Ген
| Часть серии о |
| Генетика |
|---|
 |

В биологии слово ген имеет два значения. Менделевский ген является основной единицей наследственности . Молекулярный ген представляет собой последовательность нуклеотидов в ДНК , которая транскрибируется с образованием функциональной РНК . Существует два типа молекулярных генов: гены, кодирующие белки, и некодирующие гены. [1] [2] [3]
Во время экспрессии гена (синтеза РНК или белка из гена) ДНК сначала копируется в РНК . РНК может быть непосредственно функциональной или быть промежуточной матрицей для синтеза белка.
организма Передача генов потомству является основой наследования фенотипических признаков от одного поколения к другому. гены составляют различные последовательности ДНК, называемые генотипом , которые специфичны для каждого конкретного человека в генофонде популяции Эти данного вида . Генотип, наряду с факторами окружающей среды и развития, в конечном итоге определяет фенотип человека.
Большинство биологических признаков возникает под совместным влиянием полигенов (набора различных генов) и взаимодействий генов и окружающей среды . Некоторые генетические черты видны мгновенно, например цвет глаз или количество конечностей, другие нет, например группа крови , риск определенных заболеваний или тысячи основных биохимических процессов, составляющих жизнь .
Ген может приобретать мутации в своей последовательности различных вариантов, известных как аллели , что приводит к появлению в популяции . Эти аллели кодируют несколько разные версии гена, что может вызывать разные фенотипические признаки. [4] Гены развиваются в результате естественного отбора или выживания наиболее приспособленных и генетического дрейфа аллелей.
Термин «ген» был введен датским ботаником, физиологом растений и генетиком Вильгельмом Йохансеном в 1909 году. [5] Оно вдохновлено древнегреческим : γόνος, gonos , что означает потомство и продолжение рода.
Определения [ править ]
Существует много разных способов использования термина «ген», основанных на различных аспектах его наследования, отбора, биологической функции или молекулярной структуры, но большинство этих определений делятся на две категории: менделевский ген или молекулярный ген. [1] [6] [7] [8] [9]
Менделевский ген является классическим геном генетики и относится к любому наследственному признаку. Этот ген описан в книге «Эгоистичный ген» . [10] Более подробное обсуждение этой версии гена можно найти в статьях Генетика и Геноцентрированный взгляд на эволюцию .
Определение молекулярного гена чаще используется в биохимии, молекулярной биологии и большей части генетики — ген, который описывается с точки зрения последовательности ДНК. [1] Существует множество различных определений этого гена, некоторые из которых вводят в заблуждение или неверны. [6] [11]
Очень ранние работы в области молекулярной генетики предложили концепцию, согласно которой один ген производит один белок (первоначально «один ген — один фермент»). [12] [13] Однако гены, продуцирующие репрессорные РНК, были предложены в 1950-х годах. [14] а к 1960-м годам в учебниках использовались определения молекулярных генов, в том числе определения функциональных молекул РНК, таких как рибосомальная РНК и тРНК (некодирующие гены), а также гены, кодирующие белки. [15]
Идея о двух видах генов до сих пор является частью определения гена в большинстве учебников. Например,
Основная функция генома — производство молекул РНК. Выбранные части нуклеотидной последовательности ДНК копируются в соответствующую нуклеотидную последовательность РНК, которая либо кодирует белок (если это мРНК), либо образует «структурную» РНК, такую как транспортная РНК (тРНК) или рибосомальная РНК (рРНК). молекула. Каждый участок спирали ДНК, образующий функциональную молекулу РНК, представляет собой ген. [16]
Мы определяем ген как последовательность ДНК, которая транскрибируется. В это определение входят гены, которые не кодируют белки (не все транскрипты являются информационной РНК). Определение обычно исключает области генома, которые контролируют транскрипцию, но сами не транскрибируются. Мы столкнемся с некоторыми исключениями из нашего определения гена – как ни удивительно, не существует определения, которое было бы полностью удовлетворительным. [17]
Ген — это последовательность ДНК, которая кодирует диффундирующий продукт. Этот продукт может быть белком (как в большинстве генов) или РНК (как в случае генов, кодирующих тРНК и рРНК). Важнейшей особенностью является то, что продукт диффундирует из места синтеза, чтобы действовать в другом месте. [18]
Важными частями таких определений являются: (1) ген соответствует единице транскрипции; (2) гены продуцируют как мРНК, так и некодирующие РНК; и (3) регуляторные последовательности контролируют экспрессию гена, но не являются частью самого гена. Однако есть еще одна важная часть определения, и она подчеркивается в книге Костаса Кампуракиса « Осмысление генов» .
Поэтому в этой книге я буду рассматривать гены как последовательности ДНК, кодирующие информацию для функциональных продуктов, будь то белки или молекулы РНК. Под «кодированием информации» я имею в виду, что последовательность ДНК используется в качестве матрицы для производства молекулы РНК или белка, выполняющего некоторую функцию. [6]
Акцент на функции имеет важное значение, поскольку существуют участки ДНК, которые производят нефункциональные транскрипты, и их нельзя назвать генами. К ним относятся очевидные примеры, такие как транскрибируемые псевдогены, а также менее очевидные примеры, такие как мусорная РНК, образующаяся в виде шума из-за ошибок транскрипции. Согласно этому определению, чтобы квалифицироваться как настоящий ген, необходимо доказать, что транскрипт выполняет биологическую функцию. [6]
Ранние предположения о размере типичного гена основывались на генетическом картировании с высоким разрешением, а также на размере белков и молекул РНК. В то время (1965 г.) длина в 1500 пар оснований казалась разумной. [15] Это было основано на идее, что ген — это ДНК, которая непосредственно отвечает за производство функционального продукта. Открытие интронов в 1970-х годах означало, что многие эукариотические гены были намного больше, чем можно было бы предположить по размеру функционального продукта. Например, типичные гены, кодирующие белки млекопитающих, имеют длину около 62 000 пар оснований (транскрибируемая область), и, поскольку их около 20 000, они занимают около 35–40% генома млекопитающих (включая геном человека). [19] [20] [21]
Несмотря на то, что как гены, кодирующие белок, так и некодирующие гены известны уже более 50 лет, до сих пор существует ряд учебников, веб-сайтов и научных публикаций, которые определяют ген как последовательность ДНК, определяющую белок. Другими словами, определение ограничивается генами, кодирующими белки. Вот пример из недавней статьи в журнале American Scientist.
... чтобы по-настоящему оценить потенциальное значение генов de novo, мы опирались на строгое определение слова «ген», с которым может согласиться почти каждый эксперт. Во-первых, чтобы нуклеотидную последовательность можно было считать истинным геном, должна присутствовать открытая рамка считывания (ORF). ORF можно рассматривать как «сам ген»; он начинается со стартовой отметки, общей для каждого гена, и заканчивается одним из трех возможных сигналов финиша. Один из ключевых ферментов в этом процессе, РНК-полимераза, мчится по цепи ДНК, как поезд по монорельсовой дороге, транскрибируя ее в форму информационной РНК. Этот момент подводит нас ко второму важному критерию: настоящий ген — это тот, который одновременно транскрибируется и транслируется. То есть настоящий ген сначала используется в качестве матрицы для создания временной информационной РНК, которая затем транслируется в белок. [22]
Это ограниченное определение настолько распространено, что в последнее время оно породило множество статей, критикующих это «стандартное определение» и призывающих к новому расширенному определению, включающему некодирующие гены. [23] [24] [25] Однако это так называемое «новое» определение существует уже более полувека, и непонятно, почему некоторые современные авторы игнорируют некодирующие гены. [ редакция ]
Хотя некоторые определения могут быть применимы более широко, чем другие, фундаментальная сложность биологии означает, что ни одно определение гена не может полностью охватить все аспекты. Не все геномы представляют собой ДНК (например, РНК-вирусы ). [26] Бактериальные опероны представляют собой несколько областей, кодирующих белок, транскрибируемых в отдельные большие мРНК, альтернативный сплайсинг позволяет одной геномной области кодировать несколько продуктов района, а транс-сплайсинг объединяет мРНК из более короткой кодирующей последовательности по всему геному. [27] [28] [29] Поскольку молекулярные определения исключают такие элементы, как интроны, промоторы и другие регуляторные области , вместо этого они считаются «связанными» с геном и влияют на его функцию.
Иногда используется еще более широкое рабочее определение, чтобы охватить сложность этих разнообразных явлений, где ген определяется как объединение геномных последовательностей, кодирующих согласованный набор потенциально перекрывающихся функциональных продуктов. [30] Это определение классифицирует гены по их функциональным продуктам (белкам или РНК), а не по их конкретным локусам ДНК, при этом регуляторные элементы классифицируются как области , связанные с генами . [30]
История [ править ]
Открытие дискретных единиц наследственных

Существование дискретных наследуемых единиц было впервые предположено Грегором Менделем (1822–1884). [31] С 1857 по 1864 год в Брно , Австрийская империя (сегодняшняя Чехия), он изучал закономерности наследования у 8000 обыкновенных съедобных растений гороха , отслеживая различные признаки от родителя к потомству. Он описал их математически как 2 н комбинации, где n — количество различающихся характеристик исходного гороха. Хотя он не использовал термин «ген» , он объяснил свои результаты с точки зрения дискретных унаследованных единиц, которые порождают наблюдаемые физические характеристики. Это описание стало прообразом Вильгельма Йохансена различия между генотипом (генетическим материалом организма) и фенотипом (наблюдаемыми признаками этого организма). Мендель также был первым, кто продемонстрировал независимый ассортимент , различие между доминантными и рецессивными признаками, различие между гетерозиготой и гомозиготой , а также феномен прерывистого наследования.
До работы Менделя доминирующей теорией наследственности была теория смешанного наследования . [32] это предположило, что каждый родитель вносил свой вклад в процесс оплодотворения и что черты родителей смешивались и смешивались, чтобы произвести потомство. Чарльз Дарвин разработал теорию наследственности, которую он назвал пангенезисом , от греческого pan («все, целое») и генезиса («рождение»)/genos («происхождение»). [33] [34] Дарвин использовал термин «геммула» для описания гипотетических частиц, которые смешивались во время размножения.
Работа Менделя осталась практически незамеченной после ее первой публикации в 1866 году, но была вновь открыта в конце 19 века Хьюго де Фрисом , Карлом Корренсом и Эрихом фон Чермаком , которые (утверждали, что пришли) пришли к аналогичным выводам в своих собственных исследованиях. [35] В частности, в 1889 году Гюго де Врис опубликовал свою книгу « Внутриклеточный пангенезис» , [36] в котором он постулировал, что разные признаки имеют индивидуальных наследственных носителей и что наследование определенных признаков у организмов происходит в виде частиц. Де Врис назвал эти единицы «пангенами» ( Pangens на немецком языке) в честь теории пангенезиса Дарвина 1868 года.
Двадцать лет спустя, в 1909 году, Вильгельм Йохансен ввел термин «ген». [5] а в 1906 году Уильям Бейтсон — по « генетике ». [37] [30] в то время как Эдуард Страсбургер , среди других, все еще использовал термин «панген» для фундаментальной физической и функциональной единицы наследственности. [36] : Предисловие переводчика, viii
Открытие ДНК [ править ]
Прогресс в понимании генов и наследственности продолжался на протяжении всего 20 века. дезоксирибонуклеиновая кислота (ДНК) является молекулярным хранилищем генетической информации. Эксперименты 1940-1950-х годов показали, что [38] [39] Структура ДНК была изучена Розалиндой Франклин и Морисом Уилкинсом с помощью рентгеновской кристаллографии , что побудило Джеймса Д. Уотсона и Фрэнсиса Крика опубликовать модель двухцепочечной молекулы ДНК, спаренные нуклеотидные основания которой указывают на убедительную гипотезу механизма генетическая репликация. [40] [41]
В начале 1950-х годов преобладала точка зрения, что гены в хромосоме действуют как отдельные объекты, расположенные как бусины на нитке. Эксперименты Бензера с использованием мутантов , дефектных по области rII бактериофага Т4 (1955–1959), показали, что отдельные гены имеют простую линейную структуру и, вероятно, эквивалентны линейному участку ДНК. [42] [43]
В совокупности эти исследования установили центральную догму молекулярной биологии , которая гласит, что белки транслируются с РНК , которая транскрибируется с ДНК . С тех пор было показано, что из этой догмы есть исключения, такие как транскрипция ретровирусов обратная . Современное исследование генетики на уровне ДНК известно как молекулярная генетика .
В 1972 году Уолтер Фирс и его команда первыми определили последовательность гена: последовательность белка оболочки бактериофага MS2 . [44] Последующая разработка с обрывом цепи секвенирования ДНК в 1977 году Фредериком Сэнгером повысила эффективность секвенирования и превратила его в рутинный лабораторный инструмент. [45] Автоматизированная версия метода Сэнгера использовалась на ранних этапах проекта «Геном человека» . [46]
Современный синтез и его преемники [ править ]
Теории, разработанные в начале 20-го века для объединения менделевской генетики с дарвиновской эволюцией, называются современным синтезом — термин, введенный Джулианом Хаксли . [47]
Этот взгляд на эволюцию был подчеркнут Джорджа К. Уильямса геноцентричным взглядом на эволюцию . Он предположил, что менделевский ген является единицей естественного отбора с определением: «то, что разделяется и рекомбинируется с заметной частотой». [48] : 24 Связанные с этим идеи, подчеркивающие центральную роль менделевских генов и важность естественного отбора в эволюции, были популяризированы Ричардом Докинсом . [10] [49]
Развитие нейтральной теории эволюции в конце 1960-х годов привело к признанию того, что случайный генетический дрейф является основным фактором эволюции и что нейтральная теория должна быть нулевой гипотезой молекулярной эволюции. [50] Это привело к построению филогенетических деревьев и разработке молекулярных часов , которые лежат в основе всех методов датирования с использованием последовательностей ДНК. Эти методы не ограничиваются молекулярными последовательностями генов, но могут использоваться на всех сегментах ДНК генома.
Молекулярная основа [ править ]

ДНК [ править ]
Подавляющее большинство организмов кодируют свои гены в длинных цепях ДНК (дезоксирибонуклеиновая кислота). ДНК состоит из цепи , состоящей из четырех типов нуклеотидных субъединиц, каждая из которых состоит из пятиуглеродного сахара ( 2-дезоксирибозы ), фосфатной группы и одного из четырех оснований аденина , цитозина , гуанина и тимина . [51] : 2.1
Две цепи ДНК закручиваются друг вокруг друга, образуя двойную спираль ДНК , в которой фосфатно-сахарный остов закручивается по спирали снаружи, а основания направлены внутрь, при этом основания аденина соединяются с тимином, а основания гуанина с цитозином. Специфичность спаривания оснований возникает потому, что аденин и тимин образуют две водородные связи , тогда как цитозин и гуанин образуют три водородные связи. Таким образом, две цепи двойной спирали должны быть комплементарными , а их последовательность оснований совпадать, так что аденины одной цепи соединяются с тиминами другой цепи и так далее. [51] : 4.1
Благодаря химическому составу пентозных остатков оснований нити ДНК обладают направленностью. Один конец полимера ДНК содержит открытую гидроксильную группу дезоксирибозы ; это известно как 3'-конец молекулы. Другой конец содержит открытую фосфатную группу; это 5'-конец . Две нити двойной спирали идут в противоположных направлениях. Синтез нуклеиновой кислоты, включая ДНК репликацию и транскрипцию , происходит в направлении 5'→3', поскольку новые нуклеотиды добавляются посредством реакции дегидратации , в которой обнаженный 3'-гидроксил используется в качестве нуклеофила . [52] : 27.2
Экспрессия транскрипции генов, закодированных в ДНК, начинается с гена в РНК , второй тип нуклеиновой кислоты, очень похожий на ДНК, но мономеры которого содержат сахарную рибозу, а не дезоксирибозу . РНК также содержит основание урацил вместо тимина . Молекулы РНК менее стабильны, чем ДНК, и обычно являются одноцепочечными. Гены, кодирующие белки, состоят из серии трехнуклеотидных последовательностей , называемых кодонами , которые служат «словами» на генетическом «языке». Генетический код определяет соответствие при трансляции белка между кодонами и аминокислотами . Генетический код практически одинаков для всех известных организмов. [51] : 4.1
Хромосомы [ править ]


Полный набор генов в организме или клетке известен как его геном , который может храниться в одной или нескольких хромосомах . Хромосома состоит из одной очень длинной спирали ДНК, на которой закодированы тысячи генов. [51] : 4.2 Область хромосомы, в которой расположен конкретный ген, называется ее локусом . Каждый локус содержит одну аллель гена; однако члены популяции могут иметь разные аллели в этом локусе, каждый со слегка отличающейся последовательностью генов.
Большинство эукариотических генов хранятся в наборе крупных линейных хромосом. Хромосомы упакованы внутри ядра в комплекс с запасными белками, называемыми гистонами, образуя единицу, называемую нуклеосомой . ДНК, упакованная и конденсированная таким образом, называется хроматином . [51] : 4.2 Способ хранения ДНК в гистонах, а также химические модификации самого гистона регулируют, доступен ли конкретный участок ДНК для экспрессии генов . Помимо генов, хромосомы эукариот содержат последовательности, обеспечивающие копирование ДНК без деградации концевых участков и сортировку на дочерние клетки во время клеточного деления: точки начала репликации , теломеры и центромеры . [51] : 4.2 Истоки репликации — это области последовательности, в которых инициируется репликация ДНК для создания двух копий хромосомы. Теломеры представляют собой длинные участки повторяющихся последовательностей, которые замыкают концы линейных хромосом и предотвращают деградацию кодирующих и регуляторных областей во время репликации ДНК . Длина теломер уменьшается каждый раз, когда геном реплицируется, и это участвует в процессе старения . [54] Центромера необходима для связывания волокон веретена и разделения сестринских хроматид на дочерние клетки во время клеточного деления . [51] : 18.2
Прокариоты ( бактерии и археи ) обычно хранят свои геномы в одной большой кольцевой хромосоме . Точно так же некоторые эукариотические органеллы содержат оставшуюся кольцевую хромосому с небольшим количеством генов. [51] : 14.4 Прокариоты иногда дополняют свою хромосому дополнительными небольшими кольцами ДНК, называемыми плазмидами , которые обычно кодируют лишь несколько генов и могут передаваться от одного человека к другому. Например, гены устойчивости к антибиотикам обычно кодируются бактериальными плазмидами и могут передаваться между отдельными клетками, даже принадлежащими к разным видам, посредством горизонтального переноса генов . [55]
В то время как хромосомы прокариот относительно богаты генами, хромосомы эукариот часто содержат участки ДНК, которые не выполняют никакой очевидной функции. Простые одноклеточные эукариоты имеют относительно небольшое количество такой ДНК, тогда как геномы сложных многоклеточных организмов , включая человека, содержат абсолютное большинство ДНК без выявленной функции. [56] Эту ДНК часто называют « мусорной ДНК ». Однако более поздние исследования показывают, что, хотя ДНК, кодирующая белок, составляет едва 2% генома человека , около 80% оснований в геноме могут быть экспрессированы, поэтому термин «мусорная ДНК» может быть неправильным. [27]
Структура и функции [ править ]
Структура [ править ]


Структура гена, кодирующего белок, состоит из множества элементов, из которых реальная последовательность, кодирующая белок, часто составляет лишь небольшую часть. К ним относятся интроны и нетранслируемые области зрелой мРНК. Некодирующие гены также могут содержать интроны, которые удаляются во время процессинга для получения зрелой функциональной РНК.
Все гены связаны с регуляторными последовательностями , необходимыми для их экспрессии. Во-первых, генам необходима последовательность промотора . Промотор распознается и связывается факторами транскрипции , которые рекрутируют и помогают РНК-полимеразе связываться с областью для инициации транскрипции. [51] : 7.1 Распознавание обычно происходит в виде консенсусной последовательности, такой как блок ТАТА . Ген может иметь более одного промотора, в результате чего информационные РНК ( мРНК ) различаются по тому, насколько далеко они простираются на 5'-конце. [58] Гены с высокой степенью транскрипции имеют «сильные» промоторные последовательности, которые образуют прочные ассоциации с факторами транскрипции, тем самым инициируя транскрипцию с высокой скоростью. Другие гены имеют «слабые» промоторы, которые образуют слабые ассоциации с факторами транскрипции и реже инициируют транскрипцию. [51] : 7.2 Эукариотические промоторные области гораздо сложнее и их труднее идентифицировать, чем прокариотические промоторы. [51] : 7.3
Кроме того, гены могут иметь регуляторные области, расположенные на много тысяч оснований выше или ниже гена, которые изменяют экспрессию. Они действуют путем связывания с факторами транскрипции, которые затем заставляют ДНК образовывать петлю, так что регуляторная последовательность (и связанный фактор транскрипции) становится близко к сайту связывания РНК-полимеразы. [59] Например, энхансеры усиливают транскрипцию, связывая белок- активатор , который затем помогает привлечь РНК-полимеразу к промотору; и наоборот, сайленсеры связывают белки- репрессоры и делают ДНК менее доступной для РНК-полимеразы. [60]
Зрелая информационная РНК, полученная из генов, кодирующих белок, содержит нетранслируемые области на обоих концах, которые содержат сайты связывания для рибосом , РНК-связывающих белков , микроРНК , а также терминатор , а также стартовые и стоп-кодоны . [61] Кроме того, большинство открытых рамок считывания эукариот содержат нетранслируемые интроны , которые удаляются, и экзоны , которые соединяются вместе в процессе, известном как сплайсинг РНК . Наконец, концы транскриптов генов определяются сайтами расщепления и полиаденилирования (CPA) , где вновь образованная пре-мРНК расщепляется, и к 3'-концу добавляется цепочка из ~200 аденозинмонофосфатов. Поли (А) -хвост защищает зрелую мРНК от деградации и выполняет другие функции, влияя на трансляцию, локализацию и транспорт транскрипта из ядра. Сплайсинг с последующим CPA приводит к образованию окончательной зрелой мРНК , которая кодирует белок или продукт РНК. [62]
Многие некодирующие гены у эукариот имеют разные механизмы терминации транскрипции и не имеют поли(А)-хвостов.
Многие прокариотические гены организованы в опероны с множеством кодирующих белок последовательностей, которые транскрибируются как единое целое. [63] [64] Гены оперона транскрибируются в виде непрерывной информационной РНК , называемой полицистронной мРНК . Термин цистрон в этом контексте эквивалентен гену. Транскрипция мРНК оперона часто контролируется репрессором , который может находиться в активном или неактивном состоянии в зависимости от присутствия специфических метаболитов. [65] ДНК в начале оперона, называемой операторной областью , и подавляет транскрипцию оперона В активном состоянии репрессор связывается с последовательностью ; когда репрессор неактивен, может произойти транскрипция оперона (см., например, оперон Lac ). Продукты генов-оперонов обычно имеют родственные функции и участвуют в одной и той же регуляторной сети . [51] : 7.3
Сложность [ править ]
Хотя многие гены имеют простую структуру, как и большая часть биологии, другие могут быть весьма сложными или представлять собой необычные крайние случаи. Эукариотические гены часто имеют интроны, которые часто намного больше экзонов. [66] [67] и в эти интроны могут даже быть вложены другие гены . [68] Ассоциированные энхансеры могут находиться на расстоянии многих тысяч оснований или даже находиться на совершенно разных хромосомах, действующих посредством физического контакта между двумя хромосомами. [69] [70] Один ген может кодировать множество различных функциональных продуктов путем альтернативного сплайсинга , и, наоборот, ген может быть разделен по хромосомам, но эти транскрипты снова объединяются в функциональную последовательность путем транс-сплайсинга . [71] Также возможно, что перекрывающиеся гены имеют общую часть своей последовательности ДНК либо на противоположных цепях, либо на одной и той же цепи (в другой рамке считывания или даже в одной и той же рамке считывания). [72]
Экспрессия гена [ править ]
Во всех организмах требуется два шага, чтобы прочитать информацию, закодированную в ДНК гена, и произвести указанный ею белок. Сначала ДНК гена транскрибируется в информационную РНК ( мРНК ). [51] : 6.1 Во-вторых, эта мРНК транслируется в белок. [51] : 6.2 Гены, кодирующие РНК, еще должны пройти первый этап, но не транслируются в белок. [73] Процесс производства биологически функциональной молекулы РНК или белка называется экспрессией гена , а полученная молекула называется генным продуктом .
Генетический код [ править ]

Нуклеотидная последовательность ДНК гена определяет аминокислотную последовательность белка через генетический код . Наборы из трех нуклеотидов, известные как кодоны , соответствуют определенной аминокислоте. [51] : 6 Принцип, согласно которому три последовательных основания ДНК кодируют каждую аминокислоту, был продемонстрирован в 1961 году с использованием мутаций сдвига рамки считывания в гене rIIB бактериофага Т4. [74] (см. эксперимент Крика, Бреннера и др. ).
Кроме того, « старт-кодон » и три « стоп-кодона » обозначают начало и конец области, кодирующей белок . Существует 64 возможных кодона (четыре возможных нуклеотида в каждом из трех положений, следовательно, 4 возможных кодона). 3 возможные кодоны) и всего 20 стандартных аминокислот; следовательно, код является избыточным, и несколько кодонов могут обозначать одну и ту же аминокислоту. Соответствие между кодонами и аминокислотами практически универсально для всех известных живых организмов. [75]
Транскрипция [ править ]
Транскрипция производит одноцепочечную молекулу РНК , известную как информационная РНК , нуклеотидная последовательность которой комплементарна ДНК, с которой она была транскрибирована. [51] : 6.1 мРНК действует как промежуточное звено между геном ДНК и его конечным белковым продуктом. ДНК гена используется в качестве матрицы для создания комплементарной мРНК. мРНК соответствует последовательности кодирующей цепи ДНК гена , поскольку она синтезируется как дополнение к цепи матрицы . Транскрипция осуществляется ферментом, называемым РНК -полимеразой , который считывает цепочку матрицы в направлении от 3’ к 5’ и синтезирует РНК от 5’ к 3’ . Чтобы инициировать транскрипцию, полимераза сначала распознает и связывает промоторную область гена. Таким образом, основным механизмом регуляции генов является блокирование или секвестрация промоторной области либо путем прочного связывания молекулами- репрессорами , которые физически блокируют полимеразу, либо путем организации ДНК так, что промоторная область недоступна. [51] : 7
У прокариот транскрипция происходит в цитоплазме ; для очень длинных транскриптов трансляция может начинаться с 5'-конца РНК, в то время как 3'-конец все еще транскрибируется. У эукариот транскрипция происходит в ядре, где хранится ДНК клетки. Молекула РНК, продуцируемая полимеразой, известна как первичный транскрипт и подвергается посттранскрипционным модификациям перед экспортом в цитоплазму для трансляции. Одной из выполненных модификаций является сплайсинг интронов , которые представляют собой последовательности в транскрибируемой области, которые не кодируют белок. Альтернативные механизмы сплайсинга могут привести к тому, что зрелые транскрипты одного и того же гена будут иметь разные последовательности и, таким образом, кодировать разные белки. Это основная форма регуляции в эукариотических клетках, а также встречается у некоторых прокариот. [51] : 7.5 [76]
Перевод [ править ]

Трансляция — это процесс, при котором зрелая молекула мРНК используется в качестве матрицы для синтеза нового белка . [51] : 6.2 Трансляция осуществляется рибосомами , крупными комплексами РНК и белка, ответственными за проведение химических реакций по присоединению новых аминокислот к растущей полипептидной цепи путем образования пептидных связей . Генетический код считывается по три нуклеотида одновременно в единицах, называемых кодонами , посредством взаимодействия со специализированными молекулами РНК, называемыми транспортными РНК (тРНК). Каждая тРНК имеет три неспаренных основания, известных как антикодон , которые комплементарны кодону, который она считывает на мРНК. ТРНК также ковалентно присоединена к аминокислоте, указанной комплементарным кодоном. Когда тРНК связывается с комплементарным кодоном в цепи мРНК, рибосома присоединяет свой аминокислотный груз к новой полипептидной цепи, которая синтезируется от аминоконца к карбоксильному концу . Во время и после синтеза большинство новых белков должны свернуть свою активную трехмерную структуру, прежде чем они смогут выполнять свои клеточные функции. [51] : 3
Регламент [ править ]
Гены регулируются таким образом, что они экспрессируются только тогда, когда продукт необходим, поскольку экспрессия требует ограниченных ресурсов. [51] : 7 Клетка регулирует экспрессию своих генов в зависимости от ее внешней среды (например, доступных питательных веществ , температуры и других стрессов ), ее внутренней среды (например, цикла клеточного деления , метаболизма , статуса инфекции ) и ее конкретной роли в многоклеточном организме. Экспрессию генов можно регулировать на любом этапе: от инициации транскрипции до процессинга РНК и посттрансляционной модификации белка. Регуляция генов метаболизма лактозы в E. coli ( lac -оперон ) была первым подобным механизмом, описанным в 1961 году. [77]
Гены РНК [ править ]
Типичный ген, кодирующий белок, сначала копируется в РНК в качестве промежуточного продукта при производстве конечного белкового продукта. [51] : 6.1 В других случаях молекулы РНК являются реальными функциональными продуктами, как, например, при синтезе рибосомальной РНК и транспортной РНК . Некоторые РНК, известные как рибозимы, способны выполнять ферментативные функции , тогда как другие, такие как микроРНК и рибопереключатели, выполняют регуляторную роль. Последовательности ДНК , с которых транскрибируются такие РНК, известны как некодирующие гены РНК . [73]
Некоторые вирусы хранят весь свой геном в виде РНК и вообще не содержат ДНК. [78] [79] Поскольку они используют РНК для хранения генов, их клеточные хозяева могут синтезировать их белки сразу после заражения и без задержки в ожидании транскрипции. [80] С другой стороны, РНК- ретровирусы , такие как ВИЧ , требуют обратной транскрипции своего генома из РНК в ДНК, прежде чем их белки смогут быть синтезированы.
Наследование [ править ]

Организмы наследуют свои гены от родителей. Бесполые организмы просто наследуют полную копию генома своего родителя. Половые организмы имеют две копии каждой хромосомы, поскольку они наследуют по одному полному набору от каждого родителя. [51] : 1
Менделевское наследование [ править ]
Согласно менделевскому наследованию организма , вариации фенотипа (наблюдаемых физических и поведенческих характеристик) частично обусловлены вариациями его генотипа (определенного набора генов). Каждый ген определяет определенный признак с различной последовательностью гена ( аллелей ), приводящей к различным фенотипам. Большинство эукариотических организмов (таких как растения гороха, над которыми работал Мендель) имеют по два аллеля для каждого признака, по одному унаследованному от каждого родителя. [51] : 20
Аллели в локусе могут быть доминантными или рецессивными ; доминантные аллели дают начало соответствующим фенотипам при спаривании с любой другой аллелью того же признака, тогда как рецессивные аллели дают начало соответствующему фенотипу только при спаривании с другой копией того же аллеля. Зная генотипы организмов, можно определить, какие аллели доминантные, а какие рецессивные. Например, если аллель, определяющая высокие стебли у растений гороха, доминирует над аллелем, определяющим короткие стебли, то растения гороха, унаследовавшие один высокий аллель от одного родителя и один короткий аллель от другого родителя, также будут иметь высокие стебли. Работа Менделя продемонстрировала, что аллели независимо сортируются при образовании гамет или зародышевых клеток , обеспечивая вариации в следующем поколении. Хотя менделевское наследование остается хорошей моделью для многих признаков, детерминированных отдельными генами (включая ряд хорошо известных генетических нарушений ), оно не включает физические процессы репликации ДНК и деления клеток. [81] [82]
Репликация ДНК и деление клеток [ править ]
Рост, развитие и размножение организмов зависят от деления клеток ; процесс, при котором одна клетка делится на две обычно идентичные дочерние клетки . Для этого необходимо сначала сделать дубликат каждого гена в геноме в процессе, называемом репликацией ДНК . [51] : 5.2 Копии создаются специализированными ферментами , известными как ДНК-полимеразы , которые «читают» одну цепь двойной спирали ДНК, известную как матричная цепь, и синтезируют новую комплементарную цепь. Поскольку двойная спираль ДНК удерживается вместе за счет спаривания оснований , последовательность одной цепи полностью определяет последовательность ее комплемента; следовательно, для создания точной копии ферменту необходимо прочитать только одну цепь. Процесс репликации ДНК полуконсервативный ; то есть копия генома, унаследованная каждой дочерней клеткой, содержит одну исходную и одну вновь синтезированную цепь ДНК. [51] : 5.2
Скорость репликации ДНК в живых клетках была впервые измерена как скорость удлинения ДНК фага Т4 в инфицированной фагом E. coli и оказалась впечатляюще быстрой. [83] В период экспоненциального роста ДНК при 37 °С скорость элонгации составила 749 нуклеотидов в секунду.
После завершения репликации ДНК клетка должна физически разделить две копии генома и разделиться на две отдельные мембраносвязанные клетки. [51] : 18.2 У прокариот ( бактерий и архей ) это обычно происходит посредством относительно простого процесса, называемого бинарным делением , при котором каждый кольцевой геном прикрепляется к клеточной мембране и разделяется на дочерние клетки, когда мембрана инвагинирует , разделяя цитоплазму на две мембраносвязанные части. . Бинарное деление происходит чрезвычайно быстро по сравнению со скоростью деления клеток у эукариот . Деление эукариотических клеток — более сложный процесс, известный как клеточный цикл ; Репликация ДНК происходит во время фазы этого цикла, известной как S-фаза , тогда как процесс сегрегации хромосом и расщепления цитоплазмы происходит во время M-фазы . [51] : 18.1
Молекулярная наследственность [ править ]
Дупликация и передача генетического материала от одного поколения клеток к другому является основой молекулярной наследственности и связи между классической и молекулярной картиной генов. Организмы наследуют характеристики своих родителей, поскольку клетки потомства содержат копии генов родительских клеток. У организмов, размножающихся бесполым путем , потомство будет генетической копией или клоном родительского организма. У организмов, размножающихся половым путем , специализированная форма деления клеток, называемая мейозом, приводит к образованию клеток, называемых гаметами или зародышевыми клетками , которые являются гаплоидными или содержат только одну копию каждого гена. [51] : 20.2 Гаметы, вырабатываемые самками, называются яйцеклетками или яйцеклетками, а гаметы, вырабатываемые самцами, называются сперматозоидами . Две гаметы сливаются, образуя диплоидную оплодотворенную яйцеклетку — одну клетку, содержащую два набора генов, по одной копии каждого гена от матери и одной от отца. [51] : 20
В процессе мейотического деления клеток иногда может произойти событие, называемое генетической рекомбинацией или кроссинговером , при котором длина ДНК на одной хроматиде заменяется длиной ДНК на соответствующей гомологичной несестринской хроматиде. Это может привести к реассортации связанных иначе аллелей. [51] : 5.5 Менделевский принцип независимого ассортимента утверждает, что каждый из двух родительских генов каждого признака независимо сортируется в гаметы; какой аллель организм наследует по одному признаку, не связан с тем, какой аллель он наследует по другому признаку. Фактически это справедливо только для генов, которые не находятся в одной хромосоме или расположены очень далеко друг от друга на одной хромосоме. Чем ближе два гена лежат в одной хромосоме, тем теснее они будут связаны в гаметах и тем чаще они будут появляться вместе (так называемое генетическое сцепление ). [84] Гены, расположенные очень близко, практически никогда не разделяются, поскольку крайне маловероятно, что между ними произойдет точка кроссовера. [84]
эволюция Молекулярная
Мутация [ править ]
Репликация ДНК по большей части чрезвычайно точна, однако ошибки ( мутации ). случаются [51] : 7.6 Частота ошибок в эукариотических клетках может составлять всего 10 −8 на нуклеотид на репликацию, [85] [86] тогда как для некоторых РНК-вирусов оно может достигать 10 −3 . [87] Это значит, что каждое поколение, каждый геном человека накапливает около 30 новых мутаций. [88] Небольшие мутации могут быть вызваны репликацией ДНК и последствиями повреждения ДНК и включают точечные мутации, при которых изменяется одно основание, и мутации сдвига рамки считывания, при которых одно основание вставляется или удаляется. Любая из этих мутаций может изменить ген по миссенсу (изменение кодона , кодирующего другую аминокислоту) или по нонсенсу (преждевременный стоп-кодон ). [89] Более крупные мутации могут быть вызваны ошибками рекомбинации и вызывать хромосомные аномалии, включая дупликацию , делецию, перестановку или инверсию больших участков хромосомы. Кроме того, механизмы репарации ДНК могут вносить мутационные ошибки при восстановлении физических повреждений молекулы. Восстановление, даже при мутации, более важно для выживания, чем восстановление точной копии, например, при восстановлении двухцепочечных разрывов . [51] : 5.4
Когда в популяции вида присутствует несколько различных аллелей гена, это называется полиморфным . Большинство различных аллелей функционально эквивалентны, однако некоторые аллели могут вызывать разные фенотипические признаки . Самая распространенная аллель гена называется диким типом , а редкие аллели называются мутантами . Генетическая изменчивость относительных частот различных аллелей в популяции обусловлена как естественным отбором, так и генетическим дрейфом . [90] Аллель дикого типа не обязательно является предком менее распространенных аллелей и не обязательно является более подходящей .
Большинство мутаций внутри генов нейтральны и не влияют на фенотип организма ( молчащие мутации ). Некоторые мутации не меняют аминокислотную последовательность, поскольку одну и ту же аминокислоту кодируют несколько кодонов ( синонимные мутации ). Другие мутации могут быть нейтральными, если они приводят к изменениям аминокислотной последовательности, но белок по-прежнему функционирует аналогично новой аминокислоте (например, консервативные мутации ). Однако многие мутации вредны или даже смертельны и удаляются из популяции путем естественного отбора. Генетические нарушения являются результатом вредных мутаций и могут быть вызваны спонтанной мутацией у пораженного человека или могут передаваться по наследству. Наконец, небольшая часть мутаций приносит пользу организма , улучшая приспособленность и чрезвычайно важна для эволюции, поскольку их направленный отбор приводит к адаптивной эволюции . [51] : 7.6
последовательностей Гомология
Взаимосвязь между генами можно измерить, сравнивая последовательности их ДНК. Если уровень сходства превышает минимальное значение, можно сделать вывод, что гены происходят от общего предка; они гомологичны . [91] [92] Гены, связанные прямым происхождением от общего предка, являются ортологичными: они обычно встречаются в одном и том же локусе у разных видов. Гены, родственные в результате дупликации генов, являются парологичными генами. [93] [94]
Часто полагают, что функции ортологичных генов более сходны, чем функции паралогичных, хотя различия минимальны. [95] [96]
Происхождение новых генов [ править ]
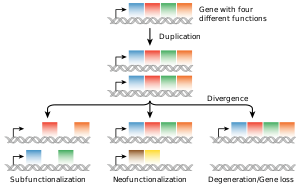
Наиболее распространенным источником новых генов в эукариотических линиях является дупликация генов , которая приводит к изменению числа копий существующего гена в геноме. [97] [98] Полученные гены (паралоги) могут затем расходиться по последовательности и функциям. Сформированные таким образом наборы генов составляют генное семейство . Дупликация и потеря генов внутри семьи являются обычным явлением и представляют собой основной источник эволюционного биоразнообразия . [99] Иногда дупликация гена может привести к образованию нефункциональной копии гена, или функциональная копия может подвергаться мутациям, которые приводят к потере функции; такие нефункциональные гены называются псевдогенами . [51] : 7.6
Гены-сироты , последовательность которых не имеет сходства с существующими генами, встречаются реже, чем дубликаты генов. Геном человека содержит примерно 18 [100] до 60 [101] гены, не имеющие идентифицируемых гомологов вне человека. Гены-сироты возникают в основном либо в результате de novo возникновения ранее некодирующей последовательности , либо в результате дупликации гена с последующим таким быстрым изменением последовательности, что исходное родство становится необнаружимым. [102] Гены de novo обычно короче и проще по структуре, чем большинство эукариотических генов, с небольшим количеством интронов или вообще без них. [97] В течение длительных периодов эволюции рождение генов de novo может быть ответственным за значительную часть таксономически ограниченных семейств генов. [103]
Горизонтальный перенос генов относится к передаче генетического материала посредством механизма, отличного от воспроизводства . Этот механизм является распространенным источником новых генов у прокариот , и иногда считается, что он больше способствует генетической изменчивости, чем дупликации генов. [104] Это распространенное средство распространения устойчивости к антибиотикам , вирулентности и адаптивных метаболических функций. [55] [105] Хотя горизонтальный перенос генов у эукариот встречается редко, вероятные примеры были идентифицированы в геномах протистов и водорослей , содержащих гены бактериального происхождения. [106] [107]
Геном [ править ]
Геном представляет собой совокупный генетический материал организма и включает в себя как гены, так и некодирующие последовательности . [108] Гены эукариот можно аннотировать с помощью FINDER. [109]
Количество генов [ править ]

Размер генома и количество кодируемых им генов сильно различаются у разных организмов. Наименьшие геномы встречаются у вирусов . [118] и вироиды (которые действуют как одиночный некодирующий ген РНК). [119] И наоборот, растения могут иметь чрезвычайно большие геномы. [120] рис . содержит >46 000 генов, кодирующих белок [114] Земли Общее количество генов, кодирующих белок ( протеом ), оценивается в 5 миллионов последовательностей. [121]
Хотя количество пар оснований ДНК в геноме человека известно с 1950-х годов, предполагаемое количество генов со временем менялось по мере того, как определения генов и методы их обнаружения уточнялись. Первоначальные теоретические предсказания количества генов человека в 1960-х и 1970-х годах были основаны на оценках мутационной нагрузки и количестве мРНК, и эти оценки, как правило, составляли около 30 000 генов, кодирующих белок. [122] [123] [124] В 1990-е годы предполагалось, что число генов достигает 100 000, а ранние данные по обнаружению мРНК ( метки выраженных последовательностей ) предполагали, что число генов превышает традиционное значение в 30 000 генов, о котором сообщалось в учебниках 1980-х годов. [125]
Первоначальный проект последовательностей генома человека подтвердил более ранние предсказания о наличии около 30 000 генов, кодирующих белки, однако эта оценка упала примерно до 19 000 в связи с продолжающимся проектом аннотаций GENCODE . [126] Число некодирующих генов точно неизвестно, но по последним оценкам Ensembl, их число составляет 26 000. [127]
Основные гены [ править ]
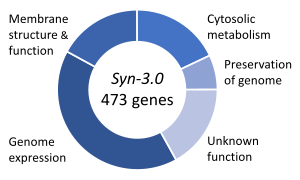
Эссенциальные гены — это набор генов, которые считаются критически важными для выживания организма. [129] Это определение предполагает наличие в изобилии всех необходимых питательных веществ и отсутствие экологического стресса. Лишь небольшая часть генов организма имеет важное значение. У бактерий примерно 250–400 генов необходимы для Escherichia coli и Bacillus subtilis , что составляет менее 10% их генов. [130] [131] [132] Половина этих генов являются ортологами у обоих организмов и в значительной степени участвуют в синтезе белка . [132] У почкующихся дрожжей Saccharomyces cerevisiae количество незаменимых генов несколько выше — 1000 генов (~20% их генов). [133] Хотя это число сложнее измерить у высших эукариот, мыши и люди, по оценкам, имеют около 2000 основных генов (~ 10% их генов). [134] Синтетический организм Syn 3 имеет минимальный геном, состоящий из 473 основных генов и квазинезаменимых генов (необходимых для быстрого роста), хотя 149 имеют неизвестную функцию. [128]
К важнейшим генам относятся гены домашнего хозяйства (критически важные для основных функций клеток). [135] а также гены, которые экспрессируются в разные периоды развития или жизненного цикла организма . [136] Гены «домашнего хозяйства» используются в качестве экспериментального контроля при анализе экспрессии генов , поскольку они конститутивно экспрессируются на относительно постоянном уровне.
Генетическая и геномная номенклатура [ править ]
Номенклатура генов была установлена Комитетом по номенклатуре генов HUGO (HGNC), комитетом Организации генома человека , для каждого известного гена человека в форме утвержденного названия гена и символа (краткой аббревиатуры ), доступ к которым можно получить через база данных, поддерживаемая HGNC. Символы выбираются уникальными, и каждый ген имеет только один символ (хотя утвержденные символы иногда меняются). Символы предпочтительно должны соответствовать другим членам семейства генов и гомологам других видов, особенно мыши , поскольку она играет роль общего модельного организма . [137]
Генная инженерия [ править ]
организма Генная инженерия — это модификация генома с помощью биотехнологий . С 1970-х годов было разработано множество методов, позволяющих специально добавлять, удалять и редактировать гены в организме. [138] Недавно разработанные методы геномной инженерии используют сконструированные нуклеазы ферменты для целевого восстановления ДНК в хромосоме, чтобы либо разрушить, либо отредактировать ген после устранения разрыва. [139] [140] [141] [142] Сопутствующий термин «синтетическая биология» иногда используется для обозначения обширной генной инженерии организма. [143]
Генная инженерия в настоящее время является рутинным инструментом исследования модельных организмов . Например, гены легко добавляются к бактериям. [144] и линии нокаутных мышей с нарушенной функцией определенного гена используются для исследования функции этого гена. [145] [146] Многие организмы были генетически модифицированы для применения в сельском хозяйстве , промышленной биотехнологии и медицине .
Для многоклеточных организмов обычно создают эмбрион , который вырастает во взрослый генетически модифицированный организм . [147] Однако геномы клеток взрослого организма можно редактировать с помощью методов генной терапии для лечения генетических заболеваний.
См. также [ править ]
- Биологический патент
- Эпигенетика
- Геноцентричный взгляд на эволюцию
- Дозировка гена
- Генофонд
- Избыточность генов
- Замалчивание генов
- Генетический алгоритм
- Гаплотип
- Список программного обеспечения для прогнозирования генов
- Списки человеческих генов
- Предиктивная медицина
- Локус количественного признака
- Эгоистичный генетический элемент
- Полногеномное секвенирование
Ссылки [ править ]
Цитаты [ править ]
- ↑ Перейти обратно: Перейти обратно: а б с Оргогозо В., Пелуффо А.Е., Моризо Б. (2016). «Менделевский ген» и «молекулярный ген»: две соответствующие концепции генетических единиц» (PDF) . Актуальные темы биологии развития . 119 : 1–26. дои : 10.1016/bs.ctdb.2016.03.002 . ПМИД 27282022 . S2CID 24583286 .
- ^ «Что такое ген?: MedlinePlus Genetics» . МедлайнПлюс . 17 сентября 2020 г. Проверено 4 января 2021 г.
- ^ Хирш ЭД (2002). Новый словарь культурной грамотности . Бостон: Хоутон Миффлин. ISBN 0-618-22647-8 . OCLC 50166721 .
- ^ Элстон Р.К., Сатагопан Дж.М., Сан С. (2012). «Генетическая терминология». Статистическая генетика человека . Методы молекулярной биологии. Том. 850. Хумана Пресс. стр. 1–9. дои : 10.1007/978-1-61779-555-8_1 . ISBN 978-1-61779-554-1 . ПМЦ 4450815 . ПМИД 22307690 .
- ↑ Перейти обратно: Перейти обратно: а б Йохансен В. (1909). Elemente der exakten Erblichkeitslehre [ Элементы точной теории наследственности ] (на немецком языке). Йена, Германия: Густав Фишер. п. 124. Со с. 124: «Dieses "etwas" in den Gameten bezw. in der Zygote, ... – kurz, было wir eben Gene nennen wollen – bedingt sind». (Это «нечто» в гаметах или зиготе, имеющее решающее значение для характера организма, обычно называют весьма двусмысленным термином Anlagen [primordium, от немецкого слова Anlage, означающего «план, расположение; черновой набросок». ] Было предложено много других терминов, в основном, к сожалению, в более тесной связи с некоторыми гипотетическими мнениями. Слово «панген», введенное Дарвином, возможно, чаще всего используется вместо слова « панген ». выбрано удачно, так как это сложное слово, содержащее корни pan (средняя форма от Πας весь, каждый) и gen (от γί-γ(ε)ν-ομαι, становиться только значением этого последнего [т.е., gen ] здесь принимается во внимание только основная идея — [а именно] о том, что признак в развивающемся организме может определяться или подвергаться влиянию «чего-то» в гаметах — не должна найти выражения никакая гипотеза о природе этого «чего-то». «должно быть постулировано или поддержано им. По этой причине кажется, что проще всего использовать изолированно последний слог gen от известного дарвиновского слова, которое единственное и представляет для нас интерес, чтобы заменить им бедное, двусмысленное слово Anlage . Поэтому мы будем говорить просто «ген» и «гены» вместо «панген» и «пангены». Слово «ген» совершенно свободно от какой-либо гипотезы; оно выражает лишь установленный факт, что в любом случае многие признаки организма определяются специфическими, отделимыми и, следовательно, независимыми «условиями», «основаниями», «планами» — словом, именно тем, что мы хотим назвать генами.)
- ↑ Перейти обратно: Перейти обратно: а б с д Кампуракис К (2017). Осмысление генов . Кембридж, Великобритания: Издательство Кембриджского университета.
- ^ Герике Н., Хагберг М. (5 декабря 2006 г.). «Определение исторических моделей функции генов и их связь с пониманием студентами генетики». Наука и образование . 16 (7–8): 849–881. Бибкод : 2007Sc&Ed..16..849G . дои : 10.1007/s11191-006-9064-4 . S2CID 144613322 .
- ^ Менье Р. (2022). «Стэнфордская энциклопедия философии: Джин» . Стэнфордская энциклопедия философии . Проверено 28 февраля 2023 г.
- ^ Келлис М., Уолд Б., Снайдер М.П., Бернштейн Б.Е., Кундадже А., Маринов Г.К. и др. (апрель 2014 г.). «Определение функциональных элементов ДНК в геноме человека» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (17): 6131–8. Бибкод : 2014PNAS..111.6131K . дои : 10.1073/pnas.1318948111 . ПМК 4035993 . ПМИД 24753594 .
- ↑ Перейти обратно: Перейти обратно: а б Докинз Р. (1976). Эгоистичный ген . Оксфорд, Великобритания: Издательство Оксфордского университета.
- ^ Штольц К., Гриффитс П. (2004). «Гены: философский анализ, подвергнутый испытанию» . История и философия наук о жизни . 26 (1): 5–28. дои : 10.1080/03919710412331341621 . JSTOR 23333378 . ПМИД 15791804 .
- ^ Бидл Г.В., Татум Э.Л. (ноябрь 1941 г.). «Генетический контроль биохимических реакций нейроспоры» . Труды Национальной академии наук Соединенных Штатов Америки . 27 (11): 499–506. Бибкод : 1941ПНАС...27..499Б . дои : 10.1073/pnas.27.11.499 . ПМЦ 1078370 . ПМИД 16588492 .
- ^ Горовиц Н.Х., Берг П., Сингер М., Ледерберг Дж., Сусман М., Добли Дж., Кроу Дж.Ф. (январь 2004 г.). «Столетие: Джордж Бидл, 1903–1989» . Генетика . 166 (1): 1–10. дои : 10.1534/генетика.166.1.1 . ПМК 1470705 . ПМИД 15020400 .
- ^ Джадсон Х.Ф. (1996). Восьмой день творения (Расширенная ред.). Плейнвью, Нью-Йорк (США): Лабораторное издательство Колд-Спринг-Харбор.
- ↑ Перейти обратно: Перейти обратно: а б Уотсон Джей Ди (1965). Молекулярная биология гена . Нью-Йорк, штат Нью-Йорк, США: WA Benjamin, Inc.
- ^ Альбертс Б., Брэй Д., Льюис Дж., Рафф М., Робертс К., Уотсон Дж.Д. (1994). Молекулярная биология клетки: третье издание . Лондон, Великобритания: ISBN Garland Publishing, Inc. 0-8153-1619-4 .
- ^ Моран Л.А., Хортон Х.Р., Скримджер К.Г., Перри, доктор медицинских наук (2012). Принципы биохимии: пятое издание . Река Аппер-Седл, Нью-Джерси, США: Пирсон.
- ^ Левин Б. (2004). Гены VIII . Река Аппер-Сэддл, Нью-Джерси, США: Пирсон/Прентис-Холл.
- ^ Пиовесан А., Пеллери М.К., Антонарос Ф., Стрипполи П., Каракаузи М. и Витале Л. (2019). «О длине, массе и GC-содержании генома человека» . Исследовательские заметки BMC . 12 (1): 106–173. дои : 10.1186/s13104-019-4137-z . ПМК 6391780 . ПМИД 30813969 .
- ^ Хубе Ф. и Франкастель С. (2015). «Интроны млекопитающих: когда мусор порождает молекулярное разнообразие» . Международный журнал молекулярных наук . 16 (3): 4429–4452. дои : 10.3390/ijms16034429 . ПМЦ 4394429 . ПМИД 25710723 .
- ^ Фрэнсис В.Р. и Верхайде Г. (2017). «Аналогичные соотношения интронов и межгенной последовательности в геномах животных» . Геномная биология и эволюция . 9 (6): 1582–1598. дои : 10.1093/gbe/evx103 . ПМЦ 5534336 . ПМИД 28633296 .
- ^ Мортола Э., Лонг М. (2021). «Превращаем мусор в нас: как рождаются гены» . Американский учёный . 109 : 174–182.
- ^ Хопкин К. (2009). «Развивающееся определение гена: с открытием того, что почти весь геном транскрибируется, определение «гена» нуждается в еще одном пересмотре». Бионаука . 59 : 928–931. дои : 10.1525/био.2009.59.11.3 . S2CID 88157272 .
- ^ Пирсон Х (2006). «Что такое ген?» . Природа . 441 (7092): 399–401. Бибкод : 2006Natur.441..398P . дои : 10.1038/441398a . ПМИД 16724031 . S2CID 4420674 .
- ^ Пенниси Э (2007). «Исследование ДНК заставляет переосмыслить, что значит быть геном» . Наука . 316 (5831): 1556–1557. дои : 10.1126/science.316.5831.1556 . ПМИД 17569836 . S2CID 36463252 .
- ^ Вольф Й.И., Казлаускас Д., Иранзо Дж., Люсия-Санс А., Кун Дж.Х., Крупович М. и др. (ноябрь 2018 г.). Раканьелло В.Р. (ред.). «Происхождение и эволюция глобального РНК-вирома» . мБио 9 (6). Эрик Делварт, Луи Энхуанес: e02329–18. дои : 10.1128/mBio.02329-18 . ПМК 6282212 . ПМИД 30482837 .
- ↑ Перейти обратно: Перейти обратно: а б Пенниси Э (июнь 2007 г.). «Геномика. Изучение ДНК заставляет переосмыслить, что значит быть геном» . Наука . 316 (5831): 1556–7. дои : 10.1126/science.316.5831.1556 . ПМИД 17569836 . S2CID 36463252 .
- ^ Маранде В., Бургер Дж. (октябрь 2007 г.). «Митохондриальная ДНК как геномная головоломка». Наука . 318 (5849). AAAS: 415. Бибкод : 2007Sci...318..415M . дои : 10.1126/science.1148033 . ПМИД 17947575 . S2CID 30948765 .
- ^ Парра Г., Реймонд А., Даббусе Н., Дермицакис Э.Т., Кастело Р., Томсон Т.М. и др. (январь 2006 г.). «Тандемный химеризм как средство увеличения сложности белков в геноме человека» . Геномные исследования . 16 (1): 37–44. дои : 10.1101/гр.4145906 . ПМЦ 1356127 . ПМИД 16344564 .
- ↑ Перейти обратно: Перейти обратно: а б с Герштейн М.Б., Брюс С., Розовский Дж.С., Чжэн Д., Ду Дж., Корбель Дж.О. и др. (июнь 2007 г.). «Что такое ген пост-КОДИРОВАНИЕ? История и обновленное определение» . Геномные исследования . 17 (6): 669–81. дои : 10.1101/гр.6339607 . ПМИД 17567988 .
- ^ Нобл Д. (сентябрь 2008 г.). «Гены и причинно-следственная связь» . Философские труды. Серия А. Математические, физические и технические науки . 366 (1878): 3001–15. Бибкод : 2008RSPTA.366.3001N . дои : 10.1098/rsta.2008.0086 . ПМИД 18559318 .
- ^ «Смешивание наследования — обзор | Темы ScienceDirect» .
- ^ «генезис» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Магнер Л.Н. (2002). История наук о жизни (Третье изд.). Марсель Деккер , CRC Press . п. 371. ИСБН 978-0-203-91100-6 .
- ^ Хениг Р.М. (2000). Монах в саду: потерянный и найденный гений Грегора Менделя, отца генетики . Бостон: Хоутон Миффлин. стр. 1–9 . ISBN 978-0395-97765-1 .
- ↑ Перейти обратно: Перейти обратно: а б де Врис Х (1889). Intracellle Pangenese [ Внутриклеточный пангенезис ] (на немецком языке). Перевод Gager CS . Йена: Верлаг фон Густава Фишера. Переведено в 1908 году с немецкого на английский издательством Open Court Publishing Co., Чикаго, 1910 год.
- ^ Бейтсон В. (1906). «Прогресс генетических исследований» . В Уилксе В. (ред.). Отчет Третьей Международной конференции по генетике 1906 года . Лондон, Англия: Королевское садоводческое общество. стр. 90–97.
... сама наука [т.е. изучение селекции и гибридизации растений] до сих пор безымянна, и мы можем описать наши занятия только громоздкими и часто вводящими в заблуждение перифразами. Чтобы преодолеть эту трудность, я предлагаю на рассмотрение Конгресса термин «Генетика» , который достаточно указывает на то, что наши труды посвящены выяснению явлений наследственности и изменчивости: другими словами, физиологии происхождения с подразумеваемым отношением к теоретические проблемы эволюционистов и систематиков и их применение к практическим проблемам селекционеров, будь то животных или растений.
- ^ Эйвери ОТ, Маклауд СМ, Маккарти М (февраль 1944 г.). «Исследование химической природы вещества, индуцирующего трансформацию пневмококков типа: индукция трансформации фракцией дезоксирибонуклеиновой кислоты, выделенной из пневмококка III типа» . Журнал экспериментальной медицины . 79 (2): 137–58. дои : 10.1084/jem.79.2.137 . ПМК 2135445 . ПМИД 19871359 . Перепечатка: Эйвери ОТ, Маклауд СМ, Маккарти М (февраль 1979 г.). «Исследование химической природы вещества, индуцирующего трансформацию пневмококков типа. Индукция трансформации фракцией дезоксирибонуклеиновой кислоты, выделенной из пневмококка III типа» . Журнал экспериментальной медицины . 149 (2): 297–326. дои : 10.1084/jem.149.2.297 . ПМК 2184805 . ПМИД 33226 .
- ^ Херши А.Д., Чейз М. (май 1952 г.). «Независимые функции вирусного белка и нуклеиновой кислоты в росте бактериофага» . Журнал общей физиологии . 36 (1): 39–56. дои : 10.1085/jgp.36.1.39 . ПМК 2147348 . ПМИД 12981234 .
- ^ Джадсон Х (1979). Восьмой день творения: творцы революции в биологии . Лабораторный пресс Колд-Спринг-Харбор. стр. 51–169. ISBN 978-0-87969-477-7 .
- ^ Уотсон Дж. Д., Крик Ф. Х. (апрель 1953 г.). «Молекулярная структура нуклеиновых кислот: структура нуклеиновой кислоты дезоксирибозы» (PDF) . Природа . 171 (4356): 737–8. Бибкод : 1953Natur.171..737W . дои : 10.1038/171737a0 . ПМИД 13054692 . S2CID 4253007 .
- ^ Бензер С. (июнь 1955 г.). «Тонкая структура генетической области бактериофага» . Труды Национальной академии наук Соединенных Штатов Америки . 41 (6): 344–54. Бибкод : 1955ПНАС...41..344Б . дои : 10.1073/pnas.41.6.344 . ПМК 528093 . ПМИД 16589677 .
- ^ Бензер С. (ноябрь 1959 г.). «К топологии тонкой генетической структуры» . Труды Национальной академии наук Соединенных Штатов Америки . 45 (11): 1607–20. Бибкод : 1959PNAS...45.1607B . дои : 10.1073/pnas.45.11.1607 . ПМК 222769 . ПМИД 16590553 .
- ^ Мин Джоу В., Хагеман Г., Изеберт М., Фирс В. (май 1972 г.). «Нуклеотидная последовательность гена, кодирующего белок оболочки бактериофага MS2». Природа . 237 (5350): 82–8. Бибкод : 1972Natur.237...82J . дои : 10.1038/237082a0 . ПМИД 4555447 . S2CID 4153893 .
- ^ Сэнгер Ф., Никлен С., Коулсон А.Р. (декабрь 1977 г.). «Секвенирование ДНК с ингибиторами обрыва цепи» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (12): 5463–7. Бибкод : 1977PNAS...74.5463S . дои : 10.1073/pnas.74.12.5463 . ПМК 431765 . ПМИД 271968 .
- ^ Адамс Ю. (2008). «Технологии секвенирования ДНК» . Знания о природном образовании . Научная таблица. 1 (1). Издательская группа «Природа»: 193.
- ^ Хаксли Дж. (1942). Эволюция: современный синтез . Кембридж, Массачусетс: MIT Press. ISBN 978-0262513661 .
- ^ Уильямс Г.К. (2001). Адаптация и естественный отбор: критика некоторых современных эволюционных идей (онлайн-изд.). Принстон: Издательство Принстонского университета. ISBN 9781400820108 .
- ^ Докинз Р. (1989). Расширенный фенотип (изд. В мягкой обложке). Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-286088-0 .
- ^ Дюре Л. (2008). «Нейтральная теория: нулевая гипотеза молекулярной эволюции» . Природное образование . 1 : 218.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть Альбертс Б. , Джонсон А., Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). Молекулярная биология клетки (Четвертое изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 .
- ^ Страйер Л., Берг Дж.М., Тимочко Дж.Л. (2002). Биохимия (5-е изд.). Сан-Франциско: WH Freeman. ISBN 978-0-7167-4955-4 .
- ^ Больцер А., Крет Г., Соловей И., Келер Д., Сарачоглу К., Фаут С. и др. (май 2005 г.). «Трехмерные карты всех хромосом в ядрах мужских фибробластов человека и розетках прометафазы» . ПЛОС Биология . 3 (5): е157. дои : 10.1371/journal.pbio.0030157 . ПМЦ 1084335 . ПМИД 15839726 .

- ^ Брэйг М., Шмитт, Калифорния (март 2006 г.). «Онкоген-индуцированное старение: тормозить развитие опухоли» . Исследования рака . 66 (6): 2881–4. дои : 10.1158/0008-5472.CAN-05-4006 . ПМИД 16540631 .
- ↑ Перейти обратно: Перейти обратно: а б Беннетт П.М. (март 2008 г.). «Плазмида, кодирующая устойчивость к антибиотикам: приобретение и перенос генов устойчивости к антибиотикам в бактериях» . Британский журнал фармакологии . 153 (Приложение 1): С347-57. дои : 10.1038/sj.bjp.0707607 . ПМК 2268074 . ПМИД 18193080 .
- ^ Международный консорциум по секвенированию генома человека (октябрь 2004 г.). «Завершение эухроматической последовательности генома человека» . Природа . 431 (7011): 931–45. Бибкод : 2004Natur.431..931H . дои : 10.1038/nature03001 . ПМИД 15496913 .
- ↑ Перейти обратно: Перейти обратно: а б Шафи, Томас; Лоу, Рохан (2017). «Строение генов эукариот и прокариот» . Викижурнал медицины . 4 (1). дои : 10.15347/wjm/2017.002 . ISSN 2002-4436 .
- ^ Мортазави А., Уильямс Б.А., МакКью К., Шеффер Л., Уолд Б. (июль 2008 г.). «Картирование и количественная оценка транскриптомов млекопитающих с помощью RNA-Seq». Природные методы . 5 (7): 621–8. дои : 10.1038/nmeth.1226 . ПМИД 18516045 . S2CID 205418589 .
- ^ Пеннаккио Л.А., Бикмор В., Дин А., Нобрега М.А., Беджерано Дж. (апрель 2013 г.). «Усилители: пять основных вопросов» . Обзоры природы. Генетика . 14 (4): 288–95. дои : 10.1038/nrg3458 . ПМЦ 4445073 . ПМИД 23503198 .
- ^ Мастон Г.А., Эванс С.К., Грин М.Р. (2006). «Транскрипционные регуляторные элементы в геноме человека» . Ежегодный обзор геномики и генетики человека . 7 : 29–59. дои : 10.1146/annurev.genom.7.080505.115623 . ПМИД 16719718 .
- ^ Миньоне Ф, Гисси С, Люни С, Пезоле Дж (28 февраля 2002 г.). «Нетранслируемые области мРНК» . Геномная биология . 3 (3): ОБЗОРЫ0004. doi : 10.1186/gb-2002-3-3-reviews0004 . ПМК 139023 . ПМИД 11897027 .
- ^ Бикнелл А.А., Сеник С., Чуа Х.Н., Рот Ф.П., Мур М.Дж. (декабрь 2012 г.). «Интроны в НТО: почему мы должны перестать их игнорировать» . Биоэссе . 34 (12): 1025–34. doi : 10.1002/bies.201200073 . ПМИД 23108796 . S2CID 5808466 .
- ^ Сальгадо Х., Морено-Хагельсиб Г., Смит Т.Ф., Колладо-Видес Дж. (июнь 2000 г.). «Опероны Escherichia coli: геномный анализ и прогнозы» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (12): 6652–7. Бибкод : 2000PNAS...97.6652S . дои : 10.1073/pnas.110147297 . ЧВК 18690 . ПМИД 10823905 .
- ^ Блюменталь Т. (ноябрь 2004 г.). «Опероны у эукариот» . Брифинги по функциональной геномике и протеомике . 3 (3): 199–211. дои : 10.1093/bfgp/3.3.199 . ПМИД 15642184 .
- ^ Джейкоб Ф., Моно Дж. (июнь 1961 г.). «Генетические регуляторные механизмы синтеза белков». Журнал молекулярной биологии . 3 (3): 318–56. дои : 10.1016/S0022-2836(61)80072-7 . ПМИД 13718526 . S2CID 19804795 .
- ^ Поццоли У, Меноцци Г, Коми ГП, Кальяни Р, Брезолин Н, Сирони М (январь 2007 г.). «Размер интрона у млекопитающих: сложность сочетается с экономией». Тенденции в генетике . 23 (1): 20–24. дои : 10.1016/j.tig.2006.10.003 . ПМИД 17070957 .
- ^ Марэ Дж., Нувелле П., Кейтли П.Д., Чарльзуорт Б. (май 2005 г.). «Размер интрона и эволюция экзонов у дрозофилы» . Генетика . 170 (1): 481–485. дои : 10.1534/genetics.104.037333 . ПМЦ 1449718 . ПМИД 15781704 .
- ^ Кумар А. (сентябрь 2009 г.). «Обзор вложенных генов в геномах эукариот» . Эукариотическая клетка . 8 (9): 1321–1329. дои : 10.1128/EC.00143-09 . ПМЦ 2747821 . ПМИД 19542305 . .
- ^ Спилианакис К.Г., Лалиоти, доктор медицинских наук, Таун Т., Ли Г.Р., Флавелл Р.А. (июнь 2005 г.). «Межхромосомные ассоциации между альтернативно экспрессируемыми локусами». Природа . 435 (7042): 637–645. Бибкод : 2005Natur.435..637S . дои : 10.1038/nature03574 . ПМИД 15880101 . S2CID 1755326 .
- ^ Уильямс А., Спилианакис К.Г., Флавелл Р.А. (апрель 2010 г.). «Межхромосомная ассоциация и регуляция генов у транс» . Тенденции в генетике . 26 (4): 188–197. дои : 10.1016/j.tig.2010.01.007 . ПМЦ 2865229 . ПМИД 20236724 .
- ^ Лэй Ц, Ли С, Цзо З, Хуан С, Ченг Х, Чжоу Р (март 2016 г.). «Эволюционный взгляд на транс-сплайсинг РНК у позвоночных» . Геномная биология и эволюция . 8 (3): 562–577. дои : 10.1093/gbe/evw025 . ПМЦ 4824033 . ПМИД 26966239 .
- ^ Райт Б.В., Моллой, член парламента, Яшке П.Р. (март 2022 г.). «Перекрывающиеся гены в природных и искусственно созданных геномах» . Обзоры природы. Генетика . 23 (3): 154–168. дои : 10.1038/s41576-021-00417-w . ПМЦ 8490965 . ПМИД 34611352 .
- ↑ Перейти обратно: Перейти обратно: а б Эдди С.Р. (декабрь 2001 г.). «Некодирующие РНК-гены и современный мир РНК». Обзоры природы. Генетика . 2 (12): 919–29. дои : 10.1038/35103511 . ПМИД 11733745 . S2CID 18347629 .
- ^ Крик Ф.Х., Барнетт Л., Бреннер С., Уоттс-Тобин Р.Дж. (декабрь 1961 г.). «Общая природа генетического кода белков». Природа . 192 (4809): 1227–32. Бибкод : 1961Natur.192.1227C . дои : 10.1038/1921227a0 . ПМИД 13882203 . S2CID 4276146 .
- ^ Крик Ф.Х. (октябрь 1962 г.). «Генетический код» . Научный американец . 207 (4). WH Freeman and Company: 66–74. Бибкод : 1962SciAm.207d..66C . дои : 10.1038/scientificamerican1062-66 . ПМИД 13882204 .
- ^ Вудсон С.А. (май 1998 г.). «Сглаживание перегибов: сплайсинг и трансляция у бактерий» . Гены и развитие . 12 (9): 1243–7. дои : 10.1101/gad.12.9.1243 . ПМИД 9573040 .
- ^ Джейкоб Ф. , Моно Дж. (июнь 1961 г.). «Генетические регуляторные механизмы синтеза белков». Журнал молекулярной биологии . 3 (3): 318–56. дои : 10.1016/S0022-2836(61)80072-7 . ПМИД 13718526 . S2CID 19804795 .
- ^ Кунин Е.В., Доля В.В. (январь 1993 г.). «Эволюция и таксономия РНК-вирусов с положительной цепью: последствия сравнительного анализа аминокислотных последовательностей». Критические обзоры по биохимии и молекулярной биологии . 28 (5): 375–430. дои : 10.3109/10409239309078440 . ПМИД 8269709 .
- ^ Воскресенье Е (2001). «Геномы РНК-вирусов». ЭЛС . дои : 10.1002/9780470015902.a0001488.pub2 . ISBN 978-0470016176 .
- ^ Доминго Э., Эскармис С., Севилья Н., Моя А., Елена С.Ф., Кер Дж. и др. (июнь 1996 г.). «Основные понятия эволюции РНК-вирусов» . Журнал ФАСЭБ . 10 (8): 859–64. дои : 10.1096/fasebj.10.8.8666162 . ПМИД 8666162 . S2CID 20865732 .
- ^ Мико I (2008). «Грегор Мендель и принципы наследования» . Знания о природном образовании . Научная таблица. 1 (1). Издательская группа «Природа»: 134.
- ^ Чиал Х (2008). «Менделевская генетика: закономерности наследования и одногенные нарушения» . Знания о природном образовании . Научная таблица. 1 (1). Издательская группа «Природа»: 63.
- ^ Маккарти Д., Миннер С., Бернштейн Х., Бернштейн С. (октябрь 1976 г.). «Скорость элонгации ДНК и распределение точек роста фага Т4 дикого типа и янтарного мутанта с задержкой ДНК». Журнал молекулярной биологии . 106 (4): 963–81. дои : 10.1016/0022-2836(76)90346-6 . ПМИД 789903 .
- ↑ Перейти обратно: Перейти обратно: а б Лобо I, Шоу К. (2008). «Открытие и типы генетического сцепления» . Знания о природном образовании . Научная таблица. 1 (1). Издательская группа «Природа»: 139.
- ^ Нахман М.В., Кроуэлл С.Л. (сентябрь 2000 г.). «Оценка скорости мутаций на нуклеотид у человека» . Генетика . 156 (1): 297–304. дои : 10.1093/генетика/156.1.297 . ПМЦ 1461236 . ПМИД 10978293 .
- ^ Роуч Дж.К., Глусман Дж., Смит А.Ф., Хафф К.Д., Хабли Р., Шеннон П.Т. и др. (апрель 2010 г.). «Анализ генетического наследования в семейном квартете методом полногеномного секвенирования» . Наука . 328 (5978): 636–9. Бибкод : 2010Sci...328..636R . дои : 10.1126/science.1186802 . ПМК 3037280 . ПМИД 20220176 .
- ^ Дрейк Дж.В., Чарльзуорт Б., Чарльзуорт Д., Кроу Дж.Ф. (апрель 1998 г.). «Скорость спонтанных мутаций» . Генетика . 148 (4): 1667–86. дои : 10.1093/генетика/148.4.1667 . ПМК 1460098 . ПМИД 9560386 .
- ^ Пьериц, Рид Э., Брюс Р. Корф и Уэйн В. Гроди, ред. Принципы и практика медицинской генетики и геномики Эмери и Римоэна: основы. Академик Пресс, 2018.
- ^ «Какие виды генных мутаций возможны?» . Домашний справочник по генетике . Национальная медицинская библиотека США. 11 мая 2015 года . Проверено 19 мая 2015 г.
- ^ Эндрюс, Калифорния (2010). «Естественный отбор, генетический дрейф и поток генов не действуют изолированно в естественных популяциях» . Знания о природном образовании . Научная таблица. 3 (10). Издательская группа «Природа»: 5.
- ^ Паттерсон С. (ноябрь 1988 г.). «Гомология в классической и молекулярной биологии» . Молекулярная биология и эволюция . 5 (6): 603–25. doi : 10.1093/oxfordjournals.molbev.a040523 . ПМИД 3065587 .
- ^ Граур Д (2016). Молекулярная и геномная эволюция . Сандерленд, Массачусетс (США): Sinauer Associates, Inc. ISBN 9781605354699 .
- ^ Граур Д (2016). Молекулярная и геномная эволюция . Сандерленд, Массачусетс (США): Sinauer Associates, Inc. ISBN 9781605354699 .
- ^ Дженсен Р.А. (2001). «Ортологи и паралоги – нам нужно разобраться» . Геномная биология . 2 (8): ВЗАИМОДЕЙСТВИЯ1002. doi : 10.1186/gb-2001-2-8-interactions1002 . ПМК 138949 . ПМИД 11532207 .
- ^ Студер Р.А., Робинсон-Речави М (май 2009 г.). «Насколько мы можем быть уверены в том, что ортологи похожи, а паралоги различаются?» . Тенденции в генетике . 25 (5): 210–6. дои : 10.1016/j.tig.2009.03.004 . ПМИД 19368988 .
- ^ Альтенхофф А.М., Штудер Р.А., Робинсон-Речави М., Дессимоз С. (2012). «Разрешение гипотезы об ортологах: ортологи имеют тенденцию быть слабо, но значительно более похожими по функциям, чем паралоги» . PLOS Вычислительная биология . 8 (5): e1002514. Бибкод : 2012PLSCB...8E2514A . дои : 10.1371/journal.pcbi.1002514 . ПМК 3355068 . ПМИД 22615551 .
- ↑ Перейти обратно: Перейти обратно: а б Герзони Д., Маклизат А. (ноябрь 2011 г.). «Происхождение человеческих генов de novo» . ПЛОС Генетика . 7 (11): e1002381. дои : 10.1371/journal.pgen.1002381 . ПМК 3213182 . ПМИД 22102832 .
- ^ Реамс А.Б., Рот-младший (февраль 2015 г.). «Механизмы дупликации и амплификации генов» . Перспективы Колд-Спринг-Харбор в биологии . 7 (2): а016592. doi : 10.1101/cshperspect.a016592 . ПМЦ 4315931 . ПМИД 25646380 .
- ^ Демут Дж.П., Де Би Т., Стажич Дж.Е., Кристианини Н., Хан М.В. (декабрь 2006 г.). «Эволюция семейств генов млекопитающих» . ПЛОС ОДИН . 1 (1): е85. Бибкод : 2006PLoSO...1...85D . дои : 10.1371/journal.pone.0000085 . ПМЦ 1762380 . ПМИД 17183716 .
- ^ Ноулз Д.Г., Маклизат А. (октябрь 2009 г.). «Недавнее происхождение генов, кодирующих человеческие белки», de novo . Геномные исследования . 19 (10): 1752–9. дои : 10.1101/гр.095026.109 . ПМЦ 2765279 . ПМИД 19726446 .
- ^ Ву Д.Д., Ирвин Д.М., Чжан Ю.П. (ноябрь 2011 г.). «Происхождение de novo генов, кодирующих человеческие белки» . ПЛОС Генетика . 7 (11): e1002379. дои : 10.1371/journal.pgen.1002379 . ПМЦ 3213175 . ПМИД 22102831 .
- ^ МакЛисахт А., Герцони Д. (сентябрь 2015 г.). «Новые гены из некодирующей последовательности: роль генов, кодирующих белки de novo, в эволюционных инновациях эукариот» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 370 (1678): 20140332. doi : 10.1098/rstb.2014.0332 . ПМЦ 4571571 . ПМИД 26323763 .
- ^ Неме Р., Таутц Д. (февраль 2013 г.). «Филогенетические закономерности появления новых генов подтверждают модель частой эволюции de novo» . БМК Геномика . 14 (1): 117. дои : 10.1186/1471-2164-14-117 . ПМК 3616865 . ПМИД 23433480 .
- ^ Treangen TJ, Rocha EP (январь 2011 г.). «Горизонтальный перенос, а не дупликация, стимулирует расширение семейств белков у прокариот» . ПЛОС Генетика . 7 (1): e1001284. дои : 10.1371/journal.pgen.1001284 . ПМК 3029252 . ПМИД 21298028 .
- ^ Охман Х., Лоуренс Дж.Г., Гройсман Э.А. (май 2000 г.). «Боковой перенос генов и природа бактериальных инноваций». Природа . 405 (6784): 299–304. Бибкод : 2000Natur.405..299O . дои : 10.1038/35012500 . ПМИД 10830951 . S2CID 85739173 .
- ^ Килинг П.Дж., Палмер Дж.Д. (август 2008 г.). «Горизонтальный перенос генов в эволюции эукариот». Обзоры природы. Генетика . 9 (8): 605–18. дои : 10.1038/nrg2386 . ПМИД 18591983 . S2CID 213613 .
- ^ Шёнкнехт Г., Чен В.Х., Тернес С.М., Барбье Г.Г., Шреста Р.П., Станке М. и др. (март 2013 г.). «Перенос генов от бактерий и архей облегчил эволюцию экстремофильных эукариот» . Наука . 339 (6124): 1207–10. Бибкод : 2013Sci...339.1207S . дои : 10.1126/science.1231707 . ПМИД 23471408 . S2CID 5502148 .
- ^ Ридли, М. (2006). Геном . Нью-Йорк, штат Нью-Йорк: Harper Perennial. ISBN 0-06-019497-9
- ^ Банерджи С., Бхандари П., Вудхаус М., Сен Т.З., Уайз Р.П., Андорф К.М. (апрель 2021 г.). «FINDER: автоматизированный пакет программного обеспечения для аннотирования эукариотических генов на основе данных RNA-Seq и связанных с ними белковых последовательностей» . БМК Биоинформатика . 44 (9): е89. дои : 10.1186/s12859-021-04120-9 . ПМК 8056616 . ПМИД 33879057 .
- ^ Уотсон, Дж.Д., Бейкер Т.А., Белл С.П., Ганн А., Левин М., Лосик Р. (2004). «Ch9-10», Молекулярная биология гена, 5-е изд., Писон Бенджамин Каммингс; ЦШЛ Пресс.
- ^ «Integr8 – Статистика генома A.thaliana» .
- ^ «Понимание основ» . Проект «Геном человека» . Проверено 26 апреля 2015 г.
- ^ «Письмо о выпуске WS227» . Червячная база. 10 августа 2011 года. Архивировано из оригинала 28 ноября 2013 года . Проверено 19 ноября 2013 г.
- ↑ Перейти обратно: Перейти обратно: а б Ю Дж., Ху С., Ван Дж., Вонг Г.К., Ли С., Лю Б. и др. (апрель 2002 г.). «Проект последовательности генома риса (Oryza sativa L. ssp. indica)». Наука . 296 (5565): 79–92. Бибкод : 2002Sci...296...79Y . дои : 10.1126/science.1068037 . ПМИД 11935017 . S2CID 208529258 .
- ^ Андерсон С., Банкир А.Т., Баррелл Б.Г., де Брёйн М.Х., Коулсон А.Р., Друэн Дж. и др. (апрель 1981 г.). «Последовательность и организация митохондриального генома человека». Природа . 290 (5806): 457–65. Бибкод : 1981Natur.290..457A . дои : 10.1038/290457a0 . ПМИД 7219534 . S2CID 4355527 .
- ^ Адамс М.Д., Селникер С.Е., Холт Р.А., Эванс К.А., Гокейн Дж.Д., Аманатидес П.Г. и др. (март 2000 г.). «Последовательность генома Drosophila melanogaster». Наука . 287 (5461): 2185–95. Бибкод : 2000Sci...287.2185. . CiteSeerX 10.1.1.549.8639 . дои : 10.1126/science.287.5461.2185 . ПМИД 10731132 .
- ^ Пертеа М., Зальцберг С.Л. (2010). «Между курицей и виноградом: оценка количества генов человека» . Геномная биология . 11 (5): 206. doi : 10.1186/gb-2010-11-5-206 . ПМК 2898077 . ПМИД 20441615 .
- ^ Белый В.А., Левин А.Дж., Скалка А.М. (декабрь 2010 г.). «Последовательности предковых одноцепочечных ДНК-вирусов в геномах позвоночных: парвовирусам и цирковирусам более 40–50 миллионов лет» . Журнал вирусологии . 84 (23): 12458–62. дои : 10.1128/JVI.01789-10 . ПМЦ 2976387 . ПМИД 20861255 .
- ^ Флорес Р., Ди Серио Ф, Эрнандес К (февраль 1997 г.). «Вироиды: некодирующие геномы». Семинары по вирусологии . 8 (1): 65–73. дои : 10.1006/smvy.1997.0107 .
- ^ Зонневельд Б.Дж. (2010). «Новые рекордсмены по максимальному размеру генома эвдикотов и однодольных растений» . Журнал ботаники . 2010 : 1–4. дои : 10.1155/2010/527357 .
- ^ Перес-Иратчета С., Палидвор Г., Андраде-Наварро М.А. (декабрь 2007 г.). «На пути к завершению протеома Земли» . Отчеты ЭМБО . 8 (12): 1135–41. дои : 10.1038/sj.embor.7401117 . ПМК 2267224 . ПМИД 18059312 .
- ^ Мюллер Х.Дж. (1966). «Генный материал как инициатор и организующая основа жизни» . Американский натуралист . 100 (915): 493–517. дои : 10.1086/282445 . JSTOR 2459205 . S2CID 84202145 .
- ^ Оно С. (1972). «В нашем геноме так много «мусорной» ДНК». Брукхейвенские симпозиумы по биологии . 23 : 366–370. ПМИД 5065367 .
- ^ Хатье К., Мюльхаузен С., Симм Д., Киллмар М. (2019). «Геном человека, кодирующий белок: аннотирование высоко висящих плодов» . Биоэссе . 41 (11): 1900066. doi : 10.1002/bies.201900066 . ПМИД 31544971 . S2CID 202732556 .
- ^ Шулер Г.Д., Богуски М.С. , Стюарт Е.А., Штейн Л.Д., Гьяпай Г., Райс К. и др. (октябрь 1996 г.). «Геновая карта генома человека». Наука . 274 (5287): 540–6. Бибкод : 1996Sci...274..540S . дои : 10.1126/science.274.5287.540 . ПМИД 8849440 . S2CID 22619 .
- ^ Чи КР (октябрь 2016 г.). «Темная сторона человеческого генома» . Природа . 538 (7624): 275–277. Бибкод : 2016Natur.538..275C . дои : 10.1038/538275а . ПМИД 27734873 .
- ^ «Сборка человека и аннотация генов» . Ансамбль . 2022 . Проверено 28 февраля 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б Хатчисон К.А., Чуанг Р.Ю., Носков В.Н., Асад-Гарсия Н., Диринк Т.Дж., Эллисман М.Х. и др. (март 2016 г.). «Дизайн и синтез минимального бактериального генома» . Наука . 351 (6280): аад6253. Бибкод : 2016Sci...351.....H . дои : 10.1126/science.aad6253 . ПМИД 27013737 .
- ^ Гласс Дж.И., Асад-Гарсия Н., Альперович Н., Юсеф С., Льюис М.Р., Маруф М. и др. (январь 2006 г.). «Основные гены минимальной бактерии» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (2): 425–30. Бибкод : 2006PNAS..103..425G . дои : 10.1073/pnas.0510013103 . ПМЦ 1324956 . ПМИД 16407165 .
- ^ Гердес С.Ю., Шолле, М.Д., Кэмпбелл Дж.В., Балажи Г., Равас Э., Догерти, М.Д. и др. (октябрь 2003 г.). «Экспериментальное определение и анализ на системном уровне основных генов Escherichia coli MG1655» . Журнал бактериологии . 185 (19): 5673–84. дои : 10.1128/jb.185.19.5673-5684.2003 . ЧВК 193955 . ПМИД 13129938 .
- ^ Баба Т., Ара Т., Хасегава М., Такай Ю., Окумура Ю., Баба М. и др. (2006). «Создание мутантов Escherichia coli K-12 с нокаутом одного гена в рамке: коллекция Кейо» . Молекулярная системная биология . 2 : 2006.0008. дои : 10.1038/msb4100050 . ПМК 1681482 . ПМИД 16738554 .
- ↑ Перейти обратно: Перейти обратно: а б Юхас М., Ройсс Д.Р., Жу Б., Commichau FM (ноябрь 2014 г.). «Основные гены Bacillus subtilis и Escherichia coli и минимальные клеточные фабрики после десятилетия геномной инженерии» . Микробиология . 160 (Часть 11): 2341–2351. дои : 10.1099/mic.0.079376-0 . ПМИД 25092907 .
- ^ Ту З, Ван Л, Сюй М, Чжоу Х, Чен Т, Сунь Ф (февраль 2006 г.). «Дальнейшее понимание генов болезней человека путем сравнения с генами домашнего хозяйства и другими генами» . БМК Геномика . 7:31 . дои : 10.1186/1471-2164-7-31 . ПМЦ 1397819 . ПМИД 16504025 .
- ^ Георгий Б., Войт Б.Ф., Бьюкан М. (май 2013 г.). «От мыши к человеку: эволюционный геномный анализ человеческих ортологов основных генов» . ПЛОС Генетика . 9 (5): e1003484. дои : 10.1371/journal.pgen.1003484 . ПМЦ 3649967 . ПМИД 23675308 .
- ^ Айзенберг Э., Леванон Э.Ю. (октябрь 2013 г.). «Человеческие гены домашнего хозяйства, еще раз». Тенденции в генетике . 29 (10): 569–74. дои : 10.1016/j.tig.2013.05.010 . ПМИД 23810203 .
- ^ Амстердам А., Хопкинс Н. (сентябрь 2006 г.). «Стратегии мутагенеза у рыбок данио для выявления генов, участвующих в развитии и заболеваниях». Тенденции в генетике . 22 (9): 473–8. дои : 10.1016/j.tig.2006.06.011 . ПМИД 16844256 .
- ^ «О ВГНК» . База данных HGNC названий генов человека . Комитет по генной номенклатуре Хьюго. Архивировано из оригинала 26 марта 2023 года . Проверено 14 мая 2015 г.
- ^ Коэн С.Н., Чанг AC (май 1973 г.). «Рециркуляция и автономная репликация фрагмента ДНК R-фактора в трансформантах Escherichia coli» . Труды Национальной академии наук Соединенных Штатов Америки . 70 (5): 1293–7. Бибкод : 1973PNAS...70.1293C . дои : 10.1073/pnas.70.5.1293 . ПМЦ 433482 . ПМИД 4576014 .
- ^ Эсвелт К.М., Ван Х.Х. (2013). «Геномная инженерия для системной и синтетической биологии» . Молекулярная системная биология . 9 (1): 641. doi : 10.1038/msb.2012.66 . ПМЦ 3564264 . ПМИД 23340847 .
- ^ Тан В.С., Карлсон Д.Ф., Уолтон М.В., Фаренкруг СК, Хакетт П.Б. (2012). «Точное редактирование геномов крупных животных». Достижения в области генетики, том 80 . Том. 80. стр. 37–97. дои : 10.1016/B978-0-12-404742-6.00002-8 . ISBN 9780124047426 . ПМЦ 3683964 . ПМИД 23084873 .
- ^ Пухта Х, Фаузер Ф (2013). «Нацеливание на гены растений: 25 лет спустя» . Международный журнал биологии развития . 57 (6–8): 629–37. дои : 10.1387/ijdb.130194hp . ПМИД 24166445 .
- ^ Ран Ф.А., Сюй П.Д., Райт Дж., Агарвала В., Скотт Д.А., Чжан Ф. (ноябрь 2013 г.). «Геномная инженерия с использованием системы CRISPR-Cas9» . Протоколы природы . 8 (11): 2281–2308. дои : 10.1038/nprot.2013.143 . ПМЦ 3969860 . ПМИД 24157548 .
- ^ Киттлсон Дж. Т., Ву Г. К., Андерсон Дж. К. (август 2012 г.). «Успехи и неудачи модульной генной инженерии». Современное мнение в области химической биологии . 16 (3–4): 329–36. дои : 10.1016/j.cbpa.2012.06.009 . ПМИД 22818777 .
- ^ Берг П., Мерц Дж.Э. (январь 2010 г.). «Личные размышления о происхождении и появлении технологии рекомбинантной ДНК» . Генетика . 184 (1): 9–17. дои : 10.1534/genetics.109.112144 . ПМЦ 2815933 . ПМИД 20061565 .
- ^ Остин С.П., Бэтти Дж.Ф., Брэдли А., Букан М., Капеччи М., Коллинз Ф.С. и др. (сентябрь 2004 г.). «Проект нокаутирующей мыши» . Природная генетика . 36 (9): 921–4. дои : 10.1038/ng0904-921 . ПМК 2716027 . ПМИД 15340423 .
- ^ Гуань С., Е С., Ян Х, Гао Дж. (февраль 2010 г.). «Обзор текущих крупномасштабных усилий по нокауту мышей». Бытие . 48 (2): 73–85. дои : 10.1002/dvg.20594 . ПМИД 20095055 . S2CID 34470273 .
- ^ Дэн С. (октябрь 2007 г.). «В честь вручения Нобелевской премии доктору Марио Р. Капечки» . Международный журнал биологических наук . 3 (7): 417–9. дои : 10.7150/ijbs.3.417 . ПМК 2043165 . ПМИД 17998949 .
Источники [ править ]
- Основной учебник
- Альбертс Б. , Джонсон А., Льюис Дж. , Рафф М. , Робертс К., Уолтер П. (2002). Молекулярная биология клетки (Четвертое изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 . – Учебник по молекулярной биологии доступен бесплатно в Интернете на сайте NCBI Bookshelf.
Дальнейшее чтение [ править ]
- Уотсон Дж.Д. , Бейкер Т.А. , Белл С.П., Ганн А., Левин М. , Лосик Р. (2013). Молекулярная биология гена (7-е изд.). Бенджамин Каммингс. ISBN 978-0-321-90537-6 .
- Докинз Р. (1990). Эгоистичный ген . Издательство Оксфордского университета. ISBN 978-0-19-286092-7 .
- Ридли М. (1999). Геном: автобиография вида в 23 главах . Четвертая власть. ISBN 978-0-00-763573-3 .
- Браун Т. (2002). Геномы (2-е изд.). Нью-Йорк: Вили-Лисс. ISBN 978-0-471-25046-3 . ПМИД 20821850 .
Внешние ссылки [ править ]
- База данных сравнительной токсикогеномики
- ДНК с самого начала – учебник по генам и ДНК
- Ген - база данных генов с возможностью поиска.
- Гены - журнал открытого доступа
- IDconverter – конвертирует идентификаторы генов между общедоступными базами данных.
- iHOP – информация, связанная гиперссылками на белки
- TranscriptomeBrowser – анализ профиля экспрессии генов
- Утилита именования белков — база данных для идентификации и исправления неверных названий генов.
- IMPC (Международный консорциум фенотипирования мышей) - Энциклопедия функции генов млекопитающих.
- Global Genes Project – ведущая некоммерческая организация, поддерживающая людей, живущих с генетическими заболеваниями.
- Обозреватель потоков кодирования , Природа