Пирамидальная клетка
| Пирамидальная клетка | |
|---|---|
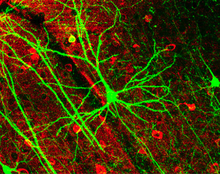
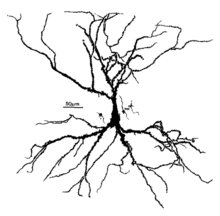
 пирамидные клетки человека Неокортексальные , окрашенные по методу Гольджи . Апикальный дендрит простирается вертикально над сомой (телом клетки), а многочисленные базальные дендриты расходятся вбок от основания тела клетки. | |
| Подробности | |
| Расположение | Кора головного мозга, особенно слои III и V |
| Форма | Многополярный пирамидальный |
| Функция | Возбуждающий проекционный нейрон |
| Нейромедиатор | Глутамат , ГАМК |
| Идентификаторы | |
| МеШ | D017966 |
| НейроЛекс ID | sao862606388 |
| ТД | H1.00.01.0.00044 |
| ФМА | 84105 |
| Анатомические термины нейроанатомии | |
Пирамидные клетки , или пирамидальные нейроны , представляют собой тип мультиполярных нейронов, обнаруженных в областях мозга, включая кору головного мозга , гиппокамп и миндалевидное тело . млекопитающих Пирамидные клетки являются первичными единицами возбуждения префронтальной коры и кортикоспинального тракта . Одной из основных структурных особенностей пирамидного нейрона является коническая форма сомы , или тела клетки, в честь которой нейрон назван. Другими ключевыми структурными особенностями пирамидной клетки являются одиночный аксон , большой апикальный дендрит , множественные базальные дендриты и наличие дендритных шипов . [1]
Пирамидные нейроны также являются одним из двух типов клеток, характерный признак которых — тельца Негри — обнаруживается при посмертной инфекции бешенства. [2] Пирамидные нейроны были впервые обнаружены и изучены Сантьяго Рамоном-и-Кахалем . [3] [4] С тех пор исследования пирамидных нейронов были сосредоточены на самых разных темах — от нейропластичности до когнитивных функций .
Структура [ править ]

Одной из основных структурных особенностей пирамидного нейрона является коническая форма сомы , или тела клетки, в честь которой нейрон назван. Другими ключевыми структурными особенностями пирамидной клетки являются одиночный аксон , большой апикальный дендрит , множественные базальные дендриты и наличие дендритных шипов . [1]
Апикальный дендрит [ править ]
Апикальный дендрит поднимается от вершины сомы пирамидной клетки. Апикальный дендрит представляет собой одиночный длинный и толстый дендрит, который разветвляется несколько раз по мере увеличения расстояния от сомы и распространяется к поверхности коры. [1]
Базальный дендрит [ править ]
Базальные дендриты возникают из основания сомы. Базальное дендритное дерево состоит из трех-пяти первичных дендритов. По мере увеличения расстояния от сомы базальные дендриты обильно ветвятся. [1]
Пирамидные клетки являются одними из крупнейших нейронов головного мозга. Как у людей, так и у грызунов длина пирамидальных клеточных тел (сом) составляет в среднем около 20 мкм. Пирамидальные дендриты обычно имеют диаметр от полмикрометра до нескольких микрометров. Длина одного дендрита обычно составляет несколько сотен микрометров. За счет ветвления общая дендритная длина пирамидальной клетки может достигать нескольких сантиметров. Аксон пирамидной клетки часто еще длиннее и сильно разветвлен, достигая общей длины многих сантиметров.
Дендритные шипы [ править ]
Дендритные шипики получают большую часть возбуждающих импульсов ( ВПСП ), которые попадают в пирамидную клетку. Дендритные шипы были впервые обнаружены Рамоном-и-Кахалем в 1888 году с использованием метода Гольджи . Рамон-и-Кахаль также был первым, кто предположил физиологическую роль увеличения площади рецептивной поверхности нейрона. Чем больше площадь поверхности пирамидной клетки, тем выше способность нейрона обрабатывать и интегрировать большие объемы информации. Дендритные шипы на соме отсутствуют, а по мере удаления от нее их число увеличивается. [4] Типичный апикальный дендрит крысы имеет не менее 3000 дендритных шипов. Средний апикальный дендрит человека примерно в два раза длиннее крысиного, поэтому количество дендритных шипов, присутствующих на апикальном дендрите человека, может достигать 6000. [5]
Рост развитие и
Дифференциация [ править ]
Пирамидальная спецификация возникает на ранних стадиях развития головного мозга. Клетки-предшественники коммитируются в нейрональную линию в подкорковой пролиферативной желудочковой зоне (VZ) и субвентрикулярной зоне (SVZ). Незрелые пирамидные клетки мигрируют и занимают кортикальную пластинку , где они далее диверсифицируются. Эндоканнабиноиды (eCB) представляют собой один класс молекул, которые, как было показано, управляют развитием пирамидных клеток и поиском аксональных путей. [6] Было показано, что факторы транскрипции, такие как Ctip2 и Sox5, способствуют направлению, в котором пирамидные нейроны направляют свои аксоны. [7]
Раннее постнатальное развитие [ править ]
Было показано, что пирамидные клетки у крыс претерпевают множество быстрых изменений в раннем постнатальном периоде. Было показано, что между 3 и 21 днями постнатального развития пирамидные клетки удваивают размер сомы, увеличивают длину апикального дендрита в пять раз и увеличивают длину базального дендрита в тринадцать раз. мембраны Другие изменения включают снижение потенциала покоя , уменьшение сопротивления мембраны и увеличение пиковых значений потенциалов действия . [8]
Сигнализация [ править ]
Как и дендриты в большинстве других нейронов, дендриты обычно являются входными областями нейрона, а аксон — выходными. И аксоны, и дендриты сильно разветвлены. Большое количество разветвлений позволяет нейрону отправлять и получать сигналы от множества разных нейронов.
Пирамидные нейроны, как и другие нейроны, имеют многочисленные потенциалзависимые ионные каналы . В пирамидных клетках содержится большое количество Na. + , Как 2+ и К + каналы в дендритах и некоторые каналы в соме. [9] [10] Ионные каналы внутри дендритов пирамидальных клеток имеют свойства, отличные от свойств ионных каналов одного и того же типа в соме пирамидных клеток. [11] [12] Напряжение-управляемый Ca 2+ каналы в дендритах пирамидных клеток активируются подпороговыми ВПСП и потенциалами обратного распространения . Степень обратного распространения потенциалов действия внутри пирамидальных дендритов зависит от K + каналы. К + каналы в дендритах пирамидных клеток обеспечивают механизм контроля амплитуды потенциалов действия. [13]
Способность пирамидных нейронов интегрировать информацию зависит от количества и распределения получаемых ими синаптических входов. Одна пирамидальная клетка получает около 30 000 возбуждающих входов и 1700 тормозных ( IPSP ) входов. Возбуждающие (ВПСП) входы заканчиваются исключительно на дендритных шипиках, тогда как тормозящие (ТПСП) входы заканчиваются на дендритных стержнях, соме и даже аксоне. Пирамидные нейроны могут возбуждаться нейромедиатором глутаматом . [1] [14] и ингибируется нейромедиатором ГАМК . [1]

Классификации стрельбы [ править ]
Пирамидные нейроны были разделены на различные подклассы в зависимости от их реакции на импульсы тока длительностью 400–1000 миллисекунд. Это нейроны RSad, RSna и IB.
Рсад [ править ]
Пирамидные нейроны RSad, или адаптирующиеся регулярные импульсные нейроны , активируются отдельными потенциалами действия (ПД), за которыми следует гиперполяризующий послепотенциал. Послепотенциал увеличивается по продолжительности, что создает частоты спайков адаптацию (SFA) в нейроне. [15]
РСна [ править ]
Пирамидные нейроны RSna, или неадаптирующиеся регулярные импульсные нейроны, после импульса запускают серию потенциалов действия. Эти нейроны не проявляют признаков адаптации. [15]
ИБ [ править ]
Пирамидные нейроны IB, или взрывные нейроны, реагируют на пороговые импульсы вспышкой из двух-пяти быстрых потенциалов действия. Пирамидные нейроны IB не обнаруживают адаптации. [15]
Молекулярные классификации
Есть несколько исследований, показывающих, что морфологические и электрические свойства пирамидных клеток можно определить на основе экспрессии генов, измеренной с помощью секвенирования отдельных клеток . [16] В нескольких исследованиях предлагается классификация отдельных клеток у мышей. [17] и человек [18] нейроны, основанные на экспрессии генов, могли бы объяснить различные свойства нейронов. Типы нейронов в этих классификациях разделены на возбуждающие, тормозные и сотни соответствующих суббайтов. Например, пирамидные клетки 2-3 слоя у человека относятся к типу FREM3. [16] и часто имеют большое количество Ih-тока [19] генерируется HCN-каналом .
Функция [ править ]
Корково-спинномозговой путь [ править ]
Пирамидные нейроны являются основным типом нервных клеток в кортикоспинальном тракте . Нормальный двигательный контроль зависит от развития связей между аксонами кортикоспинального тракта и спинного мозга. Аксоны пирамидных клеток следуют таким сигналам, как факторы роста, для создания определенных связей. При правильном соединении пирамидные клетки принимают участие в схеме, отвечающей за двигательную функцию, управляемую зрением. [20]
Познание [ править ]
Пирамидные нейроны префронтальной коры участвуют в когнитивных способностях. У млекопитающих сложность пирамидных клеток увеличивается от задних к передним отделам мозга. Степень сложности пирамидных нейронов, вероятно, связана с когнитивными способностями различных видов антропоидов. Пирамидные клетки в префронтальной коре, по-видимому, отвечают за обработку входных данных от первичной слуховой коры, первичной соматосенсорной коры и первичной зрительной коры, которые обрабатывают сенсорные модальности. [21] Эти клетки могут также играть решающую роль в распознавании сложных объектов в областях зрительной обработки коры. [3] По сравнению с другими видами, больший размер клеток и сложность пирамидных нейронов, а также определенные закономерности клеточной организации и функций коррелируют с эволюцией человеческого познания. [22]
и обучение Память
Пирамидные клетки гиппокампа необходимы для определенных типов памяти и обучения. Они образуют синапсы, которые помогают интегрировать синаптические напряжения в их сложные дендритные деревья посредством взаимодействия с мшистыми волокнами гранулярных клеток. Поскольку это влияет на постсинаптические напряжения, создаваемые активацией мшистых волокон, размещение шиповатых выростов на базальных и апикальных дендритах важно для формирования памяти. Обеспечивая динамический контроль чувствительности пирамидных клеток CA3, такое скопление синапсов мшистых волокон на пирамидальных клетках может способствовать инициации соматических спайков. Взаимодействия между пирамидальными клетками и примерно 41 бутоном мшистых волокон, каждый из которых происходит из уникальной гранулярной клетки, подчеркивают роль этих бутонов в обработке информации и синаптических связях, которые необходимы для памяти и обучения. По сути, входные сигналы мшистых волокон принимаются пирамидальными клетками гиппокампа, которые интегрируют синаптические напряжения в свою дендритную архитектуру. Расположение колючих выступов и скопление синапсов влияют на чувствительность и способствуют обработке информации, касающейся памяти и обучения. [23]
См. также [ править ]
- Пирамидный тракт
- Клетки-люстры - иннервируют начальные сегменты пирамидных аксонов.
- Нейрон шиповника
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с д и ж Мегиас М., Эмри З., Фройнд Т.Ф., Гуляс А.И. (2001). «Общее количество и распределение тормозных и возбуждающих синапсов на пирамидальных клетках CA1 гиппокампа». Нейронаука . 102 (3): 527–540. дои : 10.1016/S0306-4522(00)00496-6 . ПМИД 11226691 . S2CID 16458290 .
- ^ Скетчи Групп, ООО. «2.3 рабдовирус» . ЭскизныйМедицинский . Архивировано из оригинала 13 апреля 2017 г.
- ^ Jump up to: Перейти обратно: а б Элстон Г.Н. (ноябрь 2003 г.). «Кора, познание и клетка: новое понимание пирамидного нейрона и префронтальной функции» . Кора головного мозга . 13 (11): 1124–1138. дои : 10.1093/cercor/bhg093 . ПМИД 14576205 .
- ^ Jump up to: Перейти обратно: а б Гарсиа-Лопес П., Гарсиа-Марин В., Фрейре М. (ноябрь 2006 г.). «Трёхмерная реконструкция и количественное исследование пирамидной клетки гистологического препарата Кахаля» . Журнал неврологии . 26 (44): 11249–11252. doi : 10.1523/JNEUROSCI.3543-06.2006 . ПМК 6674523 . ПМИД 17079652 .
- ^ Лаберж Д., Касевич Р. (ноябрь 2007 г.). «Апикальная дендритная теория сознания». Нейронные сети . 20 (9): 1004–1020. дои : 10.1016/j.neunet.2007.09.006 . ПМИД 17920812 .
- ^ Малдер Дж., Агуадо Т., Кеймпема Э., Барабас К., Баллестер Росадо С.Дж., Нгуен Л. и др. (июнь 2008 г.). «Передача сигналов эндоканнабиноидов контролирует спецификацию пирамидных клеток и формирование паттерна аксонов дальнего действия» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (25): 8760–8765. Бибкод : 2008PNAS..105.8760M . дои : 10.1073/pnas.0803545105 . ПМЦ 2438381 . ПМИД 18562289 .
- ^ Фишелл Дж., Ханашима К. (февраль 2008 г.). «Пирамидные нейроны взрослеют и меняют свое мнение» . Нейрон . 57 (3): 333–338. дои : 10.1016/j.neuron.2008.01.018 . ПМИД 18255026 . S2CID 15095100 .
- ^ Чжан З.В. (март 2004 г.). «Созревание пирамидных нейронов V слоя в префронтальной коре крыс: внутренние свойства и синаптическая функция». Журнал нейрофизиологии . 91 (3): 1171–1182. дои : 10.1152/jn.00855.2003 . ПМИД 14602839 .
- ^ Спрустон Н. (март 2008 г.). «Пирамидные нейроны: дендритная структура и синаптическая интеграция». Обзоры природы. Нейронаука . 9 (3): 206–221. дои : 10.1038/nrn2286 . ПМИД 18270515 . S2CID 1142249 .
- ^ Георгиев Д.Д., Колев С.К., Коэн Э., Глейзбрук Дж.Ф. (декабрь 2020 г.). «Вычислительная мощность пирамидных нейронов коры головного мозга». Исследования мозга . 1748 : 147069. arXiv : 2009.10615 . doi : 10.1016/j.brainres.2020.147069 . ПМИД 32858030 . S2CID 221277603 .
- ^ Голдинг Н.Л., Микус Т.Дж., Кац Ю., Кэт В.Л., Спрустон Н. (октябрь 2005 г.). «Факторы, опосредующие мощное ослабление напряжения вдоль дендритов пирамидных нейронов CA1» . Журнал физиологии . 568 (Часть 1): 69–82. дои : 10.1113/jphysicalol.2005.086793 . ПМЦ 1474764 . ПМИД 16002454 .
- ^ Реми С., Бек Х., Яари Ю. (август 2010 г.). «Пластичность потенциал-управляемых ионных каналов в дендритах пирамидальных клеток». Современное мнение в нейробиологии . 20 (4): 503–509. дои : 10.1016/j.conb.2010.06.006 . ПМИД 20691582 . S2CID 4713853 .
- ^ Маги Дж., Хоффман Д., Колберт С., Джонстон Д. (1998). «Электрическая и кальциевая передача сигналов в дендритах пирамидных нейронов гиппокампа». Ежегодный обзор физиологии . 60 (1): 327–346. doi : 10.1146/annurev.phyol.60.1.327 . ПМИД 9558467 .
- ^ Вонг, РКС; Трауб, Р.Д. (01.01.2009), «СЕТИ | Клеточные свойства и синаптическая связь пирамидных клеток CA3: механизмы эпилептической синхронизации и эпилептогенеза» , в Шварцкройне, Филип А. (ред.), Энциклопедия фундаментальных исследований эпилепсии , Оксфорд : Academic Press, стр. 815–819, doi : 10.1016/b978-012373961-2.00215-0 , ISBN. 978-0-12-373961-2 , получено 18 ноября 2020 г.
- ^ Jump up to: Перейти обратно: а б с Франческетти С., Санчини Дж., Панзика Ф., Радичи К., Аванзини Дж. (апрель 1998 г.). «Постнатальная дифференциация огневых свойств и морфологических характеристик пирамидных нейронов V слоя сенсомоторной коры». Нейронаука . 83 (4): 1013–1024. дои : 10.1016/S0306-4522(97)00463-6 . ПМИД 9502243 . S2CID 6986307 .
- ^ Jump up to: Перейти обратно: а б Берг Дж., Соренсен С.А., Тинг Дж.Т., Миллер Дж.А., Чартран Т., Бучин А. и др. (октябрь 2021 г.). «Расширение неокортекса человека предполагает диверсификацию глутаматергических нейронов» . Природа . 598 (7879): 151–158. Бибкод : 2021Natur.598..151B . дои : 10.1038/s41586-021-03813-8 . ПМЦ 8494638 . ПМИД 34616067 .
- ^ Гувенс Н.В., Соренсен С.А., Берг Дж., Ли С., Ярски Т., Тинг Дж. и др. (июль 2019 г.). «Классификация электрофизиологических и морфологических типов нейронов зрительной коры мыши» . Природная неврология . 22 (7): 1182–1195. дои : 10.1038/s41593-019-0417-0 . ПМЦ 8078853 . ПМИД 31209381 .
- ^ Баккен Т.Э., Йорстад Н.Л., Ху К., Лейк Б.Б., Тиан В., Калмбах Б.Е. и др. (октябрь 2021 г.). «Сравнительный клеточный анализ моторной коры человека, мартышки и мыши» . Природа . 598 (7879): 111–119. Бибкод : 2021Natur.598..111B . дои : 10.1038/s41586-021-03465-8 . ПМЦ 8494640 . ПМИД 34616062 .
- ^ Калмбах Б.Е., Бучин А., Лонг Б., Клоуз Дж., Нанди А., Миллер Дж.А. и др. (декабрь 2018 г.). «h-каналы способствуют разным внутренним свойствам мембран супрагранулярных пирамидальных нейронов в коре головного мозга человека и мыши» . Нейрон . 100 (5): 1194–1208.e5. дои : 10.1016/j.neuron.2018.10.012 . ПМК 6447369 . ПМИД 30392798 . S2CID 53218514 .
- ^ Салими И., Фрил К.М., Мартин Дж.Х. (июль 2008 г.). «Стимуляция пирамидных путей восстанавливает нормальные связи кортикоспинального тракта и зрительно-моторные навыки после ранней постнатальной блокады активности моторной коры» . Журнал неврологии . 28 (29): 7426–7434. doi : 10.1523/JNEUROSCI.1078-08.2008 . ПМК 2567132 . ПМИД 18632946 .
- ^ Бейкер А., Калмбах Б., Моришима М., Ким Дж., Джуавинетт А., Ли Н., Дембров Н. (июнь 2018 г.). «Специализированные субпопуляции глубокослойных пирамидальных нейронов в неокортексе: соединение клеточных свойств с функциональными последствиями» . Журнал неврологии . 38 (24): 5441–5455. doi : 10.1523/JNEUROSCI.0150-18.2018 . ПМК 6001033 . ПМИД 29798890 .
- ^ Галахова А.А., Хант С., Уилберс Р., Хейер Д.Б., де Кок К.П., Мансвелдер Х.Д., Горюнова Н.А. (ноябрь 2022 г.). «Эволюция корковых нейронов, поддерживающих познание человека» . Тенденции в когнитивных науках . 26 (11): 909–922. дои : 10.1016/j.tics.2022.08.012 . ПМК 9561064 . ПМИД 36117080 .
- ^ Гонсалес, РБ; ДеЛеон Гальван, CJ; Рангел, Ю.М.; Клэйборн, Би Джей (12 февраля 2001 г.). «Распределение шиповатых разрастаний на пирамидных нейронах СА3 в гиппокампе крысы» . Журнал сравнительной неврологии . 430 (3): 357–368. doi : 10.1002/1096-9861(20010212)430:3<357::aid-cne1036>3.0.co;2-k . ISSN 0021-9967 . ПМИД 11169473 .