотока, а также роль в процессе восстановления и рубцевания головного и спинного мозга после инфекций и травматических повреждений. [2] Доля астроцитов в мозге точно не определена; Исследования показали, что в зависимости от используемой техники подсчета доля астроцитов варьируется в зависимости от региона и колеблется от 20% до примерно 40% всей глии. [3] Другое исследование сообщает, что астроциты являются самым многочисленным типом клеток в мозге. [2] Астроциты являются основным источником холестерина в центральной нервной системе. [4] Аполипопротеин Е транспортирует холестерин от астроцитов к нейронам и другим глиальным клеткам, регулируя передачу сигналов клетками в головном мозге. [4] Астроциты человека более чем в двадцать раз крупнее, чем в мозге грызунов, и вступают в контакт с более чем в десять раз большим количеством синапсов. [5]
Исследования, проведенные с середины 1990-х годов, показали, что астроциты размножают межклеточный Са. 2+ волны на большие расстояния в ответ на стимуляцию и, подобно нейронам, высвобождают передатчики (называемые глиотрансмиттерами ) в Ca 2+ -зависимый характер. [6] Данные свидетельствуют о том, что астроциты также передают сигналы нейронам через Ca. 2+ -зависимое высвобождение глутамата . [7] Такие открытия сделали астроциты важной областью исследований в области нейробиологии .
Структура [ править ]


Астроциты — это подтип глиальных клеток центральной нервной системы . Они также известны как астроцитарные глиальные клетки. Их многочисленные отростки имеют звездчатую форму и охватывают синапсы , образованные нейронами. У человека одна клетка-астроцит может одновременно взаимодействовать с 2 миллионами синапсов. [8] Астроциты классически идентифицируются с помощью гистологического анализа; промежуточных филаментов многие из этих клеток экспрессируют глиальный фибриллярный кислый белок (GFAP). [9] В центральной нервной системе существует несколько форм астроцитов, включая фиброзные (в белом веществе), протоплазматические (в сером веществе) и радиальные . Фиброзная глия обычно расположена в белом веществе, имеет относительно мало органелл и имеет длинные неразветвленные клеточные отростки. Этот тип часто имеет астроцитарные отростки на конце стопы , которые физически соединяют клетки с внешней стороной стенок капилляров , когда они находятся рядом с ними. Протоплазматическая глия наиболее распространена и встречается в тканях серого вещества, содержит большее количество органелл и имеет короткие и сильно разветвленные третичные отростки. Клетки радиальной глии располагаются в плоскостях, перпендикулярных осям желудочков . Один из их отростков примыкает к мягкой мозговой оболочке , а другой глубоко погружен в серое вещество. Радиальная глия в основном присутствует во время развития и играет роль в миграции нейронов . клетки Мюллера сетчатки , которые и глиальные клетки Бергмана коры мозжечка Исключением являются присутствуют еще во взрослом возрасте. Находясь вблизи мягкой мозговой оболочки, все три формы астроцитов посылают отростки, образующие пиа-глиальная мембрана .
Ранние оценки использования энергии в передаче сигналов серого вещества показали, что 95% приходится на нейроны и 5% на астроциты. [10] Однако после обнаружения того, что потенциалы действия были более эффективными, чем первоначально предполагалось, энергетический баланс был скорректирован: 70% для дендритов, 15% для аксонов и 7% для астроцитов. [11] Предыдущие исследования предполагали, что астроциты захватывают синаптический K⁺ исключительно через каналы Kir4.1 . Однако теперь стало понятно, что они также используют Na⁺/K⁺-АТФазу . С учетом этой активной буферизации потребность астроцитов в энергии увеличивается более чем на 200%. Это подтверждается 3D-реконструкциями нейропилей, показывающими схожую плотность митохондрий в обоих типах клеток, а также специфичными для клеток транскриптомными и протеомными данными, а также скоростью цикла трикарбоновых кислот. [12] Поэтому «грамм на грамм астроцитов оказывается таким же дорогим, как нейроны». [12]
Развитие [ править ]

Астроциты – это макроглиальные клетки центральной нервной системы. Астроциты происходят из гетерогенных популяций клеток-предшественников нейроэпителия развивающейся центральной нервной системы. Существует замечательное сходство между хорошо известными генетическими механизмами, определяющими происхождение различных подтипов нейронов , и механизмами макроглиальных клеток. [13] Так же, как и в случае спецификации нейрональных клеток, канонические сигнальные факторы, такие как sonic hedgehog (SHH), фактор роста фибробластов (FGF), WNT и костные морфогенетические белки (BMP), предоставляют информацию о положении развивающимся макроглиальным клеткам посредством градиентов морфогена вдоль дорсо-вентральной и передней частей. –задняя и медиально-латеральная оси. Результирующее формирование паттерна вдоль нервной оси приводит к сегментации нейроэпителия на домены-предшественники (p0, p1, p2, p3 и pMN) для различных типов нейронов в развивающемся спинном мозге. На основании нескольких исследований сейчас считается, что эта модель также применима к спецификации макроглиальных клеток. Исследования, проведенные Хохстимом и его коллегами, показали, что три отдельные популяции астроцитов возникают из доменов p1, p2 и p3. [14] Эти подтипы астроцитов можно идентифицировать по экспрессии различных факторов транскрипции (PAX6, NKX6.1 ) и маркеров клеточной поверхности ( рилин и SLIT1 ). Идентифицированы три популяции подтипов астроцитов: 1) дорсально расположенные астроциты VA1, происходящие из домена p1, экспрессируют PAX6 и рилин; 2) расположенные вентрально астроциты VA3, происходящие из p3, экспрессируют NKX6.1 и SLIT1; и 3) астроцит VA2, расположенный в промежуточном белом веществе, происходящий из домена p2, который экспрессирует PAX6, NKX6.1, рилин и SLIT1. [15] Считается, что после того, как спецификация астроцитов произошла в развивающейся ЦНС, предшественники астроцитов мигрируют в свои конечные позиции в нервной системе до того, как произойдет процесс терминальной дифференцировки .
Функция [ править ]
Астроциты помогают формировать физическую структуру мозга и, как полагают, играют ряд активных ролей, включая секрецию или поглощение нервных трансмиттеров и поддержание гематоэнцефалического барьера. [17] Была предложена концепция трехчастного синапса , подразумевающая тесные взаимоотношения, возникающие в синапсах между пресинаптическим элементом, постсинаптическим элементом и глиальным элементом. [18]
- Структурные : они участвуют в физическом структурировании мозга. Астроциты получили свое название потому, что они имеют форму звезды. Это наиболее распространенные глиальные клетки в головном мозге, тесно связанные с синапсами нейронов. Они регулируют передачу электрических импульсов в мозге.
- Резервный запас гликогена : Астроциты содержат гликоген и способны к глюконеогенезу . Астроциты, расположенные рядом с нейронами лобной коры и гиппокампа, хранят и выделяют глюкозу. Таким образом, астроциты могут подпитывать нейроны глюкозой в периоды высокого потребления глюкозы и ее дефицита. Недавнее исследование на крысах предполагает, что может быть связь между этой деятельностью и физическими упражнениями. [19]
- Метаболическая поддержка : Они снабжают нейроны питательными веществами, такими как лактат .
- Чувствительность к глюкозе : обычно связана с нейронами, обнаружение интерстициального уровня глюкозы в мозге также контролируется астроцитами. Астроциты in vitro активируются низким содержанием глюкозы, и in vivo эта активация увеличивает опорожнение желудка, что приводит к улучшению пищеварения. [20]
- Гематоэнцефалический барьер . Считалось, что концевые ножки астроцитов, окружающие эндотелиальные клетки, помогают поддерживать гематоэнцефалический барьер , и недавние исследования показывают, что они действительно играют существенную роль наряду с плотными соединениями и базальной пластинкой . [ нужна ссылка ] Однако недавно было показано, что активность астроцитов связана с кровотоком в мозге, и именно это на самом деле измеряется с помощью фМРТ. [21] [22]
- Поглощение и высвобождение медиатора : астроциты экспрессируют переносчики плазматической мембраны, такие как переносчики глутамата для нескольких нейротрансмиттеров, включая глутамат , АТФ и ГАМК . Совсем недавно было показано, что астроциты выделяют глутамат или АТФ в везикулярных клетках Ca. 2+ -зависимый характер. [23] (Это оспаривается в отношении астроцитов гиппокампа.) [24]
- Регуляция концентрации ионов во внеклеточном пространстве . Астроциты экспрессируют калиевые каналы с высокой плотностью. Когда нейроны активны, они выделяют калий , увеличивая местную внеклеточную концентрацию. Поскольку астроциты обладают высокой проницаемостью для калия, они быстро очищают избыточное накопление калия во внеклеточном пространстве. [25] Если эта функция нарушена, внеклеточная концентрация калия повысится, что приведет к деполяризации нейронов по уравнению Гольдмана . Хорошо известно, что аномальное накопление внеклеточного калия приводит к эпилептической активности нейронов. [26]
- Модуляция синаптической передачи быстрые изменения в морфологии астроцитов влияют на гетеросинаптическую передачу между нейронами. в супраоптическом ядре гипоталамуса . Было показано, что [27] В гиппокампе астроциты подавляют синаптическую передачу, высвобождая АТФ, которая гидролизуется эктонуклеотидазами образованием с аденозина . Аденозин действует на нейрональные аденозиновые рецепторы, ингибируя синаптическую передачу, тем самым увеличивая динамический диапазон, доступный для LTP . [28]
- Вазомодуляция : астроциты могут служить посредниками в нейрональной регуляции кровотока. [29]
- Стимулирование миелинизирующей активности олигодендроцитов : электрическая активность нейронов заставляет их высвобождать АТФ, что служит важным стимулом для образования миелина. Однако АТФ не действует непосредственно на олигодендроциты . Вместо этого он заставляет астроциты секретировать цитокиновый фактор, ингибирующий лейкоз (LIF), регуляторный белок, который способствует миелинизирующей активности олигодендроцитов. Это говорит о том, что астроциты играют исполнительно-координирующую роль в мозге. [30]
- Восстановление нервной системы : при повреждении нервных клеток в центральной нервной системе астроциты заполняют пространство, образуя глиальный рубец , и могут способствовать восстановлению нервов. Однако роль астроцитов в регенерации ЦНС после травмы недостаточно изучена. Глиальный рубец традиционно описывался как непроницаемый барьер для регенерации, что указывает на его отрицательную роль в регенерации аксонов. Однако недавно в ходе исследований генетической абляции было обнаружено, что астроциты действительно необходимы для регенерации. [31] Что еще более важно, авторы обнаружили, что рубец астроцитов на самом деле необходим для того, чтобы стимулированные аксоны (аксоны, которые были вынуждены расти с помощью нейротрофических добавок) проходили через поврежденный спинной мозг. [31] Астроциты, которые приобрели реактивный фенотип (так называемый астроглиоз , определяемый усилением регуляции, среди прочего, GFAP и виментина). [32] выражение, определение которого все еще обсуждается) на самом деле может быть токсичным для нейронов, высвобождая сигналы, которые могут убить нейроны. [33] Однако предстоит еще многое сделать для выяснения их роли в повреждении нервной системы.
- Долгосрочное потенцирование : ученые спорят о том, интегрируют ли астроциты обучение и память в гиппокампе. Недавно было показано, что приживление человеческих глиальных клеток-предшественников в мозг зарождающихся мышей приведет к дифференцировке клеток в астроциты. После дифференцировки эти клетки увеличивают LTP и улучшают память у мышей. [34]
- Циркадные часы : одних астроцитов достаточно, чтобы управлять молекулярными колебаниями в СХЯ и циркадным поведением у мышей, и, таким образом, они могут автономно инициировать и поддерживать сложное поведение млекопитающих. [35]
- Переключение нервной системы . На основании данных, перечисленных ниже, недавно было высказано предположение: [36] что макроглия (и астроциты в частности) действуют одновременно как конденсатор нейромедиаторов с потерями и как логический переключатель нервной системы. Т.е. макроглия либо блокирует, либо обеспечивает распространение стимула по нервной системе, в зависимости от состояния их мембран и уровня стимула.
| Тип доказательства | Описание | Ссылки |
|---|---|---|
| Доказательства кальция | Волны кальция появляются только при превышении определенной концентрации нейромедиатора. | [38] [39] [40] |
| Электрофизиологические данные | Отрицательная волна появляется, когда уровень стимула пересекает определенный порог. Форма электрофизиологического ответа отличается и имеет противоположную полярность по сравнению с характерным нервным ответом, что позволяет предположить, что в процесс могут быть вовлечены и другие клетки, а не нейроны. | [41] [42] |
| Психофизические доказательства | Отрицательный электрофизиологический ответ сопровождается действиями по принципу «все или ничего». Умеренная отрицательная электрофизиологическая реакция появляется при осознанных логических решениях, таких как задачи восприятия. Интенсивная резкая отрицательная волна появляется при эпилептических припадках и при рефлексах. | [41] [44] [42] [43] |
| Тесты на поглощение глутамата на основе радиоактивности | Тесты на поглощение глутамата показывают, что астроциты перерабатывают глутамат со скоростью, которая первоначально пропорциональна концентрации глутамата. Это подтверждает модель утечки конденсатора, где «утечка» представляет собой обработку глутамата глутаминсинтетазой глии. Кроме того, те же тесты указывают на уровень насыщения, после которого уровень поглощения нейромедиатора перестает расти пропорционально концентрации нейромедиатора. Последнее подтверждает существование порога. Графики, показывающие эти характеристики, называются графиками Михаэлиса-Ментен. | [45] |
Астроциты соединяются щелевыми контактами , образуя электрически связанный (функциональный) синцитий . [46] Из-за этой способности астроцитов общаться со своими соседями изменения в активности одного астроцита могут иметь последствия для активности других, которые довольно далеки от исходного астроцита.
Приток Ca 2+ ионов в астроциты – это существенное изменение, которое в конечном итоге генерирует волны кальция. Поскольку этот приток напрямую вызван увеличением притока крови к мозгу, волны кальция считаются своего рода функцией гемодинамического ответа . Увеличение внутриклеточной концентрации кальция может распространяться наружу через этот функциональный синцитий. Механизмы распространения волн кальция включают диффузию ионов кальция и IP3 через щелевые контакты и внеклеточную передачу сигналов АТФ . [47] Повышение уровня кальция является основной известной осью активации астроцитов и необходимо и достаточно для некоторых типов высвобождения астроцитарного глутамата. [48] Учитывая важность передачи сигналов кальция в астроцитах, были разработаны жесткие регуляторные механизмы для прогрессирования пространственно-временной передачи сигналов кальция. С помощью математического анализа было показано, что локализованный приток Ca 2+ ионов приводит к локализованному повышению цитозольной концентрации Ca 2+ ионы. [49] Кроме того, цитозольный Ca 2+ накопление не зависит от внутриклеточного потока кальция и зависит от Ca 2+ обмен черезмембрана, диффузия кальция в цитозоле, геометрия клетки, возмущение внеклеточного кальция и начальные концентрации. [49]
Трехсторонний синапс [ править ]
В дорсальном роге спинного мозга активированные астроциты способны реагировать практически на все нейротрансмиттеры. [50] и при активации высвобождают множество нейроактивных молекул, таких как глутамат , АТФ , оксид азота (NO) и простагландины (ПГ), что, в свою очередь, влияет на возбудимость нейронов. Тесная связь между астроцитами и пресинаптическими и постсинаптическими окончаниями, а также их способность интегрировать синаптическую активность и высвобождать нейромодуляторы получила название трехстороннего синапса . [18] Синаптическая модуляция астроцитов происходит из-за этой трехчастной ассоциации.
Исследование 2024 года показало, что астроциты, ранее недостаточно изученные клетки мозга, могут играть ключевую роль в продлении бодрствования без негативного воздействия на когнитивные функции и здоровье. [51]
Клиническое значение
Астроцитомы [ править ]
Астроцитомы — это первичные внутричерепные опухоли , развивающиеся из астроцитов. Также возможно, что глиальные предшественники или нейральные стволовые клетки могут давать начало астроцитомам. Эти опухоли могут возникать во многих частях головного и/или спинного мозга. Астроцитомы делятся на две категории: низкой степени (I и II) и высокой степени (III и IV). Опухоли низкой степени злокачественности чаще встречаются у детей, а опухоли высокой степени злокачественности чаще встречаются у взрослых. Злокачественные астроцитомы более распространены среди мужчин, что способствует худшей выживаемости. [52]
Пилоцитарные астроцитомы относятся к опухолям I степени. Они считаются доброкачественными и медленно растущими опухолями. Пилоцитарные астроцитомы часто имеют кистозные части, заполненные жидкостью, и узелок, который представляет собой солидную часть. Большинство из них расположены в мозжечке. Таким образом, большинство симптомов связаны с трудностями равновесия или координации. [52] Они также чаще встречаются у детей и подростков. [53]
Фибриллярные астроцитомы относятся к опухолям II степени. Они растут относительно медленно, поэтому обычно считаются доброкачественными, но проникают в окружающие здоровые ткани и могут стать злокачественными . Фибриллярные астроцитомы обычно возникают у молодых людей, у которых часто наблюдаются судороги. [53]
Анапластические астроцитомы относятся к злокачественным опухолям III степени. Они растут быстрее, чем опухоли более низкой степени злокачественности. Анапластические астроцитомы рецидивируют чаще, чем опухоли более низкой степени злокачественности, поскольку их склонность к распространению в окружающие ткани затрудняет их полное удаление хирургическим путем. [52]
Глиобластома — это рак IV степени, который может возникать из астроцитов или существующей астроцитомы. Примерно 50% всех опухолей головного мозга представляют собой глиобластомы. Глиобластомы могут содержать несколько типов глиальных клеток, включая астроциты и олигодендроциты . Глиобластомы обычно считаются наиболее инвазивным типом глиальной опухоли, поскольку они быстро растут и распространяются на близлежащие ткани. Лечение может быть сложным, поскольку один тип опухолевых клеток может погибнуть в ответ на определенное лечение, в то время как другие типы клеток могут продолжать размножаться. [52]
Нарушения нервно-психического развития [ править ]
Астроциты стали важными участниками различных нарушений нервного развития . Эта точка зрения утверждает, что дисфункция астроцитов может привести к неправильной работе нейронных сетей, что лежит в основе некоторых психических расстройств, таких как расстройства аутистического спектра и шизофрения . [54] [5]
Хроническая боль [ править ]
В нормальных условиях проведение боли начинается с некоторого вредного сигнала, за которым следует потенциал действия, переносимый ноцицептивными (чувствительными к боли) афферентными нейронами, которые вызывают возбуждающие постсинаптические потенциалы (ВПСП) в дорсальном роге спинного мозга. Затем это сообщение передается в кору головного мозга , где мы переводим эти ВПСП в «боль». С момента открытия передачи сигналов астроциты-нейроны наше понимание проведения боли резко усложнилось. Обработка боли больше не рассматривается как повторяющаяся передача сигналов от тела к мозгу, а как сложная система, которая может регулироваться как вверх, так и вниз под действием ряда различных факторов. Одним из факторов, находящихся в центре внимания недавних исследований, является синапс, усиливающий боль, расположенный в дорсальном роге спинного мозга, и роль астроцитов в инкапсуляции этих синапсов. Гарнизон и коллеги [55] были первыми, кто предположил наличие связи, когда обнаружили корреляцию между гипертрофией астроцитов в дорсальном роге спинного мозга и гиперчувствительностью к боли после повреждения периферических нервов, которая обычно считается индикатором активации глии после травмы. Астроциты обнаруживают активность нейронов и могут выделять химические передатчики, которые, в свою очередь, контролируют синаптическую активность. [50] [56] [57] Раньше считалось, что гипералгезия модулируется высвобождением вещества Р и возбуждающих аминокислот (EAA), таких как глутамат , из пресинаптических афферентных нервных окончаний в дорзальных рогах спинного мозга. Далее следует активация AMPA (α-амино-3-гидрокси-5-метил-4-изоксазолпропионовой кислоты), NMDA (N-метил-D-аспартата) и каинатных подтипов ионотропных глутаматных рецепторов . Именно активация этих рецепторов усиливает болевой сигнал, идущий по спинному мозгу. Эта идея, хотя и верна, но представляет собой чрезмерное упрощение передачи боли. ряд других нейромедиаторов и нейромодуляторов, таких как пептид, связанный с геном кальцитонина (CGRP), аденозинтрифосфат (АТФ), нейротрофический фактор головного мозга (BDNF), соматостатин , вазоактивный кишечный пептид (VIP), галанин и вазопрессин. Синтезируется целый и высвобождается в ответ на вредные раздражители . В дополнение к каждому из этих регуляторных факторов, несколько других взаимодействий между передающими боль нейронами и другими нейронами дорсального рога оказывают дополнительное влияние на пути боли.
состояния постоянной боли Два
После стойкого повреждения периферических тканей происходит высвобождение нескольких факторов из поврежденной ткани, а также из задних рогов спинного мозга. Эти факторы увеличивают чувствительность нейронов проекции боли задних рогов на последующие стимулы, называемые «спинальной сенсибилизацией», тем самым усиливая болевой импульс в мозг. Высвобождение глутамата, вещества P и пептида, связанного с геном кальцитонина (CGRP), опосредует активацию NMDAR (первоначально молчащую, поскольку он блокируется Mg2+), тем самым способствуя деполяризации постсинаптических нейронов, передающих боль (PTN). Кроме того, активация передачи сигналов IP3 и MAPK (митоген-активируемых протеинкиназ), таких как ERK и JNK , приводит к увеличению синтеза воспалительных факторов, которые изменяют функцию переносчика глутамата. ERK также дополнительно активирует AMPAR и NMDAR в нейронах. Ноцицепция дополнительно сенсибилизируется за счет ассоциации АТФ и вещества P с соответствующими рецепторами (P 2 X 3 ) и рецептором нейрокинина 1 (NK1R), а также активацией метаботропные рецепторы глутамата и высвобождение BDNF. Постоянное присутствие глутамата в синапсе в конечном итоге приводит к нарушению регуляции GLT1 и GLAST , важнейших переносчиков глутамата в астроциты. Продолжающееся возбуждение также может индуцировать активацию ERK и JNK, что приводит к высвобождению нескольких воспалительных факторов.
Поскольку неприятная боль сохраняется, сенсибилизация позвоночника создает транскрипционные изменения в нейронах заднего рога, которые приводят к изменению функции на продолжительные периоды времени. Мобилизация Ca 2+ из внутренних запасов является результатом постоянной синаптической активности и приводит к высвобождению глутамата, АТФ, фактора некроза опухоли-α (TNF-α), интерлейкина 1β ( IL-1β ), IL-6, оксида азота (NO) и простагландина E2. (ПГЕ2). Активированные астроциты также являются источником матриксной металлопротеиназы 2 ( MMP2 ), которая индуцирует расщепление про-IL-1β и поддерживает активацию астроцитов. В этом хроническом сигнальном пути р38 активируется в результате передачи сигналов IL-1β , и присутствует присутствие хемокинов, которые активизируют их рецепторы. В ответ на повреждение нервов белки теплового шока (HSP) высвобождаются и могут связываться с соответствующими TLR , что приводит к дальнейшей активации.
Другие патологии [ править ]
Другие клинически значимые патологии, связанные с астроцитами, включают астроглиоз и астроцитопатию . Примеры их включают рассеянный склероз , анти-AQP4+ оптикомиелит , энцефалит Расмуссена , болезнь Александера и боковой амиотрофический склероз . [58] Исследования показали, что астроциты могут быть вовлечены в нейродегенеративные заболевания , такие как болезнь Альцгеймера . [59] [60] болезнь Паркинсона , [61] Болезнь Хантингтона , Заикание [62] и боковой амиотрофический склероз , [63] и при острых травмах головного мозга, таких как внутримозговое кровоизлияние. [64] и черепно-мозговая травма. [65]
-позитивные астроциты и дисфункция мозга Гомори
Тип астроцитов с патологией, связанной со старением, был описан за последние пятьдесят лет. Астроциты этого подтипа обладают выступающими цитоплазматическими гранулами, которые интенсивно окрашиваются гематоксилином хромалюмовых квасцов Гомори и, следовательно, называются Гомори-позитивными (GP) астроцитами. Их можно найти по всему мозгу, но больше всего их в обонятельных луковицах, медиальной хабенуле, зубчатой извилине гиппокампа, дугообразном ядре гипоталамуса и в дорсальном мозговом веществе, сразу под постремной областью. [66]
Гомори-положительные цитоплазматические гранулы происходят из поврежденных митохондрий, поглощенных лизосомами. [67] Цитоплазматические гранулы содержат непереваренные остатки митохондриальных структур. Это содержимое включает связанные с гемом атомы меди и железа, оставшиеся от митохондриальных ферментов. [68] Эти химические вещества отвечают за псевдопероксидазную активность Гомори-положительных гранул, которую можно использовать для окрашивания этих гранул. Считается, что окислительный стресс является причиной повреждения этих астроцитов. [69] Однако точная природа этого стресса неясна.
Области мозга, обогащенные Гомори-положительными астроцитами, также содержат субпопуляцию специализированных астроцитов, которые синтезируют белок 7, связывающий жирные кислоты (FABP7). Действительно, астроциты гипоталамуса, синтезирующие FABP7, также обладают Гомори-позитивными гранулами. [70] Таким образом, связь между этими двумя глиальными особенностями очевидна. Недавние данные показали, что астроциты, но не нейроны, обладают митохондриальными ферментами, необходимыми для метаболизма жирных кислот, и что возникающий в результате окислительный стресс может повредить митохондрии. [71] Таким образом, повышенное поглощение и окисление жирных кислот в глии, содержащей FABP7, вероятно, вызовет окислительный стресс и повреждение митохондрий в этих клетках. Кроме того, недавно было показано, что белки FABP взаимодействуют с белком под названием синуклеин, вызывая повреждение митохондрий. [72]
роль патофизиологии в Возможная
Астроциты могут переносить митохондрии в соседние нейроны для улучшения функции нейронов. [73] Поэтому вполне вероятно, что повреждение митохондрий астроцитов, наблюдаемое в астроцитах GP, может влиять на активность нейронов.
С возрастом наблюдается снижение ряда функций гипоталамуса, что может быть связано с астроцитами GP. Например, астроциты GP находятся в тесном контакте с нейронами, которые вырабатывают нейромедиатор, называемый дофамином, как в гипоталамусе крысы, так и человека. [74] Дофамин, вырабатываемый этими нейронами, переносится в близлежащий гипофиз, чтобы ингибировать высвобождение гормона пролактина из гипофиза. Активность дофаминергических нейронов снижается с возрастом, что приводит к повышению уровня пролактина в крови, что может спровоцировать рак молочной железы. [75] Связанные со старением изменения в функции астроцитов могут способствовать этому изменению дофаминергической активности.
Астроциты FABP7+ находятся в тесном контакте с нейронами дугообразного ядра гипоталамуса, которые реагируют на гормон лептин, вырабатываемый жировыми клетками. Лептин-чувствительные нейроны регулируют аппетит и массу тела. Астроциты FABP7+ регулируют чувствительность этих нейронов к лептину. Таким образом, повреждение митохондрий в этих астроцитах может изменить функцию чувствительных к лептину нейронов и способствовать связанному со старением нарушению регуляции питания и веса тела. [76]
Астроциты GP также могут участвовать в гипоталамической регуляции общего метаболизма глюкозы. Недавние данные показывают, что астроциты функционируют как сенсоры глюкозы и оказывают решающее влияние на реактивность нейронов на изменения внеклеточной глюкозы. [77] Астроциты GP обладают высокопроизводительными белками-переносчиками глюкозы типа GLUT2 и, по-видимому, модулируют реакции нейронов на глюкозу. [78] Клетки гипоталамуса контролируют уровень глюкозы в крови и оказывают влияние на уровень глюкозы в крови посредством измененного воздействия на вегетативные цепи, иннервирующие клетки печени и мышц.
Важность астроцитов в связанных со старением нарушениях метаболизма глюкозы недавно была проиллюстрирована исследованиями на животных, больных диабетом. Было показано, что однократное введение белка, называемого фактором роста фибробластов-1, в гипоталамус навсегда нормализует уровень глюкозы в крови у грызунов с диабетом. Это замечательное лекарство от сахарного диабета осуществляется при помощи астроцитов. Наиболее известные гены, активируемые обработкой FGF-1, включают гены, ответственные за синтез FABP6 и FABP7 астроцитами. [79] Эти данные подтверждают важность астроцитов FABP7+ для контроля уровня глюкозы в крови. Дисфункция FABP7+/Гомори-положительных астроцитов может способствовать возрастному развитию сахарного диабета.
Астроциты GP также присутствуют в зубчатой извилине гиппокампа как в мозге грызунов, так и в мозге человека. [80] Гиппокамп претерпевает серьезные дегенеративные изменения во время старения при болезни Альцгеймера. Причины этих дегенеративных изменений в настоящее время горячо обсуждаются. Недавнее исследование показало, что уровни глиальных белков, а НЕ нейрональных белков, являются наиболее аномальными при болезни Альцгеймера. Наиболее сильно поражается глиальный белок FABP5. [81] Другое исследование показало, что 100% астроцитов гиппокампа, содержащих FABP7, также содержат FABP5. [82] Эти данные позволяют предположить, что FABP7+/Гомори-положительные астроциты могут играть роль в развитии болезни Альцгеймера. Изменение глиальной функции в этой области может поставить под угрозу функцию нейронов зубчатой извилины, а также функцию аксонов, оканчивающихся в зубчатой извилине. Многие такие аксоны берут начало в латеральной энторинальной коре, которая является первой областью мозга, демонстрирующей дегенерацию при болезни Альцгеймера. Таким образом, патология астроцитов в гиппокампе может внести вклад в патологию болезни Альцгеймера.
Исследования [ править ]
Исследование, проведенное в ноябре 2010 года и опубликованное в марте 2011 года, было проведено группой ученых из Университета Рочестера и Медицинской школы Университета Колорадо . Они провели эксперимент, пытаясь восстановить травму центральной нервной системы взрослой крысы путем замены глиальных клеток. Когда глиальные клетки были инъецированы в поврежденный участок спинного мозга взрослой крысы, астроциты были получены путем воздействия на глиальные клетки-предшественники человека костного морфогенетического белка (костный морфогенетический белок важен, поскольку считается, что он создает тканевую архитектуру по всему телу). Таким образом, в сочетании с костным белком и глиальными клетками человека они способствовали значительному восстановлению осознанной постановки стопы, росту аксонов и очевидному увеличению выживаемости нейронов в пластинках спинного мозга . С другой стороны, человеческие глиальные клетки-предшественники и астроциты, образующиеся из этих клеток при контакте с цилиарными нейротрофическими факторами, не смогли способствовать выживанию нейронов и поддержке роста аксонов в месте повреждения. [83]
В одном исследовании, проведенном в Шанхае, использовались два типа культур нейронов гиппокампа : в одной культуре нейрон выращивался из слоя астроцитов, а другая культура не контактировала ни с какими астроцитами, но вместо этого им вводили глиальную кондиционированную среду (GCM). , который в большинстве случаев подавляет быстрый рост культивируемых астроцитов в мозге крыс. В своих результатах они смогли увидеть, что астроциты играли непосредственную роль в долговременной потенциации со смешанной культурой (то есть культурой, выращенной из слоя астроцитов), но не с культурами GCM. [84]
Исследования показали, что астроциты играют важную функцию в регуляции нейральных стволовых клеток . Исследования Института глазных исследований Шепенса в Гарварде показывают, что человеческий мозг изобилует нервными стволовыми клетками, которые удерживаются в спящем состоянии благодаря химическим сигналам (эфрин-А2 и эфрин-А3), исходящим от астроцитов. Астроциты способны активировать стволовые клетки для трансформации в рабочие нейроны, подавляя высвобождение эфрина-А2 и эфрина-А3 . [85]
В исследовании, опубликованном в журнале Nature Biotechnology за 2011 г. [86] группа исследователей из Университета Висконсина сообщает, что им удалось превратить эмбриональные и индуцированные стволовые клетки человека в астроциты.
Исследование 2012 года [87] Исследования влияния марихуаны на кратковременную память показали, что ТГК активирует рецепторы CB1 астроцитов, которые вызывают рецепторов АМРА удаление из мембран связанных нейронов.
Исследование 2023 года [88] показали, что астроциты также играют активную роль в болезни Альцгеймера . Точнее, когда астроциты становятся реактивными, они вызывают патологическое воздействие бета-амилоида на последующее фосфорилирование и отложение тау, что весьма вероятно приведет к ухудшению когнитивных функций. Также в 2023 году было объявлено об открытии специализированных астроцитов, опосредующих глутаматергическую глиотрансмиссию в центральной нервной системе . [89]
Классификация [ править ]
Существует несколько различных способов классификации астроцитов.
и антигенный Происхождение фенотип
Они были установлены классической работой Raff et al. в начале 1980-х годов на зрительных нервах Крысы.
- Тип 1: Антигенно Ran2 + , ГФАП + , ФГФР3 + , А2В5 − , что напоминало «астроциты 1-го типа» зрительного нерва крысы 7-го дня постнатального периода. Они могут возникать из трипотентных клеток-предшественников, ограниченных глией (GRP), но не из бипотенциальных клеток O2A/OPC (олигодендроцитов, предшественников астроцитов 2 типа, также называемых клетками-предшественниками олигодендроцитов ).
- Тип 2: антигенно A2B5. + , ГФАП + , ФГФР3 − , Ран 2 − . Эти клетки могут развиваться in vitro либо из трипотенциального GRP (вероятно, на стадии O2A), либо из бипотенциальных клеток O2A (которые некоторые люди{{ [90] }} думаю, что, в свою очередь, могли быть получены из GRP) или in vivo, когда эти клетки-предшественники трансплантированы в места поражения (но, вероятно, не в нормальном развитии, по крайней мере, не в зрительном нерве крысы ). Астроциты типа 2 являются основным астроцитарным компонентом в постнатальных культурах зрительного нерва, которые генерируются клетками O2A, выращенными в присутствии фетальной телячьей сыворотки, но, как полагают, не существуют in vivo . [91]
Анатомическая классификация [ править ]
- Протоплазматический: содержится в сером веществе и имеет множество ветвящихся отростков, конечные ножки которых охватывают синапсы . Некоторые протоплазматические астроциты генерируются мультипотентными клетками-предшественниками субвентрикулярной зоны . [92] [93]
- Гёмёри-позитивные астроциты. Это подмножество протоплазматических астроцитов, которые содержат многочисленные цитоплазматические включения или гранулы, которые положительно окрашиваются гематоксилином хром-квасцов Гёмори . Сейчас известно, что эти гранулы образуются из остатков дегенерирующих митохондрий, попавших в лизосомы. [94] Некоторый тип окислительного стресса, по-видимому, ответственен за повреждение митохондрий в этих специализированных астроцитах. Гёмёри-положительные астроциты гораздо более распространены в дугообразном ядре гипоталамуса . и в гиппокампе, чем в других областях мозга Они могут играть роль в регуляции реакции гипоталамуса на глюкозу. [95] [96]
- Фиброзные: встречаются в белом веществе и имеют длинные тонкие неразветвленные отростки, конечные ножки которых охватывают узлы Ранвье . Некоторые фиброзные астроциты образуются радиальной глией . [97] [98] [99] [100] [101]
/ Классификация рецепторов транспортеров
- Тип GluT: экспрессируют переносчики глутамата ( EAAT1 / SLC1A3 и EAAT2 / SLC1A2 ) и отвечают на синаптическое высвобождение глутамата токами транспортера. Функция и доступность EAAT2 модулируются TAAR1 , внутриклеточным рецептором в астроцитах человека. [102]
- Тип GluR: они экспрессируют рецепторы глутамата (в основном типа mGluR и AMPA ) и отвечают на синаптическое высвобождение глутамата с помощью канально-опосредованных токов и IP3 -зависимого Ca. 2+ переходные процессы.
См. также [ править ]
- Бергмановский глиоз
- Гемистоцит
- Питуицит
- Список типов клеток человека, полученных из зародышевых листков
- Список различных типов клеток в организме взрослого человека
Ссылки [ править ]
- ^ Сузуки, Ясухиро; Са, Кила; Очиаи, Эри; Маллинз, Джереми; Йолкен, Роберт; Халонен, Сандра К. (2014). «Церебральный токсоплазмоз». Токсоплазма гондии . Эльзевир. стр. 755–796. дои : 10.1016/b978-0-12-396481-6.00023-4 . ISBN 978-0-12-396481-6 .
Астроциты являются доминирующими глиальными клетками в головном мозге, и многочисленные исследования показывают, что они играют центральную роль во внутримозговом иммунном ответе на T. gondii в головном мозге.
- ^ Jump up to: а б Фриман, MR; Рович, Д.Х. (30 октября 2013 г.). «Развивающиеся концепции глиогенеза: взгляд назад и вперед, на следующие 25 лет» . Нейрон . 80 (3): 613–23. дои : 10.1016/j.neuron.2013.10.034 . ПМК 5221505 . ПМИД 24183014 .
- ^ Верхрацкий А, Батт А.М. (2013). «Цифры: сколько глиальных клеток в мозге?». Глиальная физиология и патофизиология . Джон Уайли и сыновья. стр. 93–96. ISBN 978-0-470-97853-5 .
- ^ Jump up to: а б Ван, Хао; Кулас, Джошуа А.; Феррис, Хизер А.; Хансен, Скотт Б. (14 октября 2020 г.). «Регуляция производства бета-амилоида в нейронах с помощью холестерина, полученного из астроцитов» . bioRxiv : 2020.06.18.159632. дои : 10.1101/2020.06.18.159632 . S2CID 220044671 .
- ^ Jump up to: а б Слоан С.А., Баррес Б.А. (август 2014 г.). «Механизмы развития астроцитов и их вклад в нарушения нервного развития» . Современное мнение в нейробиологии . 27 : 75–81. дои : 10.1016/j.conb.2014.03.005 . ПМЦ 4433289 . ПМИД 24694749 .
- ^ «Роль астроцитов в центральной нервной системе» . Проверено 27 июля 2018 г.
- ^ Фиакко Т.А., Агулхон С., Маккарти К.Д. (октябрь 2008 г.). «Отличие физиологии астроцитов от фармакологии». Ежегодный обзор фармакологии и токсикологии . 49 (1): 151–74. doi : 10.1146/annurev.pharmtox.011008.145602 . ПМИД 18834310 .
- ^ Филдс Р.Д., Араке А., Йохансен-Берг Х., Лим С.С., Линч Г., Нейв К.А. и др. (октябрь 2014 г.). «Глиальная биология в обучении и познании» . Нейробиолог . 20 (5): 426–31. дои : 10.1177/1073858413504465 . ПМЦ 4161624 . ПМИД 24122821 .
- ^ Венкатеш К., Шрикант Л., Венгамма Б., Чандрасекхар С., Сандживкумар А., Мулешвара Прасад Б.К., Сарма П.В. (2013). «Дифференцировка in vitro культивируемых клеток CD34+ человека в астроциты» . Неврология Индия . 61 (4): 383–8. дои : 10.4103/0028-3886.117615 . ПМИД 24005729 .
- ^ Эттвелл, Дэвид; Лафлин, Саймон Б. (2001). «Энергетический бюджет для передачи сигналов в сером веществе мозга» . Журнал церебрального кровотока и метаболизма . 21 (10): 1133–1145. дои : 10.1097/00004647-200110000-00001 . ISSN 0271-678X . ПМИД 11598490 .
- ^ Харрис, Джулия Дж.; Жоливе, Рено; Эттвелл, Дэвид (2012). «Использование и снабжение синаптической энергией» . Нейрон . 75 (5): 762–777. дои : 10.1016/j.neuron.2012.08.019 . ISSN 0896-6273 . ПМИД 22958818 . S2CID 14988407 .
- ^ Jump up to: а б Баррос, LF (2022). «Насколько дороги астроциты?» . Журнал церебрального кровотока и метаболизма . 42 (5): 738–745. дои : 10.1177/0271678x221077343 . ISSN 0271-678X . ПМК 9254036 . ПМИД 35080185 .
- ^ Рович Д.Х., Кригштайн, Арканзас (ноябрь 2010 г.). «Генетика развития спецификации глиальных клеток позвоночных». Природа . 468 (7321): 214–22. Бибкод : 2010Natur.468..214R . дои : 10.1038/nature09611 . ПМИД 21068830 . S2CID 573477 .
- ^ Мурояма Ю., Фудзивара Ю., Оркин С.Х., Рович Д.Х. (ноябрь 2005 г.). «Спецификация астроцитов с помощью белка bHLH SCL в ограниченной области нервной трубки». Природа . 438 (7066): 360–3. Бибкод : 2005Natur.438..360M . дои : 10.1038/nature04139 . ПМИД 16292311 . S2CID 4425462 .
- ^ Хохстим С, Денин Б, Лукашевич А, Чжоу К, Андерсон DJ (май 2008 г.). «Идентификация позиционно различных подтипов астроцитов, идентичность которых определяется гомеодоменным кодом» . Клетка . 133 (3): 510–22. дои : 10.1016/j.cell.2008.02.046 . ПМК 2394859 . ПМИД 18455991 .
- ^ Чакир Т., Алсан С., Сайбашили Х., Акин А., Ульген КО (декабрь 2007 г.). «Реконструкция и анализ потоков связи между метаболическими путями астроцитов и нейронов: применение к церебральной гипоксии» . Теоретическая биология и медицинское моделирование . 4 (1): 48. дои : 10.1186/1742-4682-4-48 . ПМК 2246127 . ПМИД 18070347 .
- ^ Колб, Брайан и Уишоу, Ян К. (2008) Основы нейропсихологии человека . Стоит издательства. 6-е изд. ISBN 0716795868
- ^ Jump up to: а б Арак А., Парпура В., Санзгири Р.П., Хейдон П.Г. (май 1999 г.). «Трехсторонние синапсы: глия, непризнанный партнер». Тенденции в нейронауках . 22 (5): 208–15. дои : 10.1016/S0166-2236(98)01349-6 . ПМИД 10322493 . S2CID 7067935 .
- ^ Рейнольдс, Гретхен (22 февраля 2012 г.). «Как упражнения питают мозг» . Нью-Йорк Таймс .
- ^ Макдугал Д.Х., Виар Э., Герман Г.Е., Роджерс Р.К. (апрель 2013 г.). «Астроциты заднего мозга обнаруживают глюкопривацию и регулируют перистальтику желудка» . Автономная нейронаука . 175 (1–2): 61–9. дои : 10.1016/j.autneu.2012.12.006 . ПМЦ 3951246 . ПМИД 23313342 .
- ^ Сваминатан Н. (1 октября 2008 г.). «Загадка сканирования мозга раскрыта» . Scientific American Mind : 7. doi : 10.1038/scientificamericanmind1008-16 .
- ^ Фигли Ч.Р., Строман П.В. (февраль 2011 г.). «Роль астроцитов и активности астроцитов в нейрометаболизме, нейрососудистой связи и выработке функциональных сигналов нейровизуализации». Европейский журнал неврологии . 33 (4): 577–88. дои : 10.1111/j.1460-9568.2010.07584.x . ПМИД 21314846 . S2CID 9094771 .
- ^ Сантелло М., Вольтерра А (январь 2009 г.). «Синапсическая модуляция астроцитами посредством Ca2+-зависимого высвобождения глутамата». Нейронаука . Март 158 (1): 253–9. doi : 10.1016/j.neuroscience.2008.03.039 . ПМИД 18455880 . S2CID 9719903 .
- ^ Агулхон С., Фиакко Т.А., Маккарти К.Д. (март 2010 г.). «Кратко- и долгосрочная пластичность гиппокампа не модулируется передачей сигналов Ca2+ астроцитов». Наука . 327 (5970): 1250–4. Бибкод : 2010Sci...327.1250A . дои : 10.1126/science.1184821 . ПМИД 20203048 . S2CID 14594882 .
- ^ Уолц В. (апрель 2000 г.). «Роль астроцитов в выведении избытка внеклеточного калия». Нейрохимия Интернэшнл . 36 (4–5): 291–300. дои : 10.1016/S0197-0186(99)00137-0 . ПМИД 10732996 . S2CID 40064468 .
- ^ Габриэль С., Ньютинг М., Помпер Дж.К., Мершхемке М., Санабриа Э.Р., Эйлерс А. и др. (ноябрь 2004 г.). «Стимул и калий-индуцированная эпилептиформная активность в зубчатой извилине человека у пациентов со склерозом гиппокампа и без него» . Журнал неврологии . 24 (46): 10416–30. doi : 10.1523/JNEUROSCI.2074-04.2004 . ПМК 6730304 . ПМИД 15548657 .
- ^ Пит Р., Варгова Л., Сыкова Э., Пулен Д.А., Олиет Ш.Х. (февраль 2004 г.). «Физиологический вклад астроцитарной среды нейронов в межсинаптические перекрестные помехи» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (7): 2151–5. Бибкод : 2004PNAS..101.2151P . дои : 10.1073/pnas.0308408100 . ПМК 357067 . ПМИД 14766975 .
- ^ Паскуаль О, Каспер К.Б., Кубера С., Чжан Дж., Ревилла-Санчес Р., Сул Дж.Ю. и др. (октябрь 2005 г.). «Астроцитарная пуринергическая сигнализация координирует синаптические сети». Наука . 310 (5745): 113–6. Бибкод : 2005Sci...310..113P . дои : 10.1126/science.1116916 . ПМИД 16210541 . S2CID 36808788 .
- ^ Парри Р., Крунелли В. (январь 2003 г.). «Астроцитарный мост от синапса к кровотоку». Природная неврология . 6 (1): 5–6. дои : 10.1038/nn0103-5 . ПМИД 12494240 . S2CID 42872329 .
- ^ Ишибаши Т., Дакин К.А., Стивенс Б. , Ли П.Р., Козлов С.В., Стюарт К.Л., Филдс Р.Д. (март 2006 г.). «Астроциты способствуют миелинизации в ответ на электрические импульсы» . Нейрон . 49 (6): 823–32. дои : 10.1016/j.neuron.2006.02.006 . ПМЦ 1474838 . ПМИД 16543131 .
- ^ Jump up to: а б Андерсон М.А., Бурда Дж.Э., Рен Ю., Ао Ю., О'Ши Т.М., Кавагути Р. и др. (апрель 2016 г.). «Формирование рубцов астроцитов способствует регенерации аксонов центральной нервной системы» . Природа . 532 (7598): 195–200. Бибкод : 2016Natur.532..195A . дои : 10.1038/nature17623 . ПМЦ 5243141 . ПМИД 27027288 .
- ^ Потокар, Майя; Морита, Мицухиро; Виче, Герхард; Йоргачевски, Джерней (2 июля 2020 г.). «Разнообразие промежуточных филаментов астроцитов» . Клетки . 9 (7): 1604. doi : 10.3390/cells9071604 . ISSN 2073-4409 . ПМК 7408014 . ПМИД 32630739 .
- ^ Лидделоу С.А., Гуттенплан К.А., Кларк Л.Е., Беннетт Ф.К., Болен С.Дж., Ширмер Л. и др. (январь 2017 г.). «Нейротоксические реактивные астроциты индуцируются активированной микроглией» . Природа . 541 (7638): 481–487. Бибкод : 2017Natur.541..481L . дои : 10.1038/nature21029 . ПМК 5404890 . ПМИД 28099414 .
- ^ Хан X, Чен М, Ван Ф, Виндрем М, Ван С, Шанц С и др. (март 2013 г.). «Приживление переднего мозга человеческими глиальными клетками-предшественниками повышает синаптическую пластичность и обучаемость у взрослых мышей» . Клеточная стволовая клетка . 12 (3): 342–53. дои : 10.1016/j.stem.2012.12.015 . ПМЦ 3700554 . ПМИД 23472873 .
- ^ Бранкаччо М., Эдвардс М.Д., Паттон А.П., Смайлли Н.Дж., Чешам Дж.Э., Мэйвуд Э.С., Гастингс М.Х. (январь 2019 г.). «Клеточно-автономные часы астроцитов управляют циркадным поведением млекопитающих» . Наука . 363 (6423): 187–192. Бибкод : 2019Sci...363..187B . дои : 10.1126/science.aat4104 . ПМК 6440650 . ПМИД 30630934 .
- ^ Jump up to: а б с Носсенсон Н., Магал А., Мессер Х. (2016). «Обнаружение стимулов в результате активности нескольких нейронов: эмпирическое исследование и теоретические последствия». Нейрокомпьютинг . 174 : 822–837. дои : 10.1016/j.neucom.2015.10.007 .
- ^ Jump up to: а б Носсенсон Н. (2013). Модельное обнаружение присутствия стимула по нейрофизиологическим сигналам (PDF) . Библиотека точных наук и техники Неймана, Тель-Авивский университет: докторская диссертация, Тель-Авивский университет.
- ^ Корнелл-Белл А.Х., Финкбайнер С.М., Купер М.С., Смит С.Дж. (январь 1990 г.). «Глутамат индуцирует волны кальция в культивируемых астроцитах: передача глиальных сигналов дальнего действия». Наука . 247 (4941): 470–3. Бибкод : 1990Sci...247..470C . дои : 10.1126/science.1967852 . ПМИД 1967852 .
- ^ Джахроми Б.С., Робитайл Р., член парламента от Чарльтона (июнь 1992 г.). «Высвобождение передатчика увеличивает внутриклеточный кальций в перисинаптических шванновских клетках in situ». Нейрон . 8 (6): 1069–77. дои : 10.1016/0896-6273(92)90128-Z . ПМИД 1351731 . S2CID 6855190 .
- ^ Верхрацкий А., Орканд Р.К., Кеттенманн Х. (январь 1998 г.). «Глиальный кальций: гомеостаз и сигнальная функция». Физиологические обзоры . 78 (1): 99–141. дои : 10.1152/physrev.1998.78.1.99 . ПМИД 9457170 . S2CID 823182 .
- ^ Jump up to: а б Эберт У, Кох М (сентябрь 1997 г.). «Акустические вздрагивающие потенциалы в миндалевидном теле крысы: эффект возгорания». Физиология и поведение . 62 (3): 557–62. дои : 10.1016/S0031-9384(97)00018-8 . ПМИД 9272664 . S2CID 41925078 .
- ^ Jump up to: а б Фрот М., Маньен М., Могьер Ф., Гарсиа-Ларреа Л. (март 2007 г.). «Человеческий SII и задняя островковая доля по-разному кодируют тепловые лазерные стимулы» . Кора головного мозга . 17 (3): 610–20. дои : 10.1093/cercor/bhk007 . ПМИД 16614165 .
- ^ Jump up to: а б Перлман, Идо. «Электроретинограмма: ЭРГ Идо Перлмана - Webvision» . webvision.med.utah.edu .
- ^ Jump up to: а б Тиан Г.Ф., Азми Х., Такано Т., Сюй Кью, Пэн В., Лин Дж. и др. (сентябрь 2005 г.). «Астроцитарная основа эпилепсии» . Природная медицина . 11 (9): 973–81. дои : 10.1038/nm1277 . ПМК 1850946 . ПМИД 16116433 .
- ^ Герц Л., Шусбо А., Бехлер Н., Мукерджи С., Федоров С. (февраль 1978 г.). «Кинетические характеристики поглощения глутамата нормальными астроцитами в культурах». Нейрохимические исследования . 3 (1): 1–14. дои : 10.1007/BF00964356 . ПМИД 683409 . S2CID 8626930 .
- ^ Беннетт М.В., Контрерас Дж.Э., Букаускас Ф.Ф., Саес Х.К. (ноябрь 2003 г.). «Новые роли астроцитов: полуканалам щелевых соединений есть что сообщить» . Тенденции в нейронауках . 26 (11): 610–7. дои : 10.1016/j.tins.2003.09.008 . ПМЦ 3694339 . ПМИД 14585601 .
- ^ Ньюман Э.А. (апрель 2001 г.). «Распространение межклеточных кальциевых волн в астроцитах сетчатки и клетках Мюллера» . Журнал неврологии . 21 (7): 2215–23. doi : 10.1523/JNEUROSCI.21-07-02215.2001 . ПМК 2409971 . ПМИД 11264297 .
- ^ Парпура В., Хейдон П.Г. (июль 2000 г.). «Физиологические уровни астроцитарного кальция стимулируют высвобождение глутамата, модулируя соседние нейроны» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (15): 8629–34. Бибкод : 2000PNAS...97.8629P . дои : 10.1073/pnas.97.15.8629 . ПМК 26999 . ПМИД 10900020 .
- ^ Jump up to: а б Лопес-Каамаль Ф., Оярсун Д.А., Миддлтон Р.Х., Гарсия М.Р. (май 2014 г.). «Пространственное количественное определение цитозольного Ca 2+ Накопление в невозбудимых клетках: аналитическое исследование» . IEEE/ACM Transactions on Computational Biology and Bioinformatics . 11 (3): 592–603. doi : 10.1109/TCBB.2014.2316010 . PMID 26356026 .
- ^ Jump up to: а б Хейдон П.Г. (март 2001 г.). «ГЛИА: слушаем синапс и говорим с ним» (PDF) . Обзоры природы. Нейронаука . 2 (3): 185–93. дои : 10.1038/35058528 . ПМИД 11256079 . S2CID 15777434 .
- ^ «Менее известные клетки мозга могут быть ключом к сохранению бодрствования без ущерба для познания и здоровья» . Инсайдер ВГУ | Университет штата Вашингтон . 17 августа 2023 г. Проверено 30 марта 2024 г.
- ^ Jump up to: а б с д Астроцитомы. Архивировано 5 апреля 2012 г. в Wayback Machine . Международная ассоциация радиохирургии (2010).
- ^ Jump up to: а б Астроцитомные опухоли . Американская ассоциация неврологических хирургов (август 2005 г.).
- ^ Баркер А.Дж., Уллиан Э.М. (2008). «Новая роль астроцитов в развитии синаптических цепей» . Коммуникативная и интегративная биология . 1 (2): 207–11. дои : 10.4161/cib.1.2.7284 . ПМК 2686024 . ПМИД 19513261 .
- ^ Гарнизон Си-Джей, Догерти-премьер-министр, Каджандер К.С., Карлтон С.М. (ноябрь 1991 г.). «Окрашивание глиального фибриллярного кислого белка (GFAP) в поясничном отделе спинного мозга увеличивается после травмы, вызванной сужением седалищного нерва». Исследования мозга . 565 (1): 1–7. дои : 10.1016/0006-8993(91)91729-К . ПМИД 1723019 . S2CID 8251884 .
- ^ Вольтерра А., Мелдолеси Дж. (август 2005 г.). «Астроциты, от мозгового клея до элементов связи: революция продолжается». Обзоры природы. Нейронаука . 6 (8): 626–40. дои : 10.1038/nrn1722 . ПМИД 16025096 . S2CID 14457143 .
- ^ Халасса М.М., Феллин Т., Хейдон П.Г. (февраль 2007 г.). «Трехсторонний синапс: роль глиотрансмиссии в здоровье и болезнях». Тенденции молекулярной медицины . 13 (2): 54–63. doi : 10.1016/j.molmed.2006.12.005 . ПМИД 17207662 .
- ^ Софронев М.В. (ноябрь 2014 г.). «Астроглиоз» . Перспективы Колд-Спринг-Харбор в биологии . 7 (2): а020420. doi : 10.1101/cshperspect.a020420 . ПМЦ 4315924 . ПМИД 25380660 .
- ^ Сёльвандер С., Никитиду Э., Бролин Р., Седерберг Л., Селин Д., Ланнфельт Л., Эрландссон А. (май 2016 г.). «Накопление амилоида-β астроцитами приводит к увеличению эндосом и индуцированному микровезикулами апоптозу нейронов» . Молекулярная нейродегенерация . 11 (1): 38. дои : 10.1186/s13024-016-0098-z . ПМК 4865996 . ПМИД 27176225 .
- ^ Бхат Р., Кроу Э.П., Битто А., Мох М., Кацетос К.Д., Гарсия Ф.У. и др. (12 сентября 2012 г.). «Старение астроцитов как компонент болезни Альцгеймера» . ПЛОС ОДИН . 7 (9): e45069. Бибкод : 2012PLoSO...745069B . дои : 10.1371/journal.pone.0045069 . ПМК 3440417 . ПМИД 22984612 .
- ^ Ростами Дж., Холмквист С., Линдстрем В., Сигвардсон Дж., Вестермарк Г.Т., Ингельссон М. и др. (декабрь 2017 г.). «Человеческие астроциты переносят агрегированный альфа-синуклеин через туннельные нанотрубки» . Журнал неврологии . 37 (49): 11835–11853. doi : 10.1523/JNEUROSCI.0983-17.2017 . ПМК 5719970 . ПМИД 29089438 .
- ^ Хан ТУ, Драйна Д (август 2019 г.). «Заикающиеся мутации GNPTAB человека, введенные в организм мышей, вызывают дефицит вокализации и патологию астроцитов в мозолистом теле» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (35): 17515–17524. Бибкод : 2019PNAS..11617515H . дои : 10.1073/pnas.1901480116 . ПМЦ 6717282 . ПМИД 31405983 . S2CID 6717282 .
- ^ Марагакис, штат Нью-Джерси, доктор Ротштейн (декабрь 2006 г.). «Механизмы заболевания: астроциты при нейродегенеративных заболеваниях». Природная клиническая практика. Неврология . 2 (12): 679–89. дои : 10.1038/ncpneuro0355 . ПМИД 17117171 . S2CID 16188129 .
- ^ Рен Х., Хань Р., Чен Х, Лю Х, Ван Дж, Ван Л, Ян Х, Ван Дж (май 2020 г.). «Потенциальные терапевтические мишени для воспаления, связанного с внутримозговым кровоизлиянием: обновленная информация» . J Метаболия кровотока головного мозга . 40 (9): 1752–1768. дои : 10.1177/0271678X20923551 . ПМЦ 7446569 . ПМИД 32423330 .
- ^ Цинь Д., Ван Дж., Ле А., Ван Ти.Дж., Чен Х., Ван Дж. (апрель 2021 г.). «Черепно-мозговая травма: ультраструктурные особенности нейронального ферроптоза, активации и поляризации глиальных клеток, а также разрушения гематоэнцефалического барьера» . Клетки . 10 (5): 1009. doi : 10.3390/cells10051009 . ПМЦ 8146242 . ПМИД 33923370 .
- ^ Кифер, Дональд А.; Христос, Джейкоб Ф. (ноябрь 1976 г.). «Распределение эндогенных клеток, окрашивающих диаминобензидин, в нормальном мозге крысы» . Исследования мозга . 116 (2): 312–316. дои : 10.1016/0006-8993(76)90909-4 . ПМИД 61791 . S2CID 3069004 .
- ^ Брауэр, Джеймс Р.; Штейн, Роберт; Смолл, Лорн; Сиссе, Сориба; Шиппер, Хайман М. (ноябрь 1994 г.). «Состав Гомори-позитивных включений в астроцитах дугообразного ядра гипоталамуса: ГОМОРИ-ПОЗИТИВНЫЕ ВКЛЮЧЕНИЯ В АСТРОЦИТАХ» . Анатомическая запись . 240 (3): 407–415. дои : 10.1002/ar.1092400313 . ПМИД 7825737 . S2CID 20052516 .
- ^ Салливан, Брендан; Робисон, Грегори; Пушкарь, Юлия; Янг, Джон К.; Манайе, Кебретен Ф. (январь 2017 г.). «Накопление меди в астроцитах мозга грызунов: видовое различие» . Журнал микроэлементов в медицине и биологии . 39 : 6–13. дои : 10.1016/j.jtemb.2016.06.011 . ПМК 5141684 . ПМИД 27908425 .
- ^ Шиппер, HM (1993). «Цистеаминовая глиопатия in situ: модель клеточного стресса для биогенеза астроцитарных включений» . Журнал невропатологии и экспериментальной неврологии . 52 (4): 399–410. дои : 10.1097/00005072-199307000-00007 . ПМИД 8394877 . S2CID 42421463 .
- ^ Янг, Джон К.; Бейкер, Джеймс Х.; Мюллер, Томас (март 1996 г.). «Иммунореактивность белка, связывающего жирные кислоты головного мозга, в гомори-положительных астроцитах» . Глия . 16 (3): 218–226. doi : 10.1002/(SICI)1098-1136(199603)16:3<218::AID-GLIA4>3.0.CO;2-Y . ISSN 0894-1491 . ПМИД 8833192 . S2CID 9757285 .
- ^ Шмидт, СП; Коридон, Ти Джей; Педерсен, CB; Бросс, П.; Грегерсен, Н. (июнь 2010 г.). «Неправильное сворачивание короткоцепочечной ацил-КоА-дегидрогеназы приводит к делению митохондрий и окислительному стрессу» . Молекулярная генетика и обмен веществ . 100 (2): 155–162. дои : 10.1016/j.ymgme.2010.03.009 . ПМИД 20371198 .
- ^ Ченг, А (2021). «Влияние белков, связывающих жирные кислоты, на индуцированное альфа-синуклеином повреждение митохондрий при синуклеинопатии» . Биомедицины . 9 (5): 560. doi : 10.3390/biomedicines9050560 . ПМЦ 8156290 . ПМИД 34067791 .
- ^ английский, Кристал; Шеперд, Эндрю; Узор, Ндиди-Эсе; Трин, Ронни; Кавелаарс, Аннемике; Хейнен, Коби Дж. (декабрь 2020 г.). «Астроциты спасают здоровье нейронов после лечения цисплатином посредством переноса в митохондрии» . Acta Neuropathologica Communications . 8 (1): 36. дои : 10.1186/s40478-020-00897-7 . ISSN 2051-5960 . ПМК 7082981 . ПМИД 32197663 .
- ^ Янг, Дж. К.; Маккензи, Джей Си; Бейкер, Дж. Х. (февраль 1990 г.). «Ассоциация железосодержащих астроцитов с дофаминергическими нейронами дугообразного ядра» . Журнал нейробиологических исследований . 25 (2): 204–213. дои : 10.1002/jnr.490250208 . ISSN 0360-4012 . ПМИД 2319629 . S2CID 39851598 .
- ^ Реймонд, Марианна Дж.; Донда, Алена; Лемаршан-Беро, Тереза (1989). «Нейроэндокринные аспекты старения: экспериментальные данные» . Гормональные исследования . 31 (1–2): 32–38. дои : 10.1159/000181083 . ISSN 1423-0046 . ПМИД 2656467 .
- ^ Ясумото, Юки; Миядзаки, Хирофуми; Огата, Масаки; Кагава, Ёситеру; Ямамото, Юи; Ислам, Арифул; Ямада, Тецуя; Катагири, Хидеки; Овада, Юджи (декабрь 2018 г.). «Глиальный белок 7, связывающий жирные кислоты (FABP7), регулирует чувствительность нейронов к лептину в дугообразном ядре гипоталамуса» . Молекулярная нейробиология . 55 (12): 9016–9028. дои : 10.1007/s12035-018-1033-9 . ISSN 0893-7648 . ПМИД 29623545 . S2CID 4632807 .
- ^ Роджерс, Ричард К.; Макдугал, Дэвид Х.; Риттер, Сью; Куоллс-Крикмор, Эмили; Германн, Герлинда Э. (01 июля 2018 г.). «Реакция катехоламинергических нейронов заднего мозга мышей на глюкопривные стимулы зависит от астроцитов» . Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 315 (1): R153–R164. дои : 10.1152/ajpregu.00368.2017 . ISSN 0363-6119 . ПМК 6087883 . ПМИД 29590557 .
- ^ Янг, Джон К.; Маккензи, Джеймс К. (ноябрь 2004 г.). «Иммунореактивность GLUT2 в Гомори-положительных астроцитах гипоталамуса» . Журнал гистохимии и цитохимии . 52 (11): 1519–1524. дои : 10.1369/jhc.4A6375.2004 . ISSN 0022-1554 . ПМЦ 3957823 . ПМИД 15505347 .
- ^ Браун, Дженни М.; Бентсен, Мари А.; Рауш, Дилан М.; Фан, Бао Ань; Вик, Даниэль; Васанвала, Хузайфа; Матсен, Майлз Э.; Ачарья, Нихил; Ричардсон, Николь Э.; Чжао, Синь; Чжай, Пэн (сентябрь 2021 г.). «Роль гипоталамической передачи сигналов MAPK/ERK и центральное действие FGF1 в ремиссии диабета» . iScience . 24 (9): 102944. Бибкод : 2021iSci...24j2944B . дои : 10.1016/j.isci.2021.102944 . ПМЦ 8368994 . ПМИД 34430821 .
- ^ Янг, Дж. К. (2020). «Нейрогенез вносит решающий вклад в невропатологию болезни Альцгеймера» . Журнал отчетов о болезни Альцгеймера . 4 (1): 365–371. дои : 10.3233/ADR-200218 . ПМЦ 7592839 . ПМИД 33163897 .
- ^ Джонсон, Эрик CB; Даммер, Эрик Б.; Дуонг, Дюк М.; Пин, Линъянь; Чжоу, Маотянь; Инь, Люмин; Хиггинботэм, Ленора А.; Гуахардо, Эндрю; Уайт, Варфоломей; Тронкосо, Хуан К.; Тамбисетти, Мадхав (май 2020 г.). «Крупномасштабный протеомный анализ мозга и спинномозговой жидкости при болезни Альцгеймера выявляет ранние изменения в энергетическом обмене, связанные с активацией микроглии и астроцитов» . Природная медицина . 26 (5): 769–780. дои : 10.1038/s41591-020-0815-6 . ISSN 1078-8956 . ПМЦ 7405761 . ПМИД 32284590 .
- ^ Мацумата, Михо; Сакаёри, Нобуюки; Маэкава, Мотоко; Овада, Юджи; Ёсикава, Такео; Осуми, Норико (01 июля 2012 г.). «Влияние Fabp7 и Fabp5 на постнатальный нейрогенез гиппокампа у мышей» . Стволовые клетки . 30 (7): 1532–1543. дои : 10.1002/stem.1124 . ISSN 1066-5099 . ПМИД 22581784 . S2CID 13531289 .
- ^ Дэвис С.Дж., Ши CH, Ноубл М., Майер-Прошель М., Дэвис Дж.Э., Прошель С. (март 2011 г.). Комбс С (ред.). «Трансплантация специфических астроцитов человека способствует функциональному восстановлению после травмы спинного мозга» . ПЛОС ОДИН . 6 (3): e17328. Бибкод : 2011PLoSO...617328D . дои : 10.1371/journal.pone.0017328 . ПМК 3047562 . ПМИД 21407803 .
- ^ Ян Ю, Ге В, Чен Ю, Чжан Цз, Шен В, Ву С и др. (декабрь 2003 г.). «Вклад астроцитов в долгосрочную потенциацию гиппокампа за счет высвобождения D-серина» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (25): 15194–9. Бибкод : 2003PNAS..10015194Y . дои : 10.1073/pnas.2431073100 . ПМК 299953 . ПМИД 14638938 .
- ^ Цзяо Дж.В., Фельдхайм Д.А., Чен Д.Ф. (июнь 2008 г.). «Эфрины как негативные регуляторы нейрогенеза взрослых в различных регионах центральной нервной системы» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (25): 8778–83. Бибкод : 2008PNAS..105.8778J . дои : 10.1073/pnas.0708861105 . ПМЦ 2438395 . ПМИД 18562299 .
- ^ Кренчик Р., Вейк Дж.П., Лю Ю., Чжан З.Дж., Чжан С.К. (май 2011 г.). «Спецификация трансплантируемых подтипов астроглии из плюрипотентных стволовых клеток человека» . Природная биотехнология . 29 (6): 528–34. дои : 10.1038/nbt.1877 . ПМК 3111840 . ПМИД 21602806 . . Краткое содержание: Человеческие астроциты, выращенные из стволовых клеток в лабораторной чашке исследователями Университета Висконсина . sciencedebate.com (22 мая 2011 г.)
- ^ Хан Дж., Кеснер П., Метна-Лоран М., Дуан Т., Сюй Л., Жорж Ф. и др. (март 2012 г.). «Острые каннабиноиды нарушают рабочую память посредством модуляции астроглиальных рецепторов CB1 LTD гиппокампа» . Клетка . 148 (5): 1039–50. дои : 10.1016/j.cell.2012.01.037 . ПМИД 22385967 .
- ^ Беллавер Б., Повала Г., Феррейра П.Л., Феррари-Соуза Ж.П., Леффа Д.Т., Люсье Ф.З. и др. (май 2023 г.). «Реактивность астроцитов влияет на эффекты бета-амилоида на патологию тау при доклинической стадии болезни Альцгеймера» . Природная медицина . 29 (7): 1775–1781. дои : 10.1038/s41591-023-02380-x . ПМЦ 10353939 . ПМИД 37248300 .
- ^ Де Селья, Роберта; Ледонн, Ада; Литвин, Дэвид Грегори; Линд, Барбара Ликке; Каррьеро, Джованни; Латальята, Эмануэле Клаудио; Биндоччи, Эрика; Ди Кастро, Мария Амалия; Савчук, Ярослав; Виталий, Илария; Ранджак, Анураг; Конджиу, Мауро; Дом приходского священника, Тара; Уисден, Уильям; Харрис, Кеннет; Мамели, Мануэль; Меркури, Никола; Телли, Людовик; Вольтерра, Андреа (6 сентября 2023 г.). «Специализированные астроциты опосредуют глутаматергическую глиотрансмиссию в ЦНС» . Природы . 622 (7981): 120–129. Бибкод : 2023Natur.622..120D . doi : 10.1038/s41586-023-06502-w . ПМЦ 10550825 . ПМИД 37674083 .
- ^ Грегори Н., Прёшель С., Нобл М., Майер-Прёшель М. (январь 2002 г.). «Трипотенциальные клетки-предшественники, ограниченные глией (GRP), и развитие глии в спинном мозге: генерация бипотенциальных клеток-предшественников астроцитов олигодендроцитов типа 2 и дорсально-вентральные различия в функции клеток GRP» . Журнал неврологии . 22 (1): 248–56. doi : 10.1523/JNEUROSCI.22-01-00248.2002 . ПМК 6757619 . ПМИД 11756508 .
- ^ Фултон Б.П., Бёрн Дж.Ф., Рафф М.К. (декабрь 1992 г.). «Визуализация клеток-предшественников O-2A в зрительном нерве развивающихся и взрослых крыс путем стимулированного кисквалатом поглощения кобальта» . Журнал неврологии . 12 (12): 4816–33. doi : 10.1523/JNEUROSCI.12-12-04816.1992 . ПМЦ 6575772 . ПМИД 1281496 .
- ^ Левисон С.В., Голдман Дж.Э. (февраль 1993 г.). «И олигодендроциты, и астроциты развиваются из предшественников в субвентрикулярной зоне постнатального переднего мозга крысы». Нейрон . 10 (2): 201–12. дои : 10.1016/0896-6273(93)90311-E . ПМИД 8439409 . S2CID 1428135 .
- ^ Зерлин М., Левисон С.В., Голдман Дж.Э. (ноябрь 1995 г.). «Ранние закономерности миграции, морфогенеза и экспрессии промежуточных филаментов клеток субвентрикулярной зоны в постнатальном переднем мозге крыс» . Журнал неврологии . 15 (11): 7238–49. doi : 10.1523/JNEUROSCI.15-11-07238.1995 . ПМК 6578041 . ПМИД 7472478 .
- ^ Брауэр-младший, Стейн Р., Смолл Л., Сиссе С., Шиппер Х.М. (ноябрь 1994 г.). «Состав Гомори-положительных включений в астроцитах дугообразного ядра гипоталамуса». Анатомическая запись . 240 (3): 407–15. дои : 10.1002/ar.1092400313 . ПМИД 7825737 . S2CID 20052516 .
- ^ Янг Дж. К., Маккензи Дж. С. (ноябрь 2004 г.). «Иммунореактивность GLUT2 в Гомори-положительных астроцитах гипоталамуса» . Журнал гистохимии и цитохимии . 52 (11): 1519–24. дои : 10.1369/jhc.4A6375.2004 . ПМЦ 3957823 . ПМИД 15505347 .
- ^ Марти Н., Даллапорта М., Форец М., Эмери М., Таруссио Д., Бади И. и др. (декабрь 2005 г.). «Регуляция секреции глюкагона с помощью транспортера глюкозы 2 типа (glut2) и астроцит-зависимых сенсоров глюкозы» . Журнал клинических исследований . 115 (12): 3545–53. дои : 10.1172/jci26309 . ПМЦ 1297256 . ПМИД 16322792 .
- ^ Чой Б.Х., Лэпэм Л.В. (июнь 1978 г.). «Радиальная глия в головном мозге плода человека: комбинированное исследование по аппарату Гольджи, иммунофлуоресцентное и электронно-микроскопическое исследование». Исследования мозга . 148 (2): 295–311. дои : 10.1016/0006-8993(78)90721-7 . ПМИД 77708 . S2CID 3058148 .
- ^ Шмечель Д.Е., Ракич П. (июнь 1979 г.). «Исследование методом Гольджи радиальных глиальных клеток в развивающемся конечном мозге обезьян: морфогенез и трансформация в астроциты». Анатомия и эмбриология . 156 (2): 115–52. дои : 10.1007/BF00300010 . ПМИД 111580 . S2CID 40494903 .
- ^ Миссон Дж.П., Эдвардс М.А., Ямамото М., Кэвинесс В.С. (ноябрь 1988 г.). «Идентификация радиальных глиальных клеток в развивающейся центральной нервной системе мышей: исследования, основанные на новом иммуногистохимическом маркере». Исследования мозга. Исследования развития мозга . 44 (1): 95–108. дои : 10.1016/0165-3806(88)90121-6 . ПМИД 3069243 .
- ^ Фойгт Т. (ноябрь 1989 г.). «Развитие глиальных клеток в стенке головного мозга хорьков: прямое прослеживание их трансформации из радиальной глии в астроциты». Журнал сравнительной неврологии . 289 (1): 74–88. дои : 10.1002/cne.902890106 . ПМИД 2808761 . S2CID 24449457 .
- ^ Голдман С.А., Зухар А., Барами К., Микава Т., Недзвецки Д. (август 1996 г.). «Клетки эпендимальной / субэпендимальной зоны мозга постнатальных и взрослых певчих птиц генерируют как нейроны, так и ненейрональные братья и сестры in vitro и in vivo». Журнал нейробиологии . 30 (4): 505–20. doi : 10.1002/(SICI)1097-4695(199608)30:4<505::AID-NEU6>3.0.CO;2-7 . ПМИД 8844514 .
- ^ Сиснерос И.Е., Горпаде А. (октябрь 2014 г.). «Метамфетамин и нейротоксичность, вызванная ВИЧ-1: роль передачи сигналов цАМФ, связанного с следами амина, в астроцитах» . Нейрофармакология . 85 : 499–507. doi : 10.1016/j.neuropharm.2014.06.011 . ПМК 4315503 . ПМИД 24950453 .
Более того, сверхэкспрессия TAAR1 значительно снижала уровни EAAT-2 и клиренс глутамата, которые еще больше снижались под действием METH. В совокупности наши данные показывают, что обработка METH активировала TAAR1, что приводило к образованию внутриклеточного цАМФ в астроцитах человека и модулировало способность к выведению глутамата. Более того, молекулярные изменения уровней TAAR1 астроцитов соответствуют изменениям уровней и функции EAAT-2 астроцитов.
Дальнейшее чтение [ править ]
- Уайт Ф.А., Юнг Х., Миллер Р.Дж. (декабрь 2007 г.). «Хемокины и патофизиология нейропатической боли» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (51): 20151–8. Бибкод : 2007PNAS..10420151W . дои : 10.1073/pnas.0709250104 . ПМК 2154400 . ПМИД 18083844 .
- Миллиган Э.Д., Уоткинс Л.Р. (январь 2009 г.). «Патологическая и защитная роль глии при хронической боли» . Обзоры природы. Нейронаука . 10 (1): 23–36. дои : 10.1038/nrn2533 . ПМЦ 2752436 . ПМИД 19096368 .
- Уоткинс Л.Р., Миллиган Э.Д., Майер С.Ф. (август 2001 г.). «Глиальная активация: движущая сила патологической боли». Тенденции в нейронауках . 24 (8): 450–5. дои : 10.1016/S0166-2236(00)01854-3 . ПМИД 11476884 . S2CID 6822068 .
- Фриман М.Р. (ноябрь 2010 г.). «Спецификация и морфогенез астроцитов» . Наука . 330 (6005): 774–8. Бибкод : 2010Sci...330..774F . дои : 10.1126/science.1190928 . ПМК 5201129 . ПМИД 21051628 .
- Верхрацкий А.; Батт, AM (2013). «Цифры: сколько глиальных клеток в мозге?». Глиальная физиология и патофизиология . Джон Уайли и сыновья. стр. 93–96. ISBN 978-0-470-97853-5 .
- Рен Х., Хань Р., Чен Х, Лю Х, Ван Дж, Ван Л, Ян Х, Ван Дж (май 2020 г.). «Потенциальные терапевтические мишени для воспаления, связанного с внутримозговым кровоизлиянием: обновленная информация» . J Метаболия кровотока головного мозга . 40 (9): 1752–1768. дои : 10.1177/0271678X20923551 . ПМЦ 7446569 . ПМИД 32423330 .
Внешние ссылки [ править ]
- Клеточная база данных – астроциты
- Гистология UIUC, предмет 57
- «Астроциты». Архивировано 21 ноября 2012 г. в Wayback Machine Общества нейробиологии.
- Департамент нейробиологии Викиверситета
- Поиск NIF – астроциты через информационную структуру нейронауки