Гиперполяризация (биология)
Эта статья нуждается в дополнительных цитатах для проверки . ( май 2013 г. ) |
Гиперполяризация — это изменение клетки мембранного потенциала , которое делает его более отрицательным. Это противоположность деполяризации . Он ингибирует потенциалы действия , увеличивая стимул, необходимый для перемещения мембранного потенциала до порога потенциала действия.
Гиперполяризация часто вызывается оттоком K + ( катион ) через K + каналы или приток Cl – ( анион ) через Cl – каналы . С другой стороны, приток катионов , например Na + через На + каналы или Ca 2+ через Ка 2+ каналов , подавляет гиперполяризацию. Если в клетке есть Na + или Ca 2+ токи в состоянии покоя, то торможение этих токов также приведет к гиперполяризации. Эта реакция потенциалзависимого ионного канала и является тем, как достигается состояние гиперполяризации. В нейронах клетка входит в состояние гиперполяризации сразу после генерации потенциала действия. В состоянии гиперполяризации нейрон находится в рефрактерном периоде, который длится примерно 2 миллисекунды, в течение которого нейрон не способен генерировать последующие потенциалы действия. Натриево-калийные АТФазы перераспределяют К + и На + ионы до тех пор, пока мембранный потенциал не вернется к своему потенциалу покоя около –70 милливольт, после чего нейрон снова готов передать другой потенциал действия. [1]
Потенциал-управляемые ионные каналы и гиперполяризация
[ редактировать ]
Потенциал-управляемые ионные каналы реагируют на изменения мембранного потенциала. Потенциал-управляемые калиевые, хлоридные и натриевые каналы являются ключевыми компонентами генерации потенциала действия, а также гиперполяризации. Эти каналы работают, выбирая ион на основе электростатического притяжения или отталкивания, позволяя иону связываться с каналом. [2] При этом молекула воды, прикрепленная к каналу, высвобождается, и ион проходит через пору. Потенциалзависимые натриевые каналы открываются в ответ на раздражитель и снова закрываются. Это означает, что канал либо открыт, либо нет, части пути не открыт. Иногда канал закрывается, но его можно сразу же открыть снова (так называемое шлюзование канала), или он может быть закрыт без возможности немедленного повторного открытия (инактивация канала).
При потенциале покоя и потенциалзависимые натриевые, и калиевые каналы закрыты, но по мере того, как клеточная мембрана деполяризуется, потенциалзависимые натриевые каналы начинают открываться, и нейрон начинает деполяризоваться, создавая петлю обратной связи по току, известную как цикл Ходжкина . [2] Однако ионы калия естественным образом выходят из клетки, и если исходное событие деполяризации не было достаточно значительным, нейрон не генерирует потенциал действия. Однако если все натриевые каналы открыты, то нейрон становится в десять раз более проницаемым для натрия, чем для калия, что быстро деполяризует клетку до пика +40 мВ. [2] На этом уровне натриевые каналы начинают инактивироваться, а потенциалзависимые калиевые каналы начинают открываться. Эта комбинация закрытых натриевых каналов и открытых калиевых каналов приводит к повторной поляризации нейрона и его сноваму отрицательному состоянию. Нейрон продолжает реполяризоваться до тех пор, пока клетка не достигнет ~ –75 мВ, [2] что представляет собой равновесный потенциал ионов калия. Это точка, в которой нейрон гиперполяризован, между –70 мВ и –75 мВ. После гиперполяризации калиевые каналы закрываются, и естественная проницаемость нейрона для натрия и калия позволяет нейрону вернуться к своему потенциалу покоя -70 мВ. Во время рефрактерного периода , то есть после гиперполяризации, но до того, как нейрон вернется к своему потенциалу покоя, нейрон способен вызвать потенциал действия благодаря способности открываться натриевых каналов, однако, поскольку нейрон более негативен, он становится труднее достичь порога потенциала действия.
Каналы HCN активируются гиперполяризацией.
Недавние исследования показали, что периоды рефрактерности нейронов могут превышать 20 миллисекунд, что ставит под сомнение связь между гиперполяризацией и рефрактерностью нейронов. [3] [4]
Экспериментальная методика
[ редактировать ]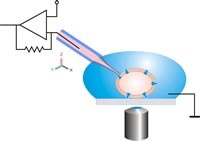
Гиперполяризация – это изменение мембранного потенциала. Нейробиологи измеряют его с помощью метода, известного как патч-фиксация , который позволяет им регистрировать ионные токи, проходящие через отдельные каналы. Это делается с помощью стеклянной микропипетки, также называемой патч-пипеткой, диаметром 1 микрометр. Есть небольшой участок, который содержит несколько ионных каналов, а остальная часть закрыта, что делает его точкой входа для тока. Использование усилителя и клещи напряжения , которая представляет собой электронную схему обратной связи, позволяет экспериментатору поддерживать мембранный потенциал в фиксированной точке, а затем клещи напряжения измеряют малейшие изменения в потоке тока. Мембранные токи, вызывающие гиперполяризацию, представляют собой либо увеличение внешнего тока, либо уменьшение входящего тока. [2]
Примеры
[ редактировать ]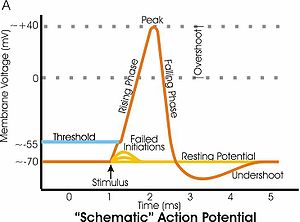
- В период послегиперполяризации после потенциала действия мембранный потенциал более отрицателен, чем когда клетка находится в состоянии покоя . На рисунке справа этот провал происходит примерно через 3–4 миллисекунды (мс) по временной шкале. Постгиперполяризация — это время, когда мембранный потенциал гиперполяризуется относительно потенциала покоя.
- Во время фазы нарастания потенциала действия мембранный потенциал меняется с отрицательного на положительный, что приводит к деполяризации. На рисунке нарастающая фаза составляет примерно от 1 до 2 мс на графике. Во время фазы подъема, когда мембранный потенциал становится положительным, мембранный потенциал продолжает деполяризоваться (превышать допустимое значение) до тех пор, пока пик потенциала действия не будет достигнут примерно на уровне +40 милливольт (мВ). После пика потенциала действия гиперполяризация реполяризует мембранный потенциал до значения покоя, сначала делая его менее положительным, пока не будет достигнуто 0 мВ, а затем продолжая делать его более отрицательным. Эта реполяризация происходит на рисунке примерно через 2–3 мс по шкале времени.
Ссылки
[ редактировать ]- ^ Пак, Филип Э. «Cliffs AP Biology, 3-е издание»
- ^ Jump up to: а б с д и Беккер, В.М., Кляйнсмит, Л.Дж., Хардин, Дж., и Бертони, GP (2009). Механизмы передачи сигнала: I. Электрическая и синаптическая передача сигналов в нейронах. Мир клетки (7-е изд.,). Сан-Франциско: Пирсон/Бенджамин Каммингс.
- ^ Варди, Рони; Тугендхафт, Яэль; Сарди, Шира; Кантер, Идо (1 июня 2021 г.). «Значительная анизотропная пластичность рефрактерного периода нейронов» . EPL (Письма по еврофизике) . 134 (6): 60007. arXiv : 2109.02041 . дои : 10.1209/0295-5075/ac177a . ISSN 0295-5075 . S2CID 237408101 .
- ^ Сарди, Шира; Варди, Рони; Тугендхафт, Яэль; Шейнин, Антон; Голденталь, Амир; Кантер, Идо (3 января 2022 г.). «Длительные анизотропные периоды абсолютной рефрактерности с быстрым временем нарастания для надежного реагирования» . Физический обзор E . 105 (1): 014401. arXiv : 2111.02689 . Бибкод : 2022PhRvE.105a4401S . дои : 10.1103/PhysRevE.105.014401 . ПМИД 35193251 . S2CID 242757511 .
Дальнейшее чтение
[ редактировать ]- Первс Д., Августин Г.Дж., Фитцпатрик Д. и др., ред. (2001). Нейронаука (2-е изд.). Сандерленд, Массачусетс: Sinauer Assoc. ISBN 0-87893-742-0 .
- Основные молекулярные, клеточные и медицинские аспекты нейрохимии, Сигел и др.