Дендритный шип
| Дендритный шип | |
|---|---|
 Шипастый дендрит стриарного среднего шиповатого нейрона. | |
 Распространенные типы дендритных шипиков. | |
| Подробности | |
| Идентификаторы | |
| латинский | геммулы дендритской |
| МеШ | D049229 |
| ТД | Х2.00.06.1.00036 |
| Анатомические термины микроанатомии | |
Дендритный шип (или шип нейрона ) представляет собой небольшой мембранозный выступ из дендрита , который обычно получает входные данные от одного аксона в синапсе . Дендритные шипики служат местом хранения синаптической силы и помогают передавать электрические сигналы телу клетки нейрона. Большинство шипов имеют выпуклую головку (головку шипа) и тонкую шейку, которая соединяет головку шипа со стержнем дендрита. Дендриты одного нейрона могут содержать от сотен до тысяч шипов. Помимо шипов, обеспечивающих анатомический субстрат для хранения памяти и синаптической передачи, они также могут служить для увеличения количества возможных контактов между нейронами. [1] Было также высказано предположение, что изменения активности нейронов положительно влияют на морфологию позвоночника. [2]
Структура [ править ]
Дендритные шипы небольшие, объем головки шипов составляет 0,01 мкм. 3 до 0,8 мкм 3 . Шипы с сильными синаптическими контактами обычно имеют большую головку шипа, которая соединяется с дендритом через перепончатую шейку. Наиболее заметными классами формы позвоночника являются «тонкий», «короткий», «грибовидный» и «раздвоенный». Исследования электронной микроскопии показали, что между этими категориями существует континуум форм. [3] Считается, что переменная форма и объем позвоночника коррелируют с силой и зрелостью каждого синапса позвоночника.
Распространение [ править ]
Дендритные шипы обычно получают возбуждающую информацию располагаются как тормозящие, так и возбуждающие соединения от аксонов, хотя иногда на одной и той же головке шипа . [4] Близость возбуждающего аксона к дендритным отросткам недостаточна для прогнозирования наличия синапса, как продемонстрировала лаборатория Лихтмана в 2015 году. [5]
расположены на дендритах большинства основных нейронов головного мозга, включая нейроны неокортекса Шипы , средние шипистые нейроны и полосатого тела клетки Пуркинье мозжечка пирамидные . Дендритные шипы встречаются при плотности до 5 шипов на 1 мкм участка дендрита. Гиппокампальные и кортикальные пирамидные нейроны могут получать десятки тысяч преимущественно возбуждающих импульсов от других нейронов на свои столь же многочисленные шипики, тогда как число шипов на дендритах нейронов Пуркинье на порядок больше.
Цитоскелет и органеллы [ править ]
Цитоскелет дендритных шипиков особенно важен с точки зрения их синаптической пластичности ; без динамического цитоскелета шипы были бы неспособны быстро менять свои объемы и форму в ответ на раздражители. Эти изменения формы могут повлиять на электрические свойства позвоночника. Цитоскелет дендритных шипиков в основном состоит из нитчатого актина ( F-актина ). Тубулин Присутствуют мономеры и белки, ассоциированные с микротрубочками (MAP), а также организованные микротрубочки . [6] Поскольку цитоскелет шипов состоит в основном из актина, это позволяет им иметь очень динамичную форму и размер. Актиновый цитоскелет непосредственно определяет морфологию позвоночника, а регуляторы актина, небольшие GTPases , такие как Rac , RhoA и CDC42 , быстро модифицируют этот цитоскелет. Сверхактивный Rac1 приводит к постоянному уменьшению дендритных шипов.
В дополнение к своей электрофизиологической активности и активности, опосредованной рецепторами, шипики, по-видимому, обладают везикулярной активностью и могут даже транслировать белки . сложенные друг на друга диски гладкой эндоплазматической сети В дендритных шипиках обнаружены (SER). Формирование этого « позвоночного аппарата » зависит от белка синаптоподина , который, как полагают, играет важную роль в переработке кальция. «гладкие» везикулы В шипиках также были обнаружены , поддерживающие везикулярную активность в дендритных шипиках. Присутствие полирибосом в шипиках также предполагает трансляционную активность белка в самом шипике, а не только в дендритах.
Морфогенез [ править ]
Морфогенез дендритных шипиков имеет решающее значение для индукции долгосрочной потенциации (LTP). [7] [8] Морфология шипика зависит от состояния актина : в глобулярной (G-актин) или нитевидной (F-актин) формах. Роль ГТФаз семейства Rho и их влияние на стабильность актина и подвижность позвоночника. [9] имеет важное значение для памяти. Если дендритный шип является основной единицей хранения информации, то способность шипа спонтанно расширяться и втягиваться должна быть ограничена. В противном случае информация может быть потеряна. Семейство ГТФаз Rho вносит значительный вклад в процесс, стимулирующий полимеризацию актина, что, в свою очередь, увеличивает размер и форму шипа. [10] Большие шипы более стабильны, чем более мелкие, и могут быть устойчивы к модификации дополнительной синаптической активностью. [11] Поскольку изменения формы и размера дендритных шипиков коррелируют с силой возбуждающих синаптических связей и в значительной степени зависят от ремоделирования лежащего в их основе актинового цитоскелета, [12] специфические механизмы регуляции актина и, следовательно, семейство GTPases Rho являются неотъемлемой частью формирования, созревания и пластичности дендритных шипиков, а также обучения и памяти.
Путь RhoA [ править ]
Одной из основных Rho GTPases, участвующих в морфогенезе шипов, является RhoA , белок, который также модулирует регуляцию и время клеточного деления. В контексте активности в нейронах RhoA активируется следующим образом: как только кальций проникает в клетку через NMDA-рецепторы , он связывается с кальмодулином и активирует CaMKII , что приводит к активации RhoA. [10] Активация белка RhoA активирует ROCK, киназу RhoA, что приводит к стимуляции киназы LIM , которая, в свою очередь, ингибирует белок кофилин . Функция кофилина заключается в реорганизации актинового цитоскелета клетки; а именно, он деполимеризует сегменты актина и, таким образом, ингибирует рост конусов роста и восстановление аксонов. [13]
Исследование, проведенное Муракоши и др. в 2011 году вовлекли Rho GTPases RhoA и Cdc42 в морфогенез дендритных шипов. Обе ГТФазы быстро активировались в одиночных дендритных шипах пирамидных нейронов в области СА1 гиппокампа крысы во время структурной пластичности, вызванной долговременными стимулами потенциации. Одновременная активация RhoA и Cdc42 привела к временному увеличению роста позвоночника до 300% в течение пяти минут, которое затем сменилось меньшим, но устойчивым ростом в течение тридцати минут. [10] Активация RhoA распространилась вокруг подвергающегося стимуляции позвоночника, и было установлено, что RhoA необходим для переходной фазы и, скорее всего, для устойчивой фазы, а также для роста позвоночника.
Путь Cdc42 [ править ]
Cdc42 участвует во многих различных функциях, включая рост дендритов, ветвление и стабильность ветвей. [14] Приток кальция в клетку через NMDA-рецепторы связывается с кальмодулином и активирует Ca2+/кальмодулин-зависимые протеинкиназы II (CaMKII). В свою очередь, CaMKII активируется, и это активирует Cdc42, после чего не происходит передачи сигналов обратной связи выше кальция и CaMKII. Если пометить зеленым флуоресцентным белком, усиленным мономером, можно увидеть, что активация Cdc42 ограничивается только стимулированным шипом дендрита. Это связано с тем, что молекула постоянно активируется во время пластичности и сразу же инактивируется после диффузии из позвоночника. Несмотря на свою разрозненную активность, Cdc42 все еще может перемещаться за пределы стимулированного позвоночника, как и RhoA. Cdc42 активирует PAK, протеинкиназу, которая специфически фосфорилирует и, следовательно, инактивирует ADF/кофилин. [15] Инактивация кофилина приводит к усилению полимеризации актина и увеличению объема шипика. Для поддержания этого увеличения объема позвоночника необходима активация Cdc42.
структурной изменения Наблюдаемые пластичности
Муракоши, Ван и Ясуда (2011) исследовали влияние активации Rho GTPase на структурную пластичность одиночных дендритных шипиков, выяснив различия между переходной и устойчивой фазами. [10]
структурной Временные изменения пластичности
Применение низкочастотной последовательности двухфотонного разблокирования глутамата в одном дендритном шипе может вызвать быструю активацию как RhoA, так и Cdc42. В течение следующих двух минут объем стимулируемого позвоночника может увеличиться до 300 процентов от первоначального размера. Однако это изменение морфологии позвоночника носит временный характер; объем позвоночника уменьшается через пять минут. Введение трансферазы C3, ингибитора Rho, или глицил-H1152, ингибитора Rock, ингибирует временное расширение позвоночника, что указывает на то, что для этого процесса каким-то образом необходима активация пути Rho-Rock. [10]
структурной изменения Устойчивые пластичности
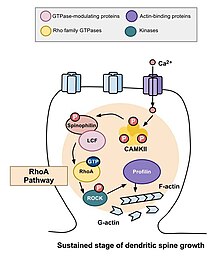
После того, как происходят описанные выше временные изменения, объем позвоночника уменьшается до тех пор, пока он не увеличится на 70–80 процентов от первоначального объема. Это устойчивое изменение структурной пластичности продлится около тридцати минут. И снова введение трансферазы C3 и Glycyl-H1152 подавляло этот рост, что позволяет предположить, что путь Rho-Rock необходим для более стойкого увеличения объема позвоночника. Кроме того, введение Cdc42-связывающего домена Wasp или ингибитора, нацеленного на активацию Pak1-3 (IPA3), уменьшает этот устойчивый рост объема, демонстрируя, что путь Cdc42-Pak также необходим для этого роста объема позвоночника. Это важно, поскольку устойчивые изменения структурной пластичности могут обеспечить механизм кодирования, поддержания и восстановления воспоминаний. Сделанные наблюдения могут свидетельствовать о том, что Rho GTPases необходимы для этих процессов. [10]
Физиология [ править ]
Активность рецепторов [ править ]
Дендритные шипики экспрессируют на своей поверхности рецепторы глутамата (например, рецептор AMPA и рецептор NMDA ). Рецептор TrkB для BDNF также экспрессируется на поверхности позвоночника и, как полагают, играет роль в выживании позвоночника. Кончик позвоночника содержит электронно-плотную область, называемую « постсинаптической плотностью » (PSD). PSD непосредственно примыкает к активной зоне синаптического аксона и занимает ~ 10% площади поверхности мембраны позвоночника; нейротрансмиттеры, высвобождаемые из активной зоны, связывают рецепторы постсинаптической плотности позвоночника. Половина синапсов аксонов и дендритных шипов физически связана кальций -зависимым кадгерином , который образует межклеточные адгезионные соединения между двумя нейронами.
Глутаматные рецепторы (GluR) локализованы в постсинаптической плотности и прикреплены элементами цитоскелета к мембране. Они расположены непосредственно над их сигнальным аппаратом, который обычно привязан к нижней стороне плазматической мембраны, что позволяет сигналам, передаваемым GluR в цитозоль, распространяться дальше соседними сигнальными элементами для активации каскадов сигнальной трансдукции . Локализация сигнальных элементов на их GluRs особенно важна для обеспечения активации сигнального каскада, поскольку GluRs не могут влиять на определенные нижестоящие эффекты без близлежащих сигналистов.
Передача сигналов от GluRs опосредована наличием большого количества белков, особенно киназ, локализованных в постсинаптической плотности. К ним относятся кальций -зависимый кальмодулин , CaMKII (кальмодулин-зависимая протеинкиназа II), PKC (протеинкиназа C), PKA (протеинкиназа A), протеинфосфатаза-1 (PP-1) и Fyn-тирозинкиназа . Некоторые сигнальные агенты, такие как CaMKII, активируются в ответ на активность.
Шипы особенно полезны для нейронов, поскольку они разделяют биохимические сигналы. Это может помочь закодировать изменения в состоянии отдельного синапса, не обязательно затрагивая состояние других синапсов того же нейрона. Длина и ширина шейки позвоночника оказывает большое влияние на степень компартментализации, при этом тонкие шипы являются наиболее биохимически изолированными шипами.
Пластичность [ править ]
Дендритные шипики очень «пластичны», то есть шипики существенно изменяются по форме, объему и количеству за небольшие промежутки времени. Поскольку шипики имеют преимущественно актиновый цитоскелет , они динамичны, и большинство шипиков меняют свою форму в течение секунд или минут из-за динамичности ремоделирования актина . Более того, количество шипов очень изменчиво, шипы приходят и исчезают; в считанные часы на пирамидальных клетках коры головного мозга спонтанно появляются или исчезают 10-20% шипиков, хотя более крупные шипики «грибовидной» формы являются наиболее стабильными.
Сохранность и пластичность позвоночника зависят от активности. [16] и независимо от деятельности. BDNF частично определяет уровни позвоночника, [17] низкий уровень активности рецепторов AMPA необходим для поддержания выживания позвоночника, а синапсическая активность с участием рецепторов NMDA способствует росту позвоночника. Кроме того, двухфотонная лазерная сканирующая микроскопия и конфокальная микроскопия показали, что объем позвоночника меняется в зависимости от типов стимулов, подаваемых в синапс.
для обучения памяти Важность и
Доказательства важности [ править ]
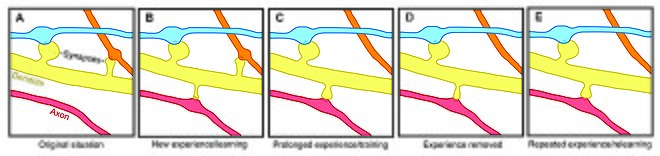
Пластичность позвоночника влияет на мотивацию , обучение и память . [18] [19] [20] В частности, долговременная память частично опосредована ростом новых дендритных шипиков (или увеличением ранее существовавших шипиков) для усиления определенного нервного пути. Поскольку дендритные шипы представляют собой пластичные структуры, на продолжительность жизни которых влияет входная активность, [21] Динамика позвоночника может играть важную роль в поддержании памяти на протяжении всей жизни.
Возрастные изменения скорости вращения позвоночника позволяют предположить, что стабильность позвоночника влияет на обучение. В молодости обновление дендритных шипов относительно велико и приводит к чистой потере шипов. [1] [22] [23] Такая высокая скорость обновления позвоночника может характеризовать критические периоды развития и отражать способность к обучению в подростковом возрасте — разные области коры демонстрируют разные уровни синаптического обмена во время развития, что, возможно, отражает различные критические периоды для конкретных областей мозга. [19] [22] Однако во взрослом возрасте большинство шипов остаются постоянными, а период полураспада шипов увеличивается. [1] Эта стабилизация происходит из-за регулируемого развитием замедления выведения позвоночника - процесса, который может лежать в основе стабилизации воспоминаний в зрелом возрасте. [1] [22]
Вызванные опытом изменения в стабильности дендритных позвонков также указывают на оборот позвоночника как на механизм, участвующий в поддержании долговременной памяти, хотя неясно, как сенсорный опыт влияет на нервные схемы. Две общие модели могут описать влияние опыта на структурную пластичность. С одной стороны, опыт и активность могут стимулировать дискретное формирование соответствующих синаптических связей, которые хранят значимую информацию, необходимую для обучения. С другой стороны, синаптические связи могут образовываться в избытке, а опыт и активность могут привести к обрезанию посторонних синаптических связей. [1]
У лабораторных животных всех возрастов обогащение окружающей среды было связано с ветвлением дендритов, плотностью шипов и общим количеством синапсов. [1] Кроме того, было доказано, что тренировка навыков приводит к формированию и стабилизации новых позвоночника, одновременно дестабилизируя старые. [18] [24] предполагая, что освоение нового навыка включает в себя процесс перестройки нейронных цепей. Поскольку степень ремоделирования позвоночника коррелирует с успехом обучения, это предполагает решающую роль синаптической структурной пластичности в формировании памяти. [24] Кроме того, изменения в стабильности и укреплении позвоночника происходят быстро и наблюдаются в течение нескольких часов после тренировки. [18] [19]
И наоборот, хотя обогащение и тренировка связаны с улучшением формирования и стабильности позвоночника, длительная сенсорная депривация приводит к увеличению скорости удаления позвоночника. [1] [22] и, следовательно, влияет на долгосрочные нейронные цепи. При восстановлении сенсорного опыта после депривации в подростковом возрасте удаление шипов ускоряется, что позволяет предположить, что опыт играет важную роль в чистой потере шипов во время развития. [22] Кроме того, было показано, что другие парадигмы сенсорной депривации, такие как обрезка усов, повышают стабильность новых шипов. [25]
Исследования неврологических заболеваний и травм пролили дополнительный свет на природу и важность поворота позвоночника. После инсульта вблизи места травмы происходит заметное увеличение структурной пластичности, а также наблюдается пяти- или восьмикратное увеличение оборота позвоночника по сравнению с контролем. [26] Дендриты быстро распадаются и собираются заново во время ишемии — как и при инсульте, у выживших наблюдалось увеличение оборота дендритов. [27] Хотя чистая потеря шипов наблюдается при болезни Альцгеймера и случаях умственной отсталости , употребление кокаина и амфетамина связано с увеличением ветвления дендритов и плотности шипов в префронтальной коре и прилежащем ядре . [28] Поскольку значительные изменения плотности позвоночника происходят при различных заболеваниях головного и спинного мозга, это предполагает сбалансированное состояние динамики позвоночника в нормальных условиях, которое может быть подвержено неравновесию при различных патологических состояниях. [28] [29]
Есть также некоторые свидетельства потери дендритных шипов в результате старения. Одно исследование на мышах отметило корреляцию между возрастным уменьшением плотности позвоночника в гиппокампе и возрастным снижением обучения и памяти в гиппокампе. [30] Новые данные также показали дендритные аномалии позвоночника в областях обработки боли ноцицептивной системы спинного мозга, включая поверхностные и промежуточные зоны дорсального рога. [31] [29] [32] [33]
В целом, данные свидетельствуют о том, что дендритные отростки имеют решающее значение для нормального функционирования головного и спинного мозга. Изменения в морфологии позвоночника могут не только влиять на синаптическую пластичность и обработку информации, но также играть ключевую роль во многих неврологических заболеваниях. Более того, даже незначительные изменения плотности или размеров дендритных отростков могут повлиять на свойства нейронной сети. [34] Это может привести к когнитивным изменениям или изменениям настроения, нарушениям обучения и памяти, а также к повышенной чувствительности к боли. [29] Более того, результаты показывают, что поддержание здоровья позвоночника с помощью таких методов лечения, как физические упражнения, когнитивная стимуляция и изменение образа жизни, может быть полезным для сохранения пластичности нейронов и улучшения неврологических симптомов.
Важность оспаривается
Несмотря на экспериментальные данные, которые предполагают роль динамики дендритных позвонков в обеспечении обучения и памяти, степень важности структурной пластичности остается спорной. Например, исследования показывают, что лишь небольшая часть шипов, сформированных во время тренировок, действительно способствует обучению на протяжении всей жизни. [24] Кроме того, формирование новых шипов может не оказывать существенного влияния на связность мозга, а формирование шипов может не оказывать такого большого влияния на сохранение памяти, как другие свойства структурной пластичности, такие как увеличение размера головок позвоночника. [35]
Моделирование [ править ]
Теоретики на протяжении десятилетий выдвигали гипотезы о потенциальной электрической функции шипов, однако наша неспособность изучить их электрические свойства до недавнего времени не позволяла теоретическим работам продвинуться слишком далеко. Недавние достижения в методах визуализации, а также более широкое использование двухфотонного извлечения глутамата привели к множеству новых открытий; теперь мы подозреваем, что существуют потенциалзависимые натрии, [36] калий, [37] и кальций [38] каналы в головках позвоночника. [39]
Теория кабеля обеспечивает теоретическую основу для самого «простого» метода моделирования течения электрических токов по пассивным нервным волокнам. Каждый позвоночник можно рассматривать как два отдела: один представляет собой шею, другой — голову позвоночника. Отделение, представляющее собой только голову позвоночника, должно нести активные свойства.
Ринзеля континуума Модель Бэра и
Чтобы облегчить анализ взаимодействия между множеством шипов, Баер и Ринцель сформулировали новую теорию кабеля, в которой распределение шипов рассматривается как континуум. [40] В этом представлении напряжение головки позвоночника представляет собой локальное пространственное среднее мембранного потенциала в соседних шипах. В формулировке сохраняется то обстоятельство, что между соседними шипами нет прямой электрической связи; Распространение напряжения вдоль дендритов — единственный способ взаимодействия шипов.
Модель спайк-диффузно-спайк [ править ]
Модель SDS задумывалась как упрощенная в вычислительном отношении версия полной модели Бэра и Ринзеля. [41] Он был разработан так, чтобы быть аналитически доступным и иметь как можно меньше свободных параметров, сохраняя при этом наиболее важные, такие как сопротивление шеи и позвоночника. Модель отказывается от приближения континуума и вместо этого использует пассивный дендрит, соединенный с возбудимыми шипами в дискретных точках. Динамика мембран в шипах моделируется с использованием процессов интеграции и огня. Пиковые события моделируются дискретным образом, при этом форма волны обычно представляется в виде прямоугольной функции.
кальция в Моделирование переходных процессов позвоночнике
Переходные процессы кальция в шипиках являются ключевым триггером синаптической пластичности. [42] NMDA-рецепторы , обладающие высокой проницаемостью для кальция, проводят ионы только в том случае, если мембранный потенциал достаточно деполяризован. Таким образом, количество кальция, поступающего в позвоночник во время синаптической активности, зависит от деполяризации головки позвоночника. Данные экспериментов по визуализации кальция ( двухфотонная микроскопия ) и компартментального моделирования показывают, что шипы с шейками с высоким сопротивлением испытывают более сильные переходные процессы кальция во время синаптической активности. [39] [43]
Развитие [ править ]
Дендритные шипы могут развиваться непосредственно из дендритных стержней или из дендритных филоподий . [44] Во время синаптогенеза дендриты быстро прорастают и втягивают филоподии, небольшие мембранные органеллы, лишенные мембранных выступов. Недавно было обнаружено, что белок I-BAR MIM способствует процессу инициации. [45] В течение первой недели рождения в мозге преобладают филоподии, на которых со временем образуются синапсы. Однако по прошествии этой первой недели филоподии заменяются колючими дендритами, а также небольшими короткими шипами, выступающими из колючих дендритов. При развитии определенных филоподий в шипики филоподии рекрутируют пресинаптический контакт с дендритом, что стимулирует образование шипиков для обеспечения специализированного постсинаптического контакта с пресинаптическими выступами.
Однако шипы требуют созревания после формирования. Незрелые шипы имеют нарушенные сигнальные возможности и обычно не имеют «голов» (или имеют очень маленькие головы), а только шеи, в то время как зрелые шипы сохраняют и голову, и шею.
Клиническое значение
Новые исследования указывают на аномалии плотности позвоночника при тревожных расстройствах. [4]
Когнитивные расстройства, такие как СДВГ , болезнь Альцгеймера , аутизм , умственная отсталость и синдром хрупкой Х-хромосомы , могут быть результатом аномалий дендритных шипов, особенно количества шипов и их зрелости. [46] [47] Соотношение зрелых и незрелых шипиков важно для их передачи сигналов, поскольку у незрелых шипиков нарушена синаптическая передача сигналов. Синдром ломкой Х-хромосомы характеризуется избытком незрелых шипов с множественными филоподиями в кортикальных дендритах.
История [ править ]
Дендритные шипики были впервые описаны в конце XIX века Сантьяго Рамоном-и-Кахалем на нейронах мозжечка. [48] Затем Рамон-и-Кахаль предположил, что дендритные шипы могут служить местами контакта между нейронами. Это было продемонстрировано более 50 лет спустя благодаря появлению электронной микроскопии. [49] До появления конфокальной микроскопии живых тканей считалось, что шипы формируются во время эмбрионального развития и затем остаются стабильными после рождения. В этой парадигме вариации синаптического веса считались достаточными для объяснения процессов памяти на клеточном уровне. Но примерно десять лет назад новые методы конфокальной микроскопии продемонстрировали, что дендритные отростки действительно являются подвижными и динамическими структурами, которые подвергаются постоянному обновлению даже после рождения. [50] [51] [44]
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с д и ж г Альварес В.А., Сабатини Б.Л. (2007). «Анатомо-физиологическая пластичность дендритных шипиков». Ежегодный обзор неврологии . 30 : 79–97. дои : 10.1146/annurev.neuro.30.051606.094222 . ПМИД 17280523 .
- ^ Такенберг С., Гори А., Брандт Р. (июнь 2009 г.). «Тонкий, короткий или грибовидный: патология позвоночника при болезни Альцгеймера». Текущие исследования болезни Альцгеймера . 6 (3): 261–8. дои : 10.2174/156720509788486554 . ПМИД 19519307 .
- ^ Офер Н., Бергер Д.Р., Кастури Н., Лихтман Дж.В., Юсте Р. (июль 2021 г.). «Ультраструктурный анализ дендритных шеек позвоночника выявляет континуум морфологии позвоночника» . Развивающая нейробиология . 81 (5): 746–757. дои : 10.1002/dneu.22829 . ПМЦ 8852350 . ПМИД 33977655 . S2CID 234472935 .
- ^ Jump up to: Перейти обратно: а б Эврард М.Р., Ли М., Шен Х., Смит С.С. (октябрь 2021 г.). «Предотвращение синаптической обрезки у подростков в прелимбической коре головного мозга мышей посредством локального нокдауна α4βδ рецепторов ГАМК А увеличивает реакцию тревоги во взрослом возрасте» . Научные отчеты . 11 (1): 21059. Бибкод : 2021NatSR..1121059E . дои : 10.1038/s41598-021-99965-8 . ПМЦ 8548505 . ПМИД 34702942 .
- ^ Кастури Н., Хейворт К.Дж., Бергер Д.Р., Шалек Р.Л., Кончелло Дж.А., Ноулз-Барли С. и др. (июль 2015 г.). «Насыщенная реконструкция объема неокортекса» . Клетка . 162 (3): 648–661. дои : 10.1016/j.cell.2015.06.054 . ПМИД 26232230 .
- ^ Капитеин Л.К., Шлагер М.А., Куйперс М., Вульф П.С., ван Спронсен М., Макинтош Ф.К., Хугенраад CC (февраль 2010 г.). «Смешанные микротрубочки направляют управляемый динеином транспорт грузов в дендриты» . Современная биология . 20 (4): 290–9. дои : 10.1016/j.cub.2009.12.052 . ПМИД 20137950 . S2CID 12180359 .
- ^ Ким Ч., Лисман Дж. Э. (июнь 1999 г.). «Роль актиновой нити в синаптической передаче и долгосрочной потенциации» . Журнал неврологии . 19 (11): 4314–4324. doi : 10.1523/JNEUROSCI.19-11-04314.1999 . ПМК 6782630 . ПМИД 10341235 .
- ^ Крукер Т., Сиггинс Г.Р., Халпейн С. (июнь 2000 г.). «Динамические актиновые филаменты необходимы для стабильной долгосрочной потенциации (LTP) в области CA1 гиппокампа» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (12): 6856–6861. Бибкод : 2000PNAS...97.6856K . дои : 10.1073/pnas.100139797 . ЧВК 18765 . ПМИД 10823894 .
- ^ Таширо А., Юсте Р. (июль 2004 г.). «Регуляция подвижности и стабильности дендритных позвонков с помощью киназы Rac1 и Rho: данные о двух формах подвижности позвоночника». Молекулярная и клеточная нейронауки . 26 (3): 429–440. дои : 10.1016/j.mcn.2004.04.001 . ПМИД 15234347 . S2CID 21100601 .
- ^ Jump up to: Перейти обратно: а б с д и ж Муракоши Х., Ван Х., Ясуда Р. (апрель 2011 г.). «Локальная, стойкая активация Rho GTPases во время пластичности одиночных дендритных шипов» . Природа . 472 (7341): 100–104. Бибкод : 2011Natur.472..100M . дои : 10.1038/nature09823 . ПМК 3105377 . ПМИД 21423166 .
- ^ Касаи Х., Мацузаки М., Ногучи Дж., Ясумацу Н., Накахара Х. (июль 2003 г.). «Взаимоотношения структура-стабильность-функция дендритных шипов». Тенденции в нейронауках . 26 (7): 360–368. дои : 10.1016/S0166-2236(03)00162-0 . ПМИД 12850432 . S2CID 18436944 .
- ^ Хотулайнен П., Хугенраад CC (май 2010 г.). «Актин в дендритных шипиках: соединение динамики с функцией» . Журнал клеточной биологии . 189 (4): 619–629. дои : 10.1083/jcb.201003008 . ПМЦ 2872912 . ПМИД 20457765 .
- ^ Кисс С., Ли Дж., Селес А., Гизатуллин Р.З., Кашуба В.И., Лушникова Т. и др. (1 января 1997 г.). «Присвоение генов ARHA и GPX1 полосам хромосом человека 3p21.3 путем гибридизации in situ и с гибридами соматических клеток». Цитогенетика и клеточная генетика . 79 (3–4): 228–230. дои : 10.1159/000134729 . ПМИД 9605859 .
- ^ Скотт Э.К., Рейтер Дж.Э., Луо Л. (апрель 2003 г.). «Малая ГТФаза Cdc42 необходима для многих аспектов дендритного морфогенеза» . Журнал неврологии . 23 (8): 3118–3123. doi : 10.1523/JNEUROSCI.23-08-03118.2003 . ПМЦ 6742332 . ПМИД 12716918 .
- ^ Калабрезе Б., Уилсон М.С., Халпейн С. (февраль 2006 г.). «Развитие и регуляция дендритных синапсов позвоночника». Физиология . 21 (1): 38–47. дои : 10.1152/физиол.00042.2005 . ПМИД 16443821 .
- ^ Де Роо М., Клаузер П., Мендес П., Полья Л., Мюллер Д. (январь 2008 г.). «Зависимое от активности формирование PSD и стабилизация вновь сформированных шипов в культурах срезов гиппокампа» . Кора головного мозга . 18 (1): 151–161. дои : 10.1093/cercor/bhm041 . ПМИД 17517683 .
- ^ Канеко М., Се Ю, Ан Джей Джей, Страйкер, член парламента, Сюй Б (апрель 2012 г.). «Дендритный синтез BDNF необходим для поздней фазы созревания позвоночника и восстановления корковых реакций после сенсорной депривации» . Журнал неврологии . 32 (14): 4790–4802. doi : 10.1523/JNEUROSCI.4462-11.2012 . ПМЦ 3356781 . ПМИД 22492034 .
- ^ Jump up to: Перейти обратно: а б с Сюй Т, Ю Х, Перлик А.Дж., Тобин В.Ф., Цвейг Дж.А., Теннант К. и др. (декабрь 2009 г.). «Быстрое формирование и избирательная стабилизация синапсов для устойчивых двигательных воспоминаний» . Природа . 462 (7275): 915–919. Бибкод : 2009Natur.462..915X . дои : 10.1038/nature08389 . ПМЦ 2844762 . ПМИД 19946267 .
- ^ Jump up to: Перейти обратно: а б с Робертс Т.Ф., Чида К.А., Кляйн М.Е., Муни Р. (февраль 2010 г.). «Быстрая стабилизация позвоночника и усиление синапсов в начале поведенческого обучения» . Природа . 463 (7283): 948–952. Бибкод : 2010Natur.463..948R . дои : 10.1038/nature08759 . ПМЦ 2918377 . ПМИД 20164928 .
- ^ Чида К.А., Муни Р. (март 2012 г.). «Оглушение вызывает специфичные для типа клеток изменения в дендритных шипиках сенсомоторного ядра, важные для обучения вокализации» . Нейрон . 73 (5): 1028–1039. дои : 10.1016/j.neuron.2011.12.038 . ПМК 3299981 . ПМИД 22405211 .
- ^ Де Роо М., Клаузер П., Мюллер Д. (сентябрь 2008 г.). «LTP способствует выборочной долгосрочной стабилизации и кластеризации дендритных шипов» . ПЛОС Биология . 6 (9): е219. дои : 10.1371/journal.pbio.0060219 . ПМЦ 2531136 . ПМИД 18788894 .
- ^ Jump up to: Перейти обратно: а б с д и Цзо Ю, Линь А, Чанг П, Ган ВБ (апрель 2005 г.). «Развитие долгосрочной стабильности дендритных позвонков в различных областях коры головного мозга» . Нейрон . 46 (2): 181–189. дои : 10.1016/j.neuron.2005.04.001 . ПМИД 15848798 . S2CID 16232150 .
- ^ Холтмаат А.Дж., Трахтенберг Дж.Т., Уилбрехт Л., Шеперд Г.М., Чжан Х., Нотт Г.В., Свобода К. (январь 2005 г.). «Переходные и постоянные дендритные шипики в неокортексе in vivo» . Нейрон . 45 (2): 279–291. дои : 10.1016/j.neuron.2005.01.003 . ПМИД 15664179 . S2CID 13320649 .
- ^ Jump up to: Перейти обратно: а б с Ян Джи, Пан Ф, Ган ВБ (декабрь 2009 г.). «Стабильно поддерживаемые дендритные шипики связаны с воспоминаниями на всю жизнь» . Природа . 462 (7275): 920–924. Бибкод : 2009Natur.462..920Y . дои : 10.1038/nature08577 . ПМЦ 4724802 . ПМИД 19946265 .
- ^ Холтмаат А., Вилбрехт Л., Нотт Г.В., Велкер Э., Свобода К. (июнь 2006 г.). «Рост позвоночника в неокортексе, зависящий от опыта и зависящий от типа клеток». Природа . 441 (7096): 979–983. Бибкод : 2006Natur.441..979H . дои : 10.1038/nature04783 . ПМИД 16791195 . S2CID 4428322 .
- ^ Браун С.Э., Ли П., Бойд Дж.Д., Делани К.Р., Мерфи Т.Х. (апрель 2007 г.). «Обширный оборот дендритных шипов и ремоделирование сосудов в кортикальных тканях, восстанавливающихся после инсульта» . Журнал неврологии . 27 (15): 4101–4109. doi : 10.1523/JNEUROSCI.4295-06.2007 . ПМК 6672555 . ПМИД 17428988 .
- ^ Браун CE, Мерфи TH (апрель 2008 г.). «Жизнь на грани: визуализация дендритного оборота позвоночника в периинфарктной зоне во время ишемического инсульта и восстановления». Нейробиолог . 14 (2): 139–146. дои : 10.1177/1073858407309854 . ПМИД 18039977 . S2CID 46267737 .
- ^ Jump up to: Перейти обратно: а б Бхатт Д.Х., Чжан С., Ган В.Б. (2009). «Динамика дендритного позвоночника». Ежегодный обзор физиологии . 71 : 261–282. doi : 10.1146/annurev.physol.010908.163140 . ПМИД 19575680 .
- ^ Jump up to: Перейти обратно: а б с Бенсон, Кертис А.; Кинг, Джаред Ф.; Реймер, Марике Л.; Кауэр, Сьерра Д.; Ваксман, Стивен Г.; Тан, Эндрю М. (3 декабря 2022 г.). «Дендритные шипы и болевая память» . Нейробиолог : 107385842211382. doi : 10.1177/10738584221138251 . ISSN 1073-8584 . ПМИД 36461773 .
- ^ фон Болен и Хальбах О., Захер С., Гасс П., Унсикер К. (март 2006 г.). «Возрастные изменения в шипах гиппокампа и недостатки пространственной памяти у мышей». Журнал нейробиологических исследований . 83 (4): 525–531. дои : 10.1002/мл.20759 . ПМИД 16447268 . S2CID 30838296 .
- ^ Бенсон, Кертис А.; Фенрих, Кейт К.; Олсон, Кай-Лан; Патва, Сирадж; Бангалор, Лакшми; Ваксман, Стивен Г.; Тан, Эндрю М. (27 мая 2020 г.). «Динритная динамика позвоночника после травмы периферических нервов: прижизненное структурное исследование» . Журнал неврологии . 40 (22): 4297–4308. doi : 10.1523/JNEUROSCI.2858-19.2020 . ISSN 0270-6474 . ПМЦ 7252482 . ПМИД 32371602 .
- ^ Цао, Сяоюй С; Паппалардо, Лаура В.; Ваксман, Стивен Дж; Тан, Эндрю М. (январь 2017 г.). «Дендритная дисгенезия позвоночника в поверхностных сенсорных нейронах дорсального рога после травмы спинного мозга» . Молекулярная боль . 13 : 174480691668801. doi : 10.1177/1744806916688016 . ISSN 1744-8069 . ПМК 5302173 . ПМИД 28326929 .
- ^ Чжао, Пэн; Хилл, Мириам; Лю, Шуцзюнь; Чен, Любин; Бангалор, Лакшми; Ваксман, Стивен Г.; Тан, Эндрю М. (01 июня 2016 г.). «Ремоделирование дендритов позвоночника после раннего и позднего ингибирования Rac1 после травмы спинного мозга: данные о биомаркере боли» . Журнал нейрофизиологии . 115 (6): 2893–2910. дои : 10.1152/Jn.01057.2015 . ISSN 0022-3077 . ПМК 4922610 . ПМИД 26936986 .
- ^ Тан, Эндрю М.; Чой, Джин-Сун; Ваксман, Стивен Г.; Хейнс, Брайан К. (октябрь 2009 г.). «Ремоделирование дендритов позвоночника после травмы спинного мозга изменяет обработку нейрональных сигналов» . Журнал нейрофизиологии . 102 (4): 2396–2409. дои : 10.1152/jn.00095.2009 . ISSN 0022-3077 . ПМИД 19692517 .
- ^ Харрис К.М., Фиала Дж.К., Острофф Л. (апрель 2003 г.). «Структурные изменения в дендритных синапсах позвоночника во время длительной потенциации» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 358 (1432): 745–748. дои : 10.1098/rstb.2002.1254 . ПМК 1693146 . ПМИД 12740121 .
- ^ Арайя Р., Николенко В., Эйзенталь К.Б., Юсте Р. (июль 2007 г.). «Натриевые каналы усиливают потенциалы позвоночника» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (30): 12347–12352. Бибкод : 2007PNAS..10412347A . дои : 10.1073/pnas.0705282104 . ЧВК 1924793 . ПМИД 17640908 .
- ^ Нго-Ань Т.Дж., Бладгуд Б.Л., Лин М., Сабатини Б.Л., Мэйли Дж., Адельман Дж.П. (май 2005 г.). «SK-каналы и NMDA-рецепторы образуют Ca2+-опосредованную петлю обратной связи в дендритных шипиках». Природная неврология . 8 (5): 642–649. дои : 10.1038/nn1449 . ПМИД 15852011 . S2CID 385712 .
- ^ Юсте Р., Денк В. (июнь 1995 г.). «Дендритные шипики как основные функциональные единицы нейрональной интеграции». Природа . 375 (6533): 682–684. Бибкод : 1995Natur.375..682Y . дои : 10.1038/375682a0 . ПМИД 7791901 . S2CID 4271356 .
- ^ Jump up to: Перейти обратно: а б Бивалес В.Г., Патирниче Д., Рупрехт В., Стеммлер М., Герц А.В., Палфи Д. и др. (февраль 2015 г.). «Локальная постсинаптическая активация потенциалзависимых натриевых каналов в дендритных шипиках гранулярных клеток обонятельной луковицы» . Нейрон . 85 (3): 590–601. дои : 10.1016/j.neuron.2014.12.051 . ПМИД 25619656 .
- ^ Баер С.М., Ринцель Дж. (апрель 1991 г.). «Распространение дендритных шипов, опосредованное возбудимыми шипами: теория континуума». Журнал нейрофизиологии . 65 (4): 874–890. дои : 10.1152/jn.1991.65.4.874 . ПМИД 2051208 .
- ^ Бресслофф ПК, Кумбс С (2000). «Уединенные волны в модели дендритного кабеля с активными шипами». SIAM Journal по прикладной математике . 61 (2): 432–453. CiteSeerX 10.1.1.104.1307 . дои : 10.1137/s0036139999356600 . JSTOR 3061734 . S2CID 3058796 .
- ^ Невиан Т., Сакманн Б. (октябрь 2006 г.). «Передача сигналов Ca2+ в позвоночнике в пластичности, зависящей от времени спайка» . Журнал неврологии . 26 (43): 11001–11013. doi : 10.1523/JNEUROSCI.1749-06.2006 . ПМК 6674669 . ПМИД 17065442 .
- ^ Грюндиц А., Холбро Н., Тиан Л., Цзо Й., Эртнер Т.Г. (декабрь 2008 г.). «Пластичность шеи позвоночника контролирует постсинаптические сигналы кальция посредством электрической компартментализации» . Журнал неврологии . 28 (50): 13457–13466. doi : 10.1523/JNEUROSCI.2702-08.2008 . ПМК 6671740 . ПМИД 19074019 .
- ^ Jump up to: Перейти обратно: а б Ёшихара Ю., Де Роо М., Мюллер Д. (апрель 2009 г.). «Формирование и стабилизация дендритного позвоночника». Современное мнение в нейробиологии . 19 (2): 146–53. дои : 10.1016/j.conb.2009.05.013 . ПМИД 19523814 . S2CID 5054448 .
- ^ Саарикангас Дж., Курдугли Н., Сенджу Ю., Чазал Г., Сегерстроле М., Минкевичене Р. и др. (июнь 2015 г.). «Вызванный MIM изгиб мембраны способствует инициации дендритных отростков» . Развивающая клетка . 33 (6): 644–659. дои : 10.1016/j.devcel.2015.04.014 . ПМИД 26051541 .
- ^ Пелучки С., Стринги Р., Марчелло Э. (январь 2020 г.). «Дендритные шипы при болезни Альцгеймера: как актиновый цитоскелет способствует синаптической недостаточности» . Международный журнал молекулярных наук . 21 (3): 908. doi : 10.3390/ijms21030908 . ПМК 7036943 . ПМИД 32019166 .
- ^ Пензес П., Кэхилл М.Э., Джонс К.А., ВанЛеувен Дж.Э., Вулфри К.М. (март 2011 г.). «Дендритная патология позвоночника при нервно-психических расстройствах» . Природная неврология . 14 (3): 285–293. дои : 10.1038/nn.2741 . ПМЦ 3530413 . ПМИД 21346746 .
- ^ Рамон-и-Кахаль С. (1888). «Строение нервных центров птиц». Преподобный Трим. Гистол. Норм . 1 :1–10.
- ^ Грей Э.Г. (июнь 1959 г.). «Электронная микроскопия синаптических контактов на дендритных отростках коры головного мозга». Природа . 183 (4675): 1592–1593. Бибкод : 1959Natur.183.1592G . дои : 10.1038/1831592a0 . PMID 13666826 . S2CID 4258584 .
- ^ Дэйли М.Э., Смит С.Дж. (май 1996 г.). «Динамика дендритной структуры в развивающихся срезах гиппокампа» . Журнал неврологии . 16 (9): 2983–2994. doi : 10.1523/JNEUROSCI.16-09-02983.1996 . ПМК 6579052 . ПМИД 8622128 .
- ^ Бонхёффер Т., Юсте Р. (сентябрь 2002 г.). «Подвижность позвоночника. Феноменология, механизмы и функции» . Нейрон . 35 (6): 1019–1027. дои : 10.1016/s0896-6273(02)00906-6 . ПМИД 12354393 . S2CID 10183317 .
Дальнейшее чтение [ править ]
- Левитан И.Б., Качмарек Л.К. (2002). Нейрон: клеточная и молекулярная биология (Третье изд.). Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-514522-9 .
- Судхоф ТК, Стивенс КФ, Коуэн ВМ (2001). Синапсы . Балтимор: Издательство Университета Джона Хопкинса. ISBN 978-0-8018-6498-8 .
- Касаи Х., Мацузаки М., Ногучи Дж., Ясумацу Н. (октябрь 2002 г.). «[Дендритные структуры и функции позвоночника]». Нихон Синкей Сейсин Якуригаку Засси = Японский журнал психофармакологии . 22 (5): 159–164. ПМИД 12451686 .
- Лишофф С., Бишоф Х.Дж. (март 2003 г.). «Динамика изменения плотности позвоночника». Поведенческие исследования мозга . 140 (1–2): 87–95. дои : 10.1016/S0166-4328(02)00271-1 . ПМИД 12644282 . S2CID 2275781 .
- Линч Дж., Рекс К.С., Галл К.М. (январь 2007 г.). «Консолидация LTP: субстраты, объяснительная сила и функциональное значение». Нейрофармакология . 52 (1): 12–23. doi : 10.1016/j.neuropharm.2006.07.027 . ПМИД 16949110 . S2CID 22652804 .
- Мацузаки М., Хонкура Н., Эллис-Дэвис Г.К., Касаи Х. (июнь 2004 г.). «Структурные основы долговременной потенциации в одиночных дендритных шипиках» . Природа . 429 (6993): 761–766. Бибкод : 2004Natur.429..761M . дои : 10.1038/nature02617 . ПМК 4158816 . ПМИД 15190253 .
- Нимчинский Е.А., Сабатини Б.Л., Свобода К (2002). «Строение и функция дендритных шипиков». Ежегодный обзор физиологии . 64 : 313–353. doi : 10.1146/annurev.phyol.64.081501.160008 . ПМИД 11826272 .
- Юсте Р., Маевска А., Холтхофф К. (июль 2000 г.). «От формы к функции: компартментализация кальция в дендритных шипиках». Природная неврология . 3 (7): 653–659. дои : 10.1038/76609 . ПМИД 10862697 . S2CID 33466678 .