Калиевый канал

Калиевые каналы — наиболее широко распространенный тип ионных каналов, встречающийся практически у всех организмов. [1] Они образуют калийселективные , поры которые охватывают клеточные мембраны . Калиевые каналы обнаружены в большинстве типов клеток и контролируют широкий спектр клеточных функций. [2] [3]
Функция
[ редактировать ]Калиевые каналы проводят ионы калия вниз по их электрохимическому градиенту , причем делают это быстро (вплоть до скорости диффузии K + ионы в объеме воды) и избирательно (исключая, прежде всего, натрий, несмотря на субангстремную разницу в ионных радиусах). [4] С биологической точки зрения эти каналы устанавливают или сбрасывают потенциал покоя во многих клетках. В возбудимых клетках, таких как нейроны , задержанный противоток ионов калия формирует потенциал действия .
Способствуя регуляции продолжительности сердечного потенциала действия в сердечной мышце , нарушение работы калиевых каналов может вызвать опасные для жизни аритмии . Калиевые каналы также могут участвовать в поддержании сосудистого тонуса .
Они также регулируют клеточные процессы, такие как секреция гормонов ( например , высвобождение инсулина из бета-клеток поджелудочной железы ), поэтому их сбой может привести к заболеваниям (например, диабету ).
Некоторые токсины, такие как дендротоксин , являются мощными, поскольку блокируют калиевые каналы. [5]
Типы
[ редактировать ]Существует четыре основных класса калиевых каналов:
- Кальций-активируемый калиевый канал – открывается в ответ на присутствие ионов кальция или других сигнальных молекул.
- Внутреннее выпрямление калиевого канала - легче пропускает ток (положительный заряд) во внутрь (в клетку).
- Калиевые каналы домена тандемных пор - конститутивно открыты или обладают высокой базальной активацией, например, «калиевые каналы покоя» или «каналы утечки», которые устанавливают отрицательный мембранный потенциал нейронов.
- Потенциал-управляемые калиевые каналы — это потенциал-управляемые ионные каналы , которые открываются или закрываются в ответ на изменения трансмембранного напряжения .
В следующей таблице приведено сравнение основных классов калиевых каналов с репрезентативными примерами (полный список каналов в каждом классе см. на страницах соответствующих классов).
Дополнительные примеры фармакологических модуляторов калиевых каналов см. в разделах «Блокатор калиевых каналов» и «Открыватель калиевых каналов» .
| Сорт | Подклассы | Функция | Блокаторы | Активаторы |
|---|---|---|---|---|
| Активированный кальцием 6 Т и 1 П |
| [ нужна ссылка ]
| ||
| Внутренне исправляя 2 Т и 1 П |
|
| ||
|
| |||
|
| |||
|
| [ нужна ссылка ] | ||
| Тандемная пористая область 4 Т и 2 П |
|
| [ нужна ссылка ] | |
| Закрытый по напряжению 6 Т и 1 П |
|
|
|
Структура
[ редактировать ]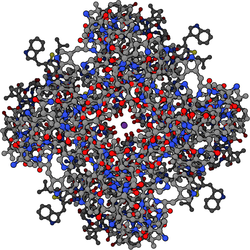
Калиевые каналы имеют тетрамерную структуру, в которой четыре идентичные белковые субъединицы объединяются с образованием четырехкратно симметричного ( С 4 ) комплекса, расположенного вокруг центральной ионпроводящей поры (т.е. гомотетрамера). Альтернативно четыре родственные, но не идентичные белковые субъединицы могут объединяться с образованием гетеротетрамерных комплексов с псевдо-C4 - симметрией. Все субъединицы калиевых каналов имеют характерную структуру пор-петлей, которая выстилает верхнюю часть поры и отвечает за селективную проницаемость для калия.
Существует более 80 млекопитающих генов калиевых каналов , которые кодируют субъединицы . Однако калиевые каналы, обнаруженные у бактерий, являются одними из наиболее изученных ионных каналов с точки зрения их молекулярной структуры. Используя рентгеновскую кристаллографию , [55] [56] Было получено глубокое понимание того, как ионы калия проходят через эти каналы и почему (меньшие) ионы натрия этого не делают. [57] 2003 года Нобелевская премия по химии была присуждена Роду Маккиннону за новаторскую работу в этой области. [58]
Селективный фильтр
[ редактировать ]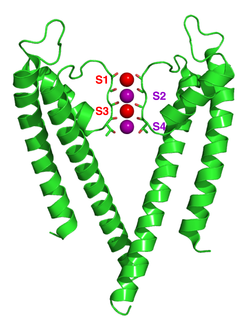
Каналы ионов калия удаляют гидратную оболочку с иона, когда он попадает в селективный фильтр. Селективный фильтр формируется последовательностью из пяти остатков, TVGYG, называемой сигнатурной последовательностью, в каждой из четырех субъединиц. Эта характерная последовательность находится внутри петли между спиралью поры и TM2/6, исторически называемой P-петлей. Эта сигнатурная последовательность высококонсервативна, за исключением того, что остаток валина в калиевых каналах прокариот часто заменяется остатком изолейцина в эукариотических каналах. Эта последовательность имеет уникальную структуру основной цепи, структурно аналогичную структурному мотиву белка гнезда . Четыре набора электроотрицательных карбонильных атомов кислорода ориентированы по направлению к центру поры фильтра и образуют квадратную антипризму, подобную водосольватирующей оболочке, вокруг каждого места связывания калия. Расстояние между карбонильными кислородами и ионами калия в местах связывания селективного фильтра такое же, как между кислородами воды в первой гидратной оболочке и ионом калия в водном растворе, что обеспечивает энергетически выгодный путь деградации. сольватация ионов. Однако ионы натрия слишком малы, чтобы заполнить пространство между карбонильными атомами кислорода. Таким образом, ионам натрия энергетически выгодно оставаться связанными с молекулами воды во внеклеточном пространстве, а не проходить через калийселективную ионную пору. [60] Эта ширина, по-видимому, поддерживается за счет водородных связей и сил Ван-дер-Ваальса внутри листа остатков ароматических аминокислот, окружающих селективный фильтр. [55] [61] Селективный фильтр открывается в сторону внеклеточного раствора, обнажая четыре карбонильных атома кислорода в остатке глицина (Gly79 в KcsA ). Следующий остаток, обращенный к внеклеточной стороне белка, — это отрицательно заряженный Asp80 (KcsA). Этот остаток вместе с пятью фильтрующими остатками образует пору, соединяющую заполненную водой полость в центре белка с внеклеточным раствором. [62]
Механизм селективности
[ редактировать ]Механизм селективности калиевых каналов остается предметом постоянных дискуссий. Карбонильные атомы кислорода сильно электроотрицательны и притягивают катионы. Фильтр может аккумулировать ионы калия в 4 местах, обычно обозначаемых от S1 до S4, начиная с внеклеточной стороны. Кроме того, один ион может связываться в полости в сайте, называемом SC, или один или несколько ионов на внеклеточной стороне в более или менее четко определенных сайтах, называемых S0 или Sext. Возможны различные варианты заполнения этих площадок. Поскольку рентгеновские структуры являются средними по многим молекулам, однако невозможно вывести фактическую заселенность непосредственно из такой структуры. В общем, есть некоторый недостаток из-за электростатического отталкивания, когда два соседних места заняты ионами. Предложения о механизме селективности были сделаны на основе молекулярно-динамического моделирования. [63] игрушечные модели связывания ионов, [64] термодинамические расчеты, [65] топологические соображения, [66] [67] и структурные различия [68] между селективными и неселективными каналами.
Механизм перемещения ионов в KcsA широко изучен с помощью теоретических расчетов и моделирования. [62] [69] Предсказание механизма ионной проводимости, в котором два дважды занятых состояния (S1, S3) и (S2, S4) играют существенную роль, было подтверждено обоими методами. Моделирование молекулярной динамики (MD) предполагает, что два внеклеточных состояния, S ext и S 0 , отражающие ионы, входящие и выходящие из фильтра, также являются важными участниками ионной проводимости.
Гидрофобная область
[ редактировать ]Эта область нейтрализует среду вокруг иона калия, так что он не притягивается к каким-либо зарядам. В свою очередь, это ускоряет реакцию.
Центральная полость
[ редактировать ]Центральная пора шириной 10 Å расположена вблизи центра трансмембранного канала, где энергетический барьер для поперечного иона самый высокий из-за гидрофобности стенки канала. Заполненная водой полость и полярный С-конец спиралей пор облегчают энергетический барьер для иона. Считается, что отталкивание предшествующих нескольких ионов калия способствует пропускной способности ионов.Наличие полости можно интуитивно понимать как один из механизмов преодоления канала диэлектрического барьера или отталкивания низкодиэлектрической мембраной за счет сохранения К + ион в водной, высокодиэлектрической среде.
Регулирование
[ редактировать ]
Поток ионов через поры калиевого канала регулируется двумя взаимосвязанными процессами, называемыми воротами и инактивацией. Гейтирование — это открытие или закрытие канала в ответ на стимулы, тогда как инактивация — это быстрое прекращение тока из открытого калиевого канала и подавление способности канала возобновлять проведение. Хотя оба процесса служат для регуляции проводимости каналов, каждый процесс может опосредоваться рядом механизмов.
Обычно считается, что шлюзование опосредовано дополнительными структурными доменами, которые воспринимают стимулы и, в свою очередь, открывают поры канала. Эти домены включают RCK-домены каналов BK, [70] [71] [72] и области датчика напряжения с управляемым по напряжению K + каналы. Считается, что эти домены реагируют на стимулы, физически открывая внутриклеточные ворота порового домена, тем самым позволяя ионам калия проходить через мембрану. Некоторые каналы имеют несколько регуляторных доменов или вспомогательных белков, которые могут модулировать ответ на стимул. Хотя механизмы продолжают обсуждаться, известны структуры ряда этих регуляторных доменов, включая RCK-домены прокариотических клеток. [73] [74] [75] и эукариотические [70] [71] [72] каналы, pH-воротный домен KcsA, [76] циклические нуклеотидные воротные домены, [77] и потенциалзависимые калиевые каналы. [78] [79]
Инактивация N-типа обычно является более быстрым механизмом инактивации и называется моделью «шара и цепи» . [80] Инактивация N-типа включает взаимодействие N-конца канала или связанного с ним белка, который взаимодействует с поровым доменом и перекрывает путь ионной проводимости наподобие «шарика». Альтернативно, инактивация C-типа, как полагают, происходит внутри самого селективного фильтра, где структурные изменения внутри фильтра делают его непроводящим. Существует ряд структурных моделей инактивированного K K C-типа. + фильтры каналов, [81] [82] [83] хотя точный механизм остается неясным.
Фармакология
[ редактировать ]Блокаторы
[ редактировать ]Блокаторы калиевых каналов ингибируют поток ионов калия через канал. Они либо конкурируют со связыванием калия внутри селективного фильтра, либо связываются снаружи фильтра, блокируя ионную проводимость. Примером одного из этих конкурентов являются ионы четвертичного аммония, которые связываются на внеклеточной поверхности. [84] [85] или центральную полость канала. [86] Ионы четвертичного аммония, блокирующие выход из центральной полости, также известны как блокаторы открытых каналов, поскольку связывание классически требует предварительного открытия цитоплазматических ворот. [87]
Ионы бария также могут блокировать токи калиевых каналов. [88] [89] путем связывания с высоким сродством внутри селективного фильтра. [90] [91] [92] [93] Считается, что это прочное связывание лежит в основе токсичности бария , ингибируя активность калиевых каналов в возбудимых клетках.
С медицинской точки зрения блокаторы калиевых каналов , такие как 4-аминопиридин и 3,4-диаминопиридин , исследовались для лечения таких состояний, как рассеянный склероз . [49] Нецелевое воздействие лекарств может привести к синдрому удлиненного интервала QT , индуцированному лекарственными средствами , — потенциально опасному для жизни состоянию. Чаще всего это происходит из-за воздействия на калиевый канал hERG в сердце. Соответственно, все новые лекарства проходят доклинические испытания на кардиобезопасность.
Активаторы
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( май 2019 г. ) |
Мускариновый калиевый канал
[ редактировать ]
Некоторые типы калиевых каналов активируются мускариновыми рецепторами и называются мускариновыми калиевыми каналами (I KACh ). Эти каналы представляют собой гетеротетрамер, состоящий из двух субъединиц GIRK1 и двух субъединиц GIRK4 . [94] [95] Примерами являются калиевые каналы в сердце, которые при активации парасимпатическими сигналами через мускариновые рецепторы М2 вызывают ток калия наружу, что замедляет частоту сердечных сокращений . [96] [97]
В изобразительном искусстве
[ редактировать ]Родерик Маккиннон заказал «Рождение идеи» — скульптуру высотой 5 футов (1,5 м), основанную на калиевом канале KcsA. [98] Работа содержит проволочный объект, изображающий внутреннюю часть канала, и объект из дутого стекла, изображающий основную полость конструкции канала.
См. также
[ редактировать ]- Кальциевый канал - комплекс ионных каналов, через который проходят ионы кальция.
- Канал ионов калия с внутренним выпрямителем - группа трансмембранных белков, которые пассивно транспортируют ионы калия.
- Калий в биологии - Использование калия организмами
- Семейство транспортеров калия (Trk) - семейство транспортных белков.
- Пермеаз, поглощающий калий - семейные
- Натрий-ионный канал - трансмембранный белок, пропускающий ионы натрия внутрь и наружу.
Ссылки
[ редактировать ]- ^ Литтлтон Дж. Т., Ганецкий Б. (апрель 2000 г.). «Ионные каналы и синаптическая организация: анализ генома дрозофилы» . Нейрон . 26 (1): 35–43. дои : 10.1016/S0896-6273(00)81135-6 . ПМИД 10798390 . S2CID 5694563 .
- ^ Хилле, Бертиль (2001). «Глава 5: Калийные и хлоридные каналы». Ионные каналы возбудимых мембран . Сандерленд, Массачусетс: Синауэр. стр. 131–168. ISBN 978-0-87893-321-1 .
- ^ Джесселл Т.М., Кандел Э.Р. , Шварц Дж.Х. (2000). «Глава 6: Ионные каналы». Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. стр. 105–124 . ISBN 978-0-8385-7701-1 .
- ^ Лим С., Дудев Т. (2016). Сигел А., Сигел Х., Сигел Р.К. (ред.). «Глава 10. Селективность по калию и натрию в фильтрах с селективностью моновалентных ионных каналов». Ионы щелочных металлов: их роль в жизни . Ионы металлов в науках о жизни. 16 . Спрингер: 325–347. дои : 10.1007/978-3-319-21756-7_9 . ISBN 978-3-319-21755-0 . ПМИД 26860305 .
- ^ косвенно цитируется по ссылке 3,4,5,6 в Рем Х, Лаздунски М (июль 1988 г.). «Очистка и структура субъединиц предполагаемого белка K +-канала, идентифицированного по его свойствам связывания с дендротоксином I» . Труды Национальной академии наук Соединенных Штатов Америки . 85 (13): 4919–4923. Бибкод : 1988PNAS...85.4919R . дои : 10.1073/pnas.85.13.4919 . ПМК 280549 . ПМИД 2455300 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Ранг, HP (2015). Фармакология (8-е изд.). Эдинбург: Черчилль Ливингстон. п. 59. ИСБН 978-0-443-07145-4 .
- ^ Томпсон Дж., Бегенисич Т. (май 2000 г.). «Электростатическое взаимодействие между харибдотоксином и тетрамерным мутантом каналов шейкера K(+)» . Биофизический журнал . 78 (5): 2382–2391. Бибкод : 2000BpJ....78.2382T . дои : 10.1016/S0006-3495(00)76782-8 . ПМЦ 1300827 . ПМИД 10777734 .
- ^ Наранхо Д., Миллер С. (январь 1996 г.). «Сильно взаимодействующая пара остатков на поверхности контакта харибдотоксина и шейкерного K+-канала» . Нейрон . 16 (1): 123–130. дои : 10.1016/S0896-6273(00)80029-X . ПМИД 8562075 . S2CID 16794677 .
- ^ Ю М, Лю С.Л., Сунь П.Б., Пан Х., Тянь С.Л., Чжан Л.Х. (январь 2016 г.). «Пептидные токсины и низкомолекулярные блокаторы ВК-каналов» . Акта Фармакологика Синика . 37 (1): 56–66. дои : 10.1038/aps.2015.139 . ПМК 4722972 . ПМИД 26725735 .
- ^ Кандия С., Гарсия М.Л., Латорре Р. (август 1992 г.). «Способ действия ибериотоксина, мощного блокатора Ca (2+)-активируемого K+-канала с большой проводимостью» . Биофизический журнал . 63 (2): 583–590. Бибкод : 1992BpJ....63..583C . дои : 10.1016/S0006-3495(92)81630-2 . ПМЦ 1262182 . ПМИД 1384740 .
- ^ Стокер М., Краузе М., Педарзани П. (апрель 1999 г.). «Апамин-чувствительный Ca2+-активируемый ток K+ в пирамидных нейронах гиппокампа» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (8): 4662–4667. Бибкод : 1999PNAS...96.4662S . дои : 10.1073/pnas.96.8.4662 . ЧВК 16389 . ПМИД 10200319 .
- ^ Маклеод Дж. Ф., Лимпоэлс Дж. М., Пэн С. С., Дакс С. Л., Майерс Л. Дж., Голдер Ф. Дж. (ноябрь 2014 г.). «GAL-021, новый внутривенный блокатор BKCa-каналов, хорошо переносится и стимулирует вентиляцию легких у здоровых добровольцев» . Британский журнал анестезии . 113 (5): 875–883. дои : 10.1093/bja/aeu182 . ПМИД 24989775 .
- ^ Допико А.М., Букия А.Н., Кунтамаллаппанавар Г., Лю Дж. (2016). «Модуляция каналов БК этанолом» . Международное обозрение нейробиологии . 128 : 239–279. дои : 10.1016/bs.irn.2016.03.019 . ISBN 9780128036198 . ПМК 5257281 . ПМИД 27238266 .
- ^ Перейти обратно: а б Патнаик, Прадьот (2003). Справочник неорганических химикатов . МакГроу-Хилл. стр. 77–78 . ISBN 978-0-07-049439-8 .
- ^ Сакин Х., Син С., Палмер Л.Г., Чой Х., Уолтерс Д.Э. (февраль 2001 г.). «Регуляция РОМК внеклеточными катионами» . Биофизический журнал . 80 (2): 683–697. Бибкод : 2001BpJ....80..683S . дои : 10.1016/S0006-3495(01)76048-1 . ПМК 1301267 . ПМИД 11159436 .
- ^ Дхамун А.С., Джалифе Дж. (март 2005 г.). «Входящий выпрямительный ток (IK1) контролирует возбудимость сердца и участвует в аритмогенезе». Сердечный ритм . 2 (3): 316–324. дои : 10.1016/j.hrthm.2004.11.012 . ПМИД 15851327 .
- ^ Перейти обратно: а б с д и ж Свейл Д.Р., Хараде С.В., Дентон Дж.С. (апрель 2014 г.). «Фармакология внутренних выпрямителей калиевых каналов сердца и почек: новые инструменты интегративной физиологии и терапии» . Современное мнение в фармакологии . 15 :7–15. дои : 10.1016/j.coph.2013.11.002 . ПМК 4097192 . ПМИД 24721648 .
- ^ Ксиногалос П., Сейлер С., Шерер Д., Кеппл С., Шольц Е.П., Томас Д. и др. (декабрь 2014 г.). «Антиаритмический препарат III класса дронедарон ингибирует внутренние каналы сердца, выпрямляя каналы Kir2.1 посредством связывания по остатку E224». Архив фармакологии Наунина-Шмидеберга . 387 (12): 1153–1161. дои : 10.1007/s00210-014-1045-6 . ПМИД 25182566 . S2CID 10575229 .
- ^ Кеппл С., Шерер Д., Сейлер С., Шольц Э., Томас Д., Катус Х.А., Цитрон Е. (май 2017 г.). «Двойной механизм ингибирования внутренних выпрямляющихся каналов Kir2.x хинидином, включающий прямую блокировку пор и PIP 2 -интерференцию» . Журнал фармакологии и экспериментальной терапии . 361 (2): 209–218. дои : 10.1124/jpet.116.238287 . ПМИД 28188270 . S2CID 206502631 .
- ^ Кабальеро Р., Дольц-Гайтон П., Гомес Р., Аморос И., Барана А., Гонсалес де ла Фуэнте М. и др. (август 2010 г.). «Флекаинид увеличивает токи Kir2.1 за счет взаимодействия с цистеином 311, уменьшая индуцированное полиамином выпрямление» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (35): 15631–15636. Бибкод : 2010PNAS..10715631C . дои : 10.1073/pnas.1004021107 . ПМЦ 2932566 . ПМИД 20713726 .
- ^ Кобаяши Т., Васияма К., Икеда К. (март 2006 г.). «Ингибирование G-белка, активируемого внутренними выпрямляющими K+-каналами, ифенпродилом» . Нейропсихофармакология . 31 (3): 516–524. дои : 10.1038/sj.npp.1300844 . ПМИД 16123769 .
- ^ Соеда Ф., Фудзиеда Ю., Киносита М., Ширасаки Т., Такахама К. (май 2016 г.). «Ненаркотические противокашлевые средства центрального действия предотвращают гиперактивность у мышей: вовлечение каналов GIRK». Фармакология, биохимия и поведение . 144 : 26–32. дои : 10.1016/j.pbb.2016.02.006 . ПМИД 26892760 . S2CID 30118634 .
- ^ Ямамото Г., Соеда Ф., Ширасаки Т., Такахама К. (апрель 2011 г.). «[Является ли канал GIRK возможной мишенью при разработке нового терапевтического препарата для лечения нарушений мочеиспускания?]» . Якугаку Засси . 131 (4): 523–532. дои : 10.1248/yakushi.131.523 . ПМИД 21467791 .
- ^ Каваура К., Хонда С., Соеда Ф., Ширасаки Т., Такахама К. (май 2010 г.). «[Новое антидепрессантоподобное действие препаратов, блокирующих GIRK-каналы, у крыс]» . Якугаку Засси . 130 (5): 699–705. дои : 10.1248/yakushi.130.699 . ПМИД 20460867 .
- ^ Джин В., Лу З. (сентябрь 1998 г.). «Новый высокоаффинный ингибитор K+-каналов внутреннего выпрямителя». Биохимия . 37 (38): 13291–13299. дои : 10.1021/bi981178p . ПМИД 9748337 .
- ^ Каваура К., Огата Ю., Иноуэ М., Хонда С., Соэда Ф., Ширасаки Т., Такахама К. (декабрь 2009 г.). «Ненаркотическое противокашлевое средство центрального действия типепидин оказывает эффект, подобный антидепрессанту, в тесте принудительного плавания на крысах» (PDF) . Поведенческие исследования мозга . 205 (1): 315–318. дои : 10.1016/j.bbr.2009.07.004 . ПМИД 19616036 . S2CID 29236491 .
- ^ Кауфманн К., Ромейн И., Дэйс Е., Паскуаль С., Малик А., Ян Л. и др. (сентябрь 2013 г.). «ML297 (VU0456810), первый мощный и селективный активатор калиевого канала GIRK, проявляет противоэпилептические свойства у мышей» . ACS Химическая нейронаука . 4 (9): 1278–1286. дои : 10.1021/cn400062a . ПМЦ 3778424 . ПМИД 23730969 .
- ^ Серрано-Мартин X, Пайарес Г, Мендоса-Леон А (декабрь 2006 г.). «Глибенкламид, блокатор К+(АТФ)-каналов, проявляет антилейшманиальную активность при экспериментальном кожном лейшманиозе у мышей» . Антимикробные средства и химиотерапия . 50 (12): 4214–4216. дои : 10.1128/AAC.00617-06 . ПМК 1693980 . ПМИД 17015627 .
- ^ Лоуренс К.Л., Прокс П., Родриго Г.К., Джонс П., Хаябучи Ю., Стэнден Н.Б., Эшкрофт Ф.М. (август 2001 г.). «Гликлазид вызывает высокоаффинную блокировку КАТФ-каналов в изолированных бета-клетках поджелудочной железы мышей, но не в клетках сердца или гладкой мускулатуры артерий крысы» . Диабетология . 44 (8): 1019–1025. дои : 10.1007/s001250100595 . ПМИД 11484080 .
- ^ Перейти обратно: а б с д и ж Эньеди П., Чирьяк Г. (апрель 2010 г.). «Молекулярная основа токов утечки K+: двухпоровые доменные калиевые каналы» . Физиологические обзоры . 90 (2): 559–605. doi : 10.1152/physrev.00029.2009 . ПМИД 20393194 . S2CID 9358238 .
- ^ Перейти обратно: а б с д и ж Лотшоу Д.П. (2007). «Биофизические, фармакологические и функциональные характеристики клонированных и нативных K+-каналов с двухпоровым доменом млекопитающих». Клеточная биохимия и биофизика . 47 (2): 209–256. дои : 10.1007/s12013-007-0007-8 . ПМИД 17652773 . S2CID 12759521 .
- ^ Финк М., Лесаж Ф., Дюпра Ф., Эрто К., Рейес Р., Фоссет М., Лаздунски М. (июнь 1998 г.). «Нейрональный K+-канал с двумя P-доменами, стимулируемый арахидоновой кислотой и полиненасыщенными жирными кислотами» . Журнал ЭМБО . 17 (12): 3297–3308. дои : 10.1093/emboj/17.12.3297 . ПМК 1170668 . ПМИД 9628867 .
- ^ Гольдштейн С.А., Бокенхауэр Д., О'Келли И., Зильберберг Н. (март 2001 г.). «Каналы утечки калия и семейство субъединиц KCNK с двумя P-доменами» . Обзоры природы. Нейронаука . 2 (3): 175–184. дои : 10.1038/35058574 . ПМИД 11256078 . S2CID 9682396 .
- ^ Сано И., Инамура К., Мияке А., Мотидзуки С., Китада С., Ёкой Х. и др. (июль 2003 г.). «Новый двухпоровый домен К+-канала, TRESK, локализован в спинном мозге» . Журнал биологической химии . 278 (30): 27406–27412. дои : 10.1074/jbc.M206810200 . ПМИД 12754259 .
- ^ Чирьяк Г., Тот З.Е., Эньеди П. (апрель 2004 г.). «К+-канал с двухпоровым доменом, TRESK, активируется цитоплазматическим сигналом кальция через кальциневрин» . Журнал биологической химии . 279 (18): 18550–18558. дои : 10.1074/jbc.M312229200 . ПМИД 14981085 .
- ^ Kindler CH, Yost CS, Grey AT (апрель 1999 г.). «Местное анестезирующее ингибирование базовых калиевых каналов с двумя тандемными поровыми доменами» . Анестезиология . 90 (4): 1092–1102. дои : 10.1097/00000542-199904000-00024 . ПМИД 10201682 .
- ^ Перейти обратно: а б с Медоуз Х.Дж., Рэндалл А.Д. (март 2001 г.). «Функциональная характеристика человеческой TASK-3, кислоточувствительного двухпорового калиевого канала». Нейрофармакология . 40 (4): 551–559. дои : 10.1016/S0028-3908(00)00189-1 . ПМИД 11249964 . S2CID 20181576 .
- ^ Киндлер Ч., Пол М., Цзоу Х., Лю С., Уайнегар Б.Д., Грей А.Т., Йост CS (июль 2003 г.). «Амидные местные анестетики мощно ингибируют фоновый K+-канал TASK-2 домена тандемных пор человека (KCNK5)». Журнал фармакологии и экспериментальной терапии . 306 (1): 84–92. дои : 10.1124/jpet.103.049809 . ПМИД 12660311 . S2CID 1621972 .
- ^ Панке М.А., Личер Т., Понгс О., Фридрих П. (июнь 2003 г.). «Ингибирование каналов TREK-1 человека бупивакаином» . Анестезия и анальгезия . 96 (6): 1665–1673. дои : 10.1213/01.ANE.0000062524.90936.1F . ПМИД 12760993 . S2CID 39630495 .
- ^ Лесаж Ф., Гиймар Э., Финк М., Дюпра Ф., Лаздунски М., Роми Г., Барханин Дж. (март 1996 г.). «TWIK-1, вездесущий человек, слабо выпрямляющий K+-канал с новой структурой» . Журнал ЭМБО . 15 (5): 1004–1011. дои : 10.1002/j.1460-2075.1996.tb00437.x . ПМК 449995 . ПМИД 8605869 .
- ^ Дюпра Ф., Лесаж Ф., Финк М., Рейес Р., Эрто К., Лаздунски М. (сентябрь 1997 г.). «TASK, фоновый K+-канал человека для определения внешних изменений pH, близких к физиологическому pH» . Журнал ЭМБО . 16 (17): 5464–5471. дои : 10.1093/emboj/16.17.5464 . ПМК 1170177 . ПМИД 9312005 .
- ^ Рейес Р., Дюпра Ф., Лесаж Ф., Финк М., Салинас М., Фарман Н., Лаздунски М. (ноябрь 1998 г.). «Клонирование и экспрессия нового pH-чувствительного двухпорового домена K+-канала из почек человека» . Журнал биологической химии . 273 (47): 30863–30869. дои : 10.1074/jbc.273.47.30863 . ПМИД 9812978 .
- ^ Медоуз Х.Дж., Бенхем К.Д., Кэрнс В., Глогер И., Дженнингс С., Медхерст А.Д. и др. (апрель 2000 г.). «Клонирование, локализация и функциональная экспрессия человеческого ортолога калиевого канала TREK-1». Архив Пфлюгерса . 439 (6): 714–722. дои : 10.1007/s004240050997 . ПМИД 10784345 .
- ^ «UniProtKB — Q9NPC2 (KCNK9_HUMAN)» . Унипрот . Проверено 29 мая 2019 г.
- ^ Перейти обратно: а б Кеннард Л.Е., Чамбли-младший, Ранатунга К.М., Армстронг С.Дж., Вил Э.Л., Мэти А. (март 2005 г.). «Ингибирование калиевого канала двухпорового домена человека, TREK-1, флуоксетином и его метаболитом норфлуоксетином» . Британский журнал фармакологии . 144 (6): 821–829. дои : 10.1038/sj.bjp.0706068 . ПМК 1576064 . ПМИД 15685212 .
- ^ Патель А.Дж., Оноре Э., Лесаж Ф., Финк М., Роми Г., Лаздунски М. (май 1999 г.). «Ингаляционные анестетики активируют двухпоровые фоновые K+-каналы». Природная неврология . 2 (5): 422–426. дои : 10.1038/8084 . ПМИД 10321245 . S2CID 23092576 .
- ^ Грей А.Т., Чжао Б.Б., Киндлер CH, Уайнгар Б.Д., Мазурек М.Дж., Сюй Дж. и др. (июнь 2000 г.). «Летучие анестетики активируют базовый K+-канал KCNK5 домена тандемных пор человека». Анестезиология . 92 (6): 1722–1730. дои : 10.1097/00000542-200006000-00032 . ПМИД 10839924 . S2CID 45487917 .
- ^ Кирш Г.Е., Нарахаши Т. (июнь 1978 г.). «3,4-диаминопиридин. Новый мощный блокатор калиевых каналов» . Биофизический журнал . 22 (3): 507–512. Бибкод : 1978BpJ....22..507K . дои : 10.1016/s0006-3495(78)85503-9 . ПМЦ 1473482 . ПМИД 667299 .
- ^ Перейти обратно: а б Судья С.И., Бевер, Коннектикут (июль 2006 г.). «Блокаторы калиевых каналов при рассеянном склерозе: нейрональные Kv-каналы и эффекты симптоматического лечения». Фармакология и терапия . 111 (1): 224–259. doi : 10.1016/j.pharmthera.2005.10.006 . ПМИД 16472864 .
- ^ Тику П.Е., Новелл П.Т. (декабрь 1991 г.). «Селективное ингибирование К(+)-стимуляции Na,K-АТФазы бретилием» . Британский журнал фармакологии . 104 (4): 895–900. дои : 10.1111/j.1476-5381.1991.tb12523.x . ЧВК 1908819 . ПМИД 1667290 .
- ^ Хилле Б. (май 1967 г.). «Селективное ингибирование замедленных калиевых токов в нерве ионом тетраэтиламмония» . Журнал общей физиологии . 50 (5): 1287–1302. дои : 10.1085/jgp.50.5.1287 . ПМК 2225709 . ПМИД 6033586 .
- ^ Армстронг CM (октябрь 1971 г.). «Взаимодействие производных ионов тетраэтиламмония с калиевыми каналами гигантских аксонов» . Журнал общей физиологии . 58 (4): 413–437. дои : 10.1085/jgp.58.4.413 . ПМК 2226036 . ПМИД 5112659 .
- ^ «Амиодарон» . Аптечный банк . Проверено 28 мая 2019 г.
- ^ Рогавский М.А., Базиль К.В. (июль 2008 г.). «Новые молекулярные мишени для противоэпилептических препаратов: альфа (2) дельта, SV2A и калиевые каналы K (v) 7 / KCNQ / M» . Текущие отчеты по неврологии и нейробиологии . 8 (4): 345–352. дои : 10.1007/s11910-008-0053-7 . ПМК 2587091 . ПМИД 18590620 .
- ^ Перейти обратно: а б Дойл Д.А., Мораис Кабрал Дж., Пфюцнер Р.А., Куо А., Гулбис Дж.М., Коэн С.Л. и др. (апрель 1998 г.). «Строение калиевого канала: молекулярные основы К+-проводимости и селективности». Наука . 280 (5360): 69–77. Бибкод : 1998Sci...280...69D . дои : 10.1126/science.280.5360.69 . ПМИД 9525859 .
- ^ Маккиннон Р., Коэн С.Л., Куо А., Ли А., Чайт Б.Т. (апрель 1998 г.). «Структурная консервация в прокариотических и эукариотических калиевых каналах». Наука . 280 (5360): 106–109. Бибкод : 1998Sci...280..106M . дои : 10.1126/science.280.5360.106 . ПМИД 9525854 . S2CID 33907550 .
- ^ Армстронг С. (апрель 1998 г.). «Видение поры». Наука . 280 (5360): 56–57. дои : 10.1126/science.280.5360.56 . ПМИД 9556453 . S2CID 35339674 .
- ^ «Нобелевская премия по химии 2003 г.» . Нобелевский фонд . Проверено 16 ноября 2007 г.
- ^ Чжоу Ю, Мораис-Кабрал Дж. Х., Кауфман А., Маккиннон Р. (ноябрь 2001 г.). «Химия координации и гидратации ионов, выявленная комплексом K + канал-Fab при разрешении 2,0 А». Природа . 414 (6859): 43–48. Бибкод : 2001Natur.414...43Z . дои : 10.1038/35102009 . ПМИД 11689936 . S2CID 205022645 .
- ^ Лодиш Х., Берк А., Кайзер С., Кригер М., Бретшер А., Плох Х. и др. (2016). Молекулярно-клеточная биология (8-е изд.). Нью-Йорк, штат Нью-Йорк: WH Freeman and Company. п. 499. ИСБН 978-1-4641-8339-3 .
- ^ Зауэр Д.Б., Цзэн В., Рагунатан С., Цзян Юй (октябрь 2011 г.). «Взаимодействия с белками играют центральную роль в стабилизации фильтра селективности каналов K+ в четырехсайтовой конфигурации для селективного проникновения K+» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (40): 16634–16639. Бибкод : 2011PNAS..10816634S . дои : 10.1073/pnas.1111688108 . ПМК 3189067 . ПМИД 21933962 .
- ^ Перейти обратно: а б Хеллгрен М., Сандберг Л., Эдхольм О. (март 2006 г.). «Сравнение двух прокариотических калиевых каналов (KirBac1.1 и KcsA) в исследовании моделирования молекулярной динамики (MD)». Биофизическая химия . 120 (1): 1–9. дои : 10.1016/j.bpc.2005.10.002 . ПМИД 16253415 .
- ^ Носков С.Ю., Ру Б. (февраль 2007 г.). «Важность гидратации и динамики для селективности каналов KcsA и NaK» . Журнал общей физиологии . 129 (2): 135–143. дои : 10.1085/jgp.200609633 . ПМК 2154357 . ПМИД 17227917 .
- ^ Носков С.Ю., Бернеш С., Ру Б. (октябрь 2004 г.). «Контроль селективности ионов в калиевых каналах с помощью электростатических и динамических свойств карбонильных лигандов». Природа . 431 (7010): 830–834. Бибкод : 2004Natur.431..830N . дои : 10.1038/nature02943 . ПМИД 15483608 . S2CID 4414885 .
- ^ Варма С., Ремпе С.Б. (август 2007 г.). «Настройка архитектуры координации ионов для обеспечения избирательного разделения» . Биофизический журнал . 93 (4): 1093–1099. arXiv : физика/0608180 . Бибкод : 2007BpJ....93.1093V . дои : 10.1529/biophysj.107.107482 . ЧВК 1929028 . ПМИД 17513348 .
- ^ Томас М., Джаятилака Д., Корри Б. (октябрь 2007 г.). «Преобладающая роль координационного числа в селективности калиевых каналов» . Биофизический журнал . 93 (8): 2635–2643. Бибкод : 2007BpJ....93.2635T . дои : 10.1529/biophysj.107.108167 . ЧВК 1989715 . ПМИД 17573427 .
- ^ Бостик Д.Л., Брукс К.Л. (май 2007 г.). «Селективность в K+-каналах обусловлена топологическим контролем координированного состояния проникающих ионов» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (22): 9260–9265. Бибкод : 2007PNAS..104.9260B . дои : 10.1073/pnas.0700554104 . ПМК 1890482 . ПМИД 17519335 .
- ^ Деребе М.Г., Зауэр Д.Б., Цзэн В., Алам А., Ши Н., Цзян Й. (январь 2011 г.). «Настройка ионной селективности каналов тетрамерных катионов путем изменения количества мест связывания ионов» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (2): 598–602. Бибкод : 2011PNAS..108..598D . дои : 10.1073/pnas.1013636108 . ПМК 3021048 . ПМИД 21187421 .
- ^ Мораис-Кабрал Дж. Х., Чжоу Ю., Маккиннон Р. (ноябрь 2001 г.). «Энергетическая оптимизация скорости ионной проводимости с помощью селективного фильтра K +». Природа . 414 (6859): 37–42. Бибкод : 2001Natur.414...37M . дои : 10.1038/35102000 . ПМИД 11689935 . S2CID 4429890 .
- ^ Перейти обратно: а б Юань П., Леонетти, доктор медицинских наук, Пико А.Р., Сюн Ю, Маккиннон Р. (июль 2010 г.). «Структура аппарата активации Са2+-канала ВК человека при разрешении 3,0 А» . Наука . 329 (5988): 182–186. Бибкод : 2010Sci...329..182Y . дои : 10.1126/science.1190414 . ПМК 3022345 . ПМИД 20508092 .
- ^ Перейти обратно: а б У Ю, Ян Ю, Е С, Цзян Ю (июль 2010 г.). «Структура управляющего кольца человеческого Ca(2+)-закрытого K(+)-канала с большой проводимостью» . Природа . 466 (7304): 393–397. Бибкод : 2010Natur.466..393W . дои : 10.1038/nature09252 . ПМЦ 2910425 . ПМИД 20574420 .
- ^ Перейти обратно: а б Цзян Ю, Пико А., Каден М., Чайт Б.Т., Маккиннон Р. (март 2001 г.). «Структура домена RCK из K+-канала E. coli и демонстрация его присутствия в канале BK человека» . Нейрон . 29 (3): 593–601. дои : 10.1016/S0896-6273(01)00236-7 . ПМИД 11301020 . S2CID 17880955 .
- ^ Цзян Ю, Ли А., Чен Дж., Каден М., Чайт Б.Т., Маккиннон Р. (май 2002 г.). «Кристаллическая структура и механизм кальций-управляемого калиевого канала». Природа . 417 (6888): 515–522. Бибкод : 2002Natur.417..515J . дои : 10.1038/417515a . ПМИД 12037559 . S2CID 205029269 .
- ^ Конг С., Цзэн В., Йе С., Чен Л., Зауэр Д.Б., Лам Ю. и др. (декабрь 2012 г.). «Различные механизмы шлюзования, выявленные структурами мультилигандного закрытого канала K (+)» . электронная жизнь . 1 : е00184. doi : 10.7554/eLife.00184 . ПМК 3510474 . ПМИД 23240087 .
- ^ Цао Ю, Цзинь Х, Хуан Х, Деребе М.Г., Левин Э.Дж., Кабалиесваран В. и др. (март 2011 г.). «Кристаллическая структура переносчика ионов калия ТркХ» . Природа . 471 (7338): 336–340. Бибкод : 2011Natur.471..336C . дои : 10.1038/nature09731 . ПМК 3077569 . ПМИД 21317882 .
- ^ Уйсал С., Куэлло Л.Г., Кортес Д.М., Коиде С., Косьяков А.А., Перозо Е. (июль 2011 г.). «Механизм активационного стробирования в полноразмерном канале KcsA K+» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (29): 11896–11899. Бибкод : 2011PNAS..10811896U . дои : 10.1073/pnas.1105112108 . ПМК 3141920 . ПМИД 21730186 .
- ^ Клейтон Г.М., Сильверман В.Р., Хегинботам Л., Мораис-Кабрал Дж.Х. (ноябрь 2004 г.). «Структурные основы активации лиганда в калиевом канале, регулируемом циклическими нуклеотидами» . Клетка . 119 (5): 615–627. дои : 10.1016/j.cell.2004.10.030 . ПМИД 15550244 . S2CID 14149494 .
- ^ Цзян Ю, Ли А, Чен Дж, Рута В, Каден М, Чайт БТ, Маккиннон Р (май 2003 г.). «Рентгеновская структура потенциалзависимого K+-канала». Природа . 423 (6935): 33–41. Бибкод : 2003Natur.423...33J . дои : 10.1038/nature01580 . ПМИД 12721618 . S2CID 4347957 .
- ^ Лонг С.Б., Кэмпбелл Э.Б., Маккиннон Р. (август 2005 г.). «Кристаллическая структура потенциал-зависимого K+-канала семейства шейкеров млекопитающих» . Наука . 309 (5736): 897–903. Бибкод : 2005Sci...309..897L . дои : 10.1126/science.1116269 . ПМИД 16002581 . S2CID 6072007 .
- ^ Антц С., Факлер Б. (август 1998 г.). «Быстрая инактивация потенциалзависимых K (+)-каналов: от мультфильма к структуре». Новости физиологических наук . 13 (4): 177–182. doi : 10.1152/физиологияонлайн.1998.13.4.177 . ПМИД 11390785 . S2CID 38134756 .
- ^ Ченг В.В., Маккой Дж.Г., Томпсон А.Н., Николс К.Г. , Нимигей К.М. (март 2011 г.). «Механизм селективно-инактивационного взаимодействия в калиевых каналах KcsA» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (13): 5272–5277. Бибкод : 2011PNAS..108.5272C . дои : 10.1073/pnas.1014186108 . ПМК 3069191 . ПМИД 21402935 .
- ^ Куэлло Л.Г., Джогини В., Кортес Д.М., Перозо Э. (июль 2010 г.). «Структурный механизм инактивации С-типа в К(+) каналах» . Природа . 466 (7303): 203–208. Бибкод : 2010Natur.466..203C . дои : 10.1038/nature09153 . ПМК 3033749 . ПМИД 20613835 .
- ^ Куэлло Л.Г., Джогини В., Кортес Д.М., Пан А.С., Ганьон Д.Г., Далмас О. и др. (июль 2010 г.). «Структурная основа связи между воротами активации и инактивации в каналах K (+)» . Природа . 466 (7303): 272–275. Бибкод : 2010Natur.466..272C . дои : 10.1038/nature09136 . ПМК 3033755 . ПМИД 20613845 .
- ^ Лужков В.Б., Аквист Ю. (февраль 2005 г.). «Ионы и блокаторы в калиевых каналах: результаты моделирования свободной энергии». Biochimica et Biophysical Acta (BBA) - Белки и протеомика . 1747 (1): 109–120. дои : 10.1016/j.bbapap.2004.10.006 . ПМИД 15680245 .
- ^ Лужков В.Б., Остерберг Ф., Аквист Дж. (ноябрь 2003 г.). «Взаимосвязь структура-активность внеклеточной блокады K+-каналов ионами тетраалкиламмония» . Письма ФЭБС . 554 (1–2): 159–164. Бибкод : 2003FEBSL.554..159L . дои : 10.1016/S0014-5793(03)01117-7 . ПМИД 14596932 . S2CID 32031835 .
- ^ Posson DJ, McCoy JG, Nimigean CM (февраль 2013 г.). «Затвор, зависимый от напряжения в калиевых каналах MthK, расположен на селективном фильтре» . Структурная и молекулярная биология природы . 20 (2): 159–166. дои : 10.1038/nsmb.2473 . ПМК 3565016 . ПМИД 23262489 .
- ^ Чой К.Л., Моссман С., Обе Дж., Йеллен Дж. (март 1993 г.). «Внутренний рецептор четвертичного аммония калиевых каналов Шейкера». Нейрон . 10 (3): 533–541. дои : 10.1016/0896-6273(93)90340-w . ПМИД 8461140 . S2CID 33361945 .
- ^ Пиаста К.Н., Теобальд Д.Л., Миллер С. (октябрь 2011 г.). «Калий-селективная блокировка проникновения бария через одиночные каналы KcsA» . Журнал общей физиологии . 138 (4): 421–436. дои : 10.1085/jgp.201110684 . ПМК 3182450 . ПМИД 21911483 .
- ^ Нейтон Дж., Миллер С. (ноябрь 1988 г.). «Калий блокирует проникновение бария через активируемые кальцием калиевые каналы» . Журнал общей физиологии . 92 (5): 549–567. дои : 10.1085/jgp.92.5.549 . ПМК 2228918 . ПМИД 3235973 .
- ^ Lockless SW, Чжоу М., Маккиннон Р. (май 2007 г.). «Структурные и термодинамические свойства селективного связывания ионов в К+-канале» . ПЛОС Биология . 5 (5): е121. дои : 10.1371/journal.pbio.0050121 . ПМК 1858713 . ПМИД 17472437 .
- ^ Цзян Ю, Маккиннон Р. (март 2000 г.). «Участок бария в калиевом канале по данным рентгеновской кристаллографии» . Журнал общей физиологии . 115 (3): 269–272. дои : 10.1085/jgp.115.3.269 . ПМК 2217209 . ПМИД 10694255 .
- ^ Лам Ю.Л., Цзэн В., Зауэр Д.Б., Цзян Ю. (август 2014 г.). «Фильтр консервативного калиевого канала может иметь различные профили связывания ионов: структурный анализ связывания рубидия, цезия и бария в NaK2K» . Журнал общей физиологии . 144 (2): 181–192. дои : 10.1085/jgp.201411191 . ПМЦ 4113894 . ПМИД 25024267 .
- ^ Го Р, Цзэн В, Цуй Х, Чен Л, Е С (август 2014 г.). «Ионные взаимодействия блокад Ba2+ в канале MthK K+» . Журнал общей физиологии . 144 (2): 193–200. дои : 10.1085/jgp.201411192 . ПМЦ 4113901 . ПМИД 25024268 .
- ^ Крапивинский Г., Гордон Э.А., Викман К., Велимирович Б., Крапивинский Л., Клэпхэм Д.Е. (март 1995 г.). «G-белок-управляемый предсердный K+-канал IKACh представляет собой гетеромультимер двух белков K(+)-канала, выпрямляющих внутрь». Природа . 374 (6518): 135–141. Бибкод : 1995Natur.374..135K . дои : 10.1038/374135a0 . ПМИД 7877685 . S2CID 4334467 .
- ^ Кори С., Крапивинский Г., Крапивинский Л., Клэпхэм Д.Е. (февраль 1998 г.). «Количество и стехиометрия субъединиц в нативном предсердном K+-канале, управляемом G-белком, IKACh» . Журнал биологической химии . 273 (9): 5271–5278. дои : 10.1074/jbc.273.9.5271 . ПМИД 9478984 .
- ^ Кункель М.Т., Перальта Э.Г. (ноябрь 1995 г.). «Идентификация доменов, обеспечивающих регуляцию G-белка на внутренних калиевых каналах выпрямителя» . Клетка . 83 (3): 443–449. дои : 10.1016/0092-8674(95)90122-1 . ПМИД 8521474 . S2CID 14720432 .
- ^ Викман К., Крапивинский Г., Кори С., Кеннеди М., Немец Дж., Медина И., Клэпхэм Д.Э. (апрель 1999 г.). «Структура, активация G-белка и функциональная значимость сердечного K+-канала, управляемого G-белком, IKACh» . Анналы Нью-Йоркской академии наук . 868 (1): 386–398. Бибкод : 1999NYASA.868..386W . дои : 10.1111/j.1749-6632.1999.tb11300.x . ПМИД 10414308 . S2CID 25949938 . Архивировано из оригинала 29 января 2006 г.
- ^ Болл П. (март 2008 г.). «Тигель: Искусство, вдохновленное наукой, должно быть больше, чем просто красивая картинка» . Химический мир . 5 (3): 42–43 . Проверено 12 января 2009 г.
Внешние ссылки
[ редактировать ]- Канал Протеопедия Калиевый канал в 3D
- Калий + каналы Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- Центр нервно-мышечных заболеваний (04 марта 2008 г.). «Калиевые каналы» . Вашингтонский университет в Сент-Луисе . Проверено 10 марта 2008 г.