Бактериофаг MS2
| Эмесвирус зиндери | |
|---|---|
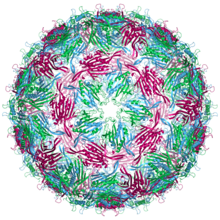 | |
| бактериофага MS2 Структура капсида . Три квазиэквивалентных конформера отмечены синим (цепь а), зеленым (цепь б) и пурпурным (цепь в) | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Ленарвирикота |
| Сорт: | Левивирицеты |
| Заказ: | Норзивирусы |
| Семья: | Фиерсвирусиды |
| Род: | Эмесвирус |
| Разновидность: | Эмесвирус зиндери |
Бактериофаг MS2 ( Emesvirus zinderi ), обычно называемый MS2, представляет собой икосаэдрический одноцепочечный РНК- вирус с положительным смыслом, который инфицирует бактерию Escherichia coli и других представителей Enterobacteriaceae . [1] MS2 является членом семейства близкородственных бактериальных вирусов, которое включает бактериофаг f2 , бактериофаг Qβ , R17 и GA. [2]
Он небольшой и содержит белок созревания, белок оболочки и геномную РНК. Он также имеет один из самых маленьких известных геномов, кодирующий четыре белка.
Жизненный цикл MS2 включает заражение бактерий фактором фертильности, что позволяет вирусу прикрепляться к волоскам, хотя механизм, с помощью которого РНК вируса попадает в бактерию, остается неизвестным. Оказавшись внутри, вирусная РНК начинает функционировать как информационная РНК для производства вирусных белков. MS2 реплицирует свой геном с плюсовой цепью, создавая минус-цепь РНК в качестве матрицы. Затем вирус собирается, и бактериальная клетка лизирует, высвобождая новые вирусы.
Вирус был выделен в 1961 году, а его геном был первым полностью секвенирован в 1976 году, что обеспечило решающее понимание генетических кодов. В практических приложениях структурные компоненты MS2 использовались для обнаружения РНК в живых клетках. Вирус также исследуется на предмет потенциального использования в доставке лекарств, визуализации опухолей и сборе света. Более того, из-за структурного сходства с норовирусами , предпочтительных условий пролиферации и отсутствия патогенности для человека MS2 служит заменой в исследованиях передачи норовирусных заболеваний.
Вирусология
[ редактировать ]Геном
[ редактировать ]
| Ген | Размер | Генный продукт | аа |
|---|---|---|---|
| вместе с (MS2g1) | 1487 нт | созревание | 393 |
| КП (МС2г2) | 510 нт | белок оболочки | 130 |
| свет (MS2g3) | 295 нт | лизисный белок | 75 |
| представитель (MS2g4) | 2055 нт | репликаза РНК , | 545 |
Геном MS2 — один из самых маленьких из известных: он состоит из 3569 нуклеотидов одноцепочечной РНК. [3] Он кодирует всего четыре белка: белок созревания ( А -белок), белок лизиса ( lys ), белок оболочки ( cp ) и белок репликазы ( rep ). [1] Ген, кодирующий lys, перекрывает как 3'-конец верхнего гена ( cp ), так и 5'-конец нижнего гена ( rep ), и был одним из первых известных примеров перекрывающихся генов . Геном положительно-цепочечной РНК служит информационной РНК и транслируется при удалении вируса внутри клетки-хозяина. Хотя все четыре белка кодируются одной и той же информационной/вирусной РНК, не все они экспрессируются на одинаковых уровнях.
Структура
[ редактировать ]
MS2 (вирусная частица) имеет диаметр около 27 нм. вирион По данным электронной микроскопии, [4] Он состоит из одной копии белка созревания и 180 копий белка оболочки (организованных в виде 90 димеров), организованных в икосаэдрическую оболочку с числом триангуляции Т=3 , защищающую находящуюся внутри геномную РНК. [5] Вирион имеет изоэлектрическую точку (pI) 3,9. [6]
Структура белка оболочки представляет собой пятинитевой β-лист с двумя α-спиралями и шпилькой . Когда капсид собран, спирали и шпилька обращены к внешней стороне частицы, а β-лист обращен внутрь. [7]
Жизненный цикл
[ редактировать ]MS2 заражает кишечные бактерии, несущие фактор фертильности (F) , — плазмиду которая позволяет клеткам служить донорами ДНК при бактериальной конъюгации . Гены плазмиды F определяют белки F- пилуса , который включает белок F-пилин, который служит вирусным рецептором. MS2 прикрепляется к F-пилину на стороне пилуса с помощью единственного белка созревания.
Как только вирусная РНК попадает в клетку, она начинает действовать как информационная РНК для производства фаговых белков. Ген самого распространенного белка — белка оболочки — можно немедленно транслировать. Начало трансляции гена репликазы обычно скрыто во вторичной структуре РНК, но может временно открываться, когда рибосомы проходят через ген белка оболочки. Трансляция репликазы также прекращается после образования большого количества белка оболочки; » РНК Димеры белка оболочки связывают и стабилизируют «операторную шпильку , блокируя запуск репликазы. Начало гена белка созревания доступно в реплицируемой РНК, но скрыто во вторичной структуре РНК в завершенной РНК MS2; это гарантирует трансляцию лишь очень небольшого количества копий белка созревания на РНК. Наконец, ген белка лизиса может быть инициирован только рибосомами, которые завершили трансляцию гена белка оболочки и «соскользнули обратно» к началу гена белка лизиса, с частотой примерно 5%. [1]

Репликация генома MS2 с плюсовой цепью требует синтеза комплементарной минус-цепи РНК, которую затем можно использовать в качестве матрицы для синтеза новой плюс-цепи РНК. Репликация MS2 изучена гораздо хуже, чем репликация близкородственного бактериофага Qβ , отчасти потому, что репликазу MS2 было трудно выделить, но она, вероятно, аналогична. [1]
Считается, что образование вириона инициируется связыванием белка созревания с РНК MS2; фактически комплекс созревания белка и РНК является инфекционным. Сборка икосаэдрической оболочки или капсида из белков оболочки может происходить в отсутствие РНК; однако сборка капсида зарождается за счет связывания димера белка оболочки со шпилькой оператора, и сборка происходит при гораздо более низких концентрациях белка оболочки, когда присутствует РНК MS2. [1]
Лизис бактерий и высвобождение вновь образовавшихся вирионов происходит при накоплении достаточного количества лизисного белка. Белок Lysis (L) образует поры в цитоплазматической мембране, что приводит к потере мембранного потенциала и разрушению клеточной стенки . [1] Известно, что лизирующий белок связывается с DnaJ через важный остаток P330. [8] Дипептидный мотив LS на белке L обнаружен во всем роде Levivirus и, по-видимому, важен для лизисной активности, хотя их различное расположение позволяет предположить, что они развивались независимо. [9]
MS2 по истории науки и использования
[ редактировать ]В 1961 году Элвин Джон Кларк выделил MS2 и признал его РНК-содержащим фагом, очень похожим на бактериофаг f2 . [10]
В 1976 году геном MS2 стал первым полностью секвенированным геномом. [3] Это было достигнуто Уолтером Фирсом и его командой, основываясь на их более ранней вехе в 1972 году, когда был полностью секвенирован первый ген — оболочечный белок MS2. [11] Эти последовательности были определены на уровне РНК. [12] Первой попыткой статистического анализа генома MS2 стал поиск закономерностей в нуклеотидной последовательности. Было идентифицировано несколько некодирующих последовательностей, однако на момент этого исследования (1979 г.) функции некодирующих паттернов были неизвестны. [13]
С 1998 года [14] Шпилька-оператор MS2 и белок оболочки нашли применение при обнаружении РНК в живых клетках (см. Мечение MS2 ). MS2 и другие вирусные капсиды в настоящее время также исследуются в качестве агентов для доставки лекарств, опухолей визуализации и сбора света. [15]
MS2 из-за его структурного сходства с норовирусами , сходных оптимальных условий пролиферации и непатогенности для человека использовался в качестве заменителя норовирусов в исследованиях передачи заболеваний. [16]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж ван Дуин Дж, Царева Н (2006). «Одноцепочечные РНК-фаги. Глава 15». В Календаре RL (ред.). Бактериофаги (второе изд.). Издательство Оксфордского университета. стр. 175–196. ISBN 978-0195148503 .
- ^ Ни Ч.З., Уайт К.А., Митчелл Р.С., Викершам Дж., Кодандапани Р., Пибоди Д.С., Эли К.Р. (декабрь 1996 г.). «Кристаллическая структура белка оболочки бактериофага GA: модель несобранного димера» . Белковая наука . 5 (12): 2485–93. дои : 10.1002/pro.5560051211 . ПМК 2143325 . ПМИД 8976557 .
- ^ Перейти обратно: а б Фирс В., Контрерас Р., Дюринк Ф., Хагеман Г., Изерентант Д., Меррегарт Дж., Мин Джоу В., Молеманс Ф., Раймакерс А., Ван ден Берге А., Волкарт Г., Изебарт М. (апрель 1976 г.). «Полная нуклеотидная последовательность РНК бактериофага MS2: первичная и вторичная структура гена репликазы». Природа . 260 (5551): 500–7. Бибкод : 1976Natur.260..500F . дои : 10.1038/260500a0 . ПМИД 1264203 . S2CID 4289674 .
- ^ Штраус Дж. Х., Зиншаймер Р. Л. (июль 1963 г.). «Очистка и свойства бактериофага MS2 и его рибонуклеиновой кислоты». Журнал молекулярной биологии . 7 : 43–54. дои : 10.1016/S0022-2836(63)80017-0 . ПМИД 13978804 .
- ^ Валегард К., Лильяс Л., Фридборг К., Унге Т. (май 1990 г.). «Трехмерная структура бактериального вируса MS2». Природа . 345 (6270): 36–41. Бибкод : 1990Natur.345...36V . дои : 10.1038/345036a0 . ПМИД 2330049 . S2CID 2803228 .
- ^ Дауд С.Е., Пиллаи С.Д., Ван С., Корапчиоглу М.Ю. (1998). «Определение специфического влияния изоэлектрической точки и размера вируса на адсорбцию и транспорт вируса через песчаные почвы» . Прил. Окружающая среда. Микробиол. 64 (2): 405–410. Бибкод : 1998ApEnM..64..405D . дои : 10.1128/аэм.64.2.405-410.1998 . ПМК 106058 . ПМИД 9464373 .
- ^ Голмохаммади Р., Валегард К., Фридборг К., Лильяс Л. (декабрь 1993 г.). «Уточненная структура бактериофага MS2 при разрешении 2,8 А». Журнал молекулярной биологии . 234 (3): 620–39. дои : 10.1006/jmbi.1993.1616 . ПМИД 8254664 .
- ^ Чамакура К.Р., Тран Дж.С., Янг Р. (июнь 2017 г.). «Лизис MS2 Escherichia coli зависит от шаперона-хозяина DnaJ» . Журнал бактериологии . 199 (12). дои : 10.1128/JB.00058-17 . ПМК 5446614 . ПМИД 28396351 .
- ^ Чамакура К.Р., Эдвардс ГБ, Янг Р. (июль 2017 г.). «Мутационный анализ белка L лизиса MS2» . Микробиология . 163 (7): 961–969. дои : 10.1099/mic.0.000485 . ПМЦ 5775895 . ПМИД 28691656 .
- ^ «Национальная академия наук: тезисы докладов, представленных на осеннем собрании 29 октября, Ла-Хойя, Калифорния, 30 октября — 1 ноября 1961 года, Лос-Анджелес». Наука . 134 (3488): 1425–37. Ноябрь 1961 г. Бибкод : 1961Sci...134.1425. . дои : 10.1126/science.134.3488.1425 . ПМИД 17795773 .
- ^ Мин Джоу В., Хагеман Г., Изеберт М., Фирс В. (май 1972 г.). «Нуклеотидная последовательность гена, кодирующего белок оболочки бактериофага MS2». Природа . 237 (5350): 82–8. Бибкод : 1972Natur.237...82J . дои : 10.1038/237082a0 . ПМИД 4555447 . S2CID 4153893 .
- ^ Сэнгер Ф., генеральный директор Air, Баррелл Б.Г., Браун Н.Л., Коулсон А.Р., Фиддес К.А., Хатчисон К.А., Слокомб П.М., Смит М. (февраль 1977 г.). «Нуклеотидная последовательность ДНК бактериофага phi X174». Природа . 265 (5596): 687–95. Бибкод : 1977Natur.265..687S . дои : 10.1038/265687a0 . ПМИД 870828 . S2CID 4206886 .
- ^ Эриксон Дж.В., Альтман Г.Г. (апрель 1979 г.). «Поиск закономерностей в нуклеотидной последовательности генома MS2». Журнал математической биологии . 7 (3): 219–30. дои : 10.1007/BF00275725 . S2CID 85199492 .
- ^ Бертран Э., Чартран П., Шефер М., Шеной С.М., Сингер Р.Х., Лонг Р.М. (октябрь 1998 г.). «Локализация частиц мРНК ASH1 в живых дрожжах» . Молекулярная клетка . 2 (4): 437–45. дои : 10.1016/S1097-2765(00)80143-4 . ПМИД 9809065 .
- ^ Глазго Дж., Таллман-Эрчек Д. (июль 2014 г.). «Производство и применение инженерных вирусных капсидов». Прикладная микробиология и биотехнология . 98 (13): 5847–58. дои : 10.1007/s00253-014-5787-3 . ПМИД 24816622 . S2CID 6212583 .
- ^ Фокс М (8 сентября 2014 г.). «Исследование показало, что вирусы распространяются в офисе как сумасшедшие» . Сегодняшнее шоу .
Внешние ссылки
[ редактировать ]- Полный геном (также изолирует R17 , DL16 и J20 )