Перекрывающийся ген
( Перекрывающийся ген или OLG ) [1] [2] Ген , экспрессируемая нуклеотидная последовательность которого частично перекрывается с экспрессируемой нуклеотидной последовательностью другого гена. [3] Таким образом, нуклеотидная последовательность может вносить вклад в функцию одного или нескольких генных продуктов . Перекрывающиеся гены присутствуют и являются фундаментальной особенностью как клеточных, так и вирусных геномов . [2] Текущее определение перекрывающегося гена значительно различается у эукариот, прокариот и вирусов. [2] У прокариот и вирусов перекрытие должно происходить между кодирующими последовательностями , а не между транскриптами мРНК , и определяется, когда эти кодирующие последовательности имеют общий нуклеотид либо на одной, либо на противоположных цепях. У эукариот перекрытие генов почти всегда определяется как перекрытие транскриптов мРНК. В частности, перекрытие генов у эукариот определяется, когда по крайней мере один нуклеотид является общим между границами первичных транскриптов мРНК двух или более генов, так что мутация основания ДНК в любой точке перекрывающейся области может повлиять на транскрипты всех генов. вовлеченный. Это определение включает 5'- и 3'- нетранслируемые области (UTR) наряду с интронами .
Оверпринтинг относится к типу перекрытия, при котором вся или часть последовательности одного гена считывается в альтернативной рамке считывания из другого гена в том же локусе . [4] Считается, что альтернативные открытые рамки считывания (ORF) создаются в результате критических нуклеотидных замен внутри экспрессируемого ранее существовавшего гена, который можно заставить экспрессировать новый белок, сохраняя при этом функцию исходного гена. [5] Было высказано предположение, что оверпринтинг является механизмом появления de novo новых генов из существующих последовательностей, либо старых генов, либо ранее некодирующих областей генома. [6] Считается, что большинство перекрывающихся генов или генов, чьи экспрессируемые нуклеотидные последовательности частично перекрываются друг с другом, возникли частично благодаря этому механизму, предполагая, что каждое перекрытие состоит из одного предкового гена и одного нового гена. [7] Впоследствии считается, что оверпринтинг также является источником новых белков, поскольку белки de novo, кодируемые этими новыми генами, обычно не имеют удаленных гомологов в базах данных. [8] Гены с надпечаткой являются особенно распространенной чертой геномной организации вирусов, которая, вероятно, значительно увеличивает количество потенциально экспрессируемых генов из небольшого набора вирусной генетической информации. [9] Вполне вероятно, что оверпринтинг ответственен за создание вирусами множества новых белков в ходе их эволюционной истории .
Классификация
[ редактировать ]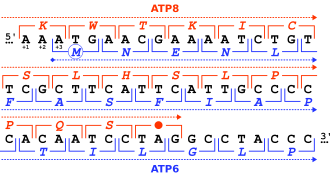
Гены могут перекрываться по-разному, и их можно классифицировать по их положению относительно друг друга. [3] [11] [12] [13] [14]
- Однонаправленное или тандемное перекрытие: 3'- конец одного гена перекрывается с 5'- концом другого гена на той же цепи. Такое расположение можно обозначить обозначением → →, где стрелки указывают рамку считывания от начала до конца.
- Конвергентное или перекрытие концов : 3'- концы двух генов перекрываются на противоположных цепях. Это можно записать как → ←.
- Дивергентное или хвостовое перекрытие: 5'- концы двух генов перекрываются на противоположных цепях. Это можно записать как ← →.
Перекрывающиеся гены также можно классифицировать по фазам , которые описывают их относительные рамки считывания : [3] [11] [12] [13] [14]
- Синфазное перекрытие происходит, когда общие последовательности используют одну и ту же рамку считывания. Это также известно как «фаза 0». Однонаправленные гены с перекрытием фазы 0 не считаются отдельными генами, а скорее альтернативными стартовыми сайтами одного и того же гена.
- Несинфазное перекрытие происходит, когда общие последовательности используют разные рамки считывания. Это может произойти на «фазе 1» или «фазе 2», в зависимости от того, смещены ли рамки считывания на 1 или 2 нуклеотида. Поскольку длина кодона составляет три нуклеотида, смещение на три нуклеотида представляет собой синфазный кадр фазы 0.
Исследования перекрывающихся генов позволяют предположить, что их эволюцию можно обобщить в двух возможных моделях. [4] В одной модели два белка, кодируемые соответствующими перекрывающимися генами, развиваются под одинаковым давлением отбора . Белки и область перекрытия высококонсервативны, когда аминокислот предпочтение отдается сильному отбору против замены . Предполагается, что перекрывающиеся гены развиваются в условиях строгих ограничений, поскольку замена одного нуклеотида способна изменить структуру и функцию двух белков одновременно. Исследование вируса гепатита B (HBV), геном ДНК которого содержит множество перекрывающихся генов, показало, что среднее количество синонимичных нуклеотидных замен на сайт в перекрывающихся кодирующих областях было значительно ниже, чем в неперекрывающихся областях. [15] То же исследование показало, что некоторые из этих перекрывающихся областей и их белков могут значительно отличаться от оригинала при слабом отборе против замены аминокислот. Например, спейсерный . домен полимеразы и область pre-S1 поверхностного белка HBV имели процент консервативных аминокислот 30% и 40% соответственно [15] Однако известно, что эти перекрывающиеся области менее важны для репликации по сравнению с перекрывающимися областями, которые были высококонсервативными среди различных штаммов HBV и абсолютно необходимы для этого процесса.
Вторая модель предполагает, что два белка и соответствующие им перекрывающиеся гены развиваются под противоположным давлением отбора: одна рамка подвергается положительному отбору , в то время как другая находится под очищающим отбором . В томбусвирусах белки p19 и p22 кодируются перекрывающимися генами, которые образуют кодирующую область длиной 549 нт, и показано, что p19 находится под положительным отбором, тогда как p22 находится под очищающим отбором. [16] Дополнительные примеры упоминаются в исследованиях с участием перекрывающихся генов вируса Сендай . [17] вирус скручивания листьев картофеля , [18] и парвовирус человека В19 . [19] Предполагается, что этот феномен перекрывающихся генов, испытывающих разное давление отбора, является следствием высокой скорости замен нуклеотидов с разными эффектами на две рамки; замены могут быть в основном несинонимичными для одного кадра, но в основном синонимичными для другого кадра. [4]
Эволюция
[ редактировать ]Перекрывающиеся гены особенно распространены в быстро развивающихся геномах, например, вирусов , бактерий и митохондрий . Они могут возникнуть тремя способами: [20]
- Путем расширения существующей открытой рамки считывания (ORF) вниз по ходу транскрипции в соседний ген из-за потери стоп-кодона ;
- Путем расширения существующей ORF вверх по ходу транскрипции в соседний ген из-за потери инициирующего кодона ;
- Путем создания новой ORF внутри существующей за счет точечной мутации .
Использование одной и той же нуклеотидной последовательности для кодирования нескольких генов может обеспечить эволюционное преимущество благодаря уменьшению размера генома и возможности транскрипции и трансляции совместной регуляции перекрывающихся генов. [12] [21] [22] [23] Перекрытие генов вводит новые эволюционные ограничения на последовательности перекрывающихся областей. [14] [24]
Происхождение новых генов
[ редактировать ]
В 1977 году Пьер-Поль Грассе предположил, что один из генов в паре мог возникнуть de novo в результате мутаций, приводящих к введению новых ORF в альтернативных рамках считывания; он описал этот механизм как надпечатку . [25] : 231 Позже это было подтверждено Сусуму Оно , который определил ген-кандидат, который мог возникнуть по этому механизму. [26] Некоторые гены de novo, возникшие таким образом, могут не оставаться перекрывающимися, а субфункциональными после дупликации генов . [6] способствуя распространению орфанных генов . Какой член перекрывающейся пары генов моложе, можно определить биоинформатически либо по более ограниченному филогенетическому распределению, либо по менее оптимизированному использованию кодонов . [9] [27] [28] Более молодые члены пары, как правило, имеют более высокую внутреннюю структурную неупорядоченность , чем старшие члены, но старшие члены также более неупорядочены, чем другие белки, предположительно в целях смягчения возросших эволюционных ограничений, вызванных перекрытием. [27] Перекрытия с большей вероятностью возникают в белках, которые уже имеют высокий уровень беспорядка. [27]
Таксономическое распространение
[ редактировать ]
Перекрывающиеся гены встречаются во всех сферах жизни , хотя и с разной частотой. Они особенно распространены в вирусных геномах.
Вирусы
[ редактировать ]
Существование перекрывающихся генов было впервые выявлено у вируса ΦX174 , чей геном был первым ДНК-геномом, секвенированным Фредериком Сэнгером в 1977 году. [29] с одноцепочечной ДНК Предыдущий анализ ΦX174, небольшого бактериофага , который инфицировал бактерию Escherichia coli , показал, что белки, вырабатываемые во время инфекции, требуют кодирующих последовательностей, длиннее, чем измеренная длина его генома. [31] Анализ полностью секвенированного генома из 5386 нуклеотидов показал, что вирус обладает значительным перекрытием между кодирующими областями, показывая, что некоторые гены (например, гены D и E) транслировались с одних и тех же последовательностей ДНК, но в разных рамках считывания. [29] [31] Было показано, что альтернативный стартовый сайт в гене репликации генома A ΦX174 экспрессирует укороченный белок с кодирующей последовательностью, идентичной С-концу исходного белка A, но обладающий другой функцией. [32] [33] Был сделан вывод, что другие неоткрытые сайты синтеза полипептидов могут быть скрыты в геноме из-за перекрывающихся генов. Было показано, что идентифицированный de novo ген другого перекрывающегося генного локуса экспрессирует новый белок, который индуцирует лизис E. coli путем ингибирования биосинтеза ее клеточной стенки [56], что позволяет предположить, что создание белка de novo в процессе оверпринтинга может иметь важное значение. фактор эволюции патогенности вирусов . [4] Другим примером является ген ORF3d в вирусе SARS-CoV 2 . [1] [34] Перекрывающиеся гены особенно распространены в вирусных геномах. [9] Некоторые исследования объясняют это наблюдение селективным давлением в сторону небольших размеров генома, опосредованным физическими ограничениями упаковки генома в вирусный капсид , особенно икосаэдрической геометрии . [35] Однако другие исследования оспаривают этот вывод и утверждают, что распределение перекрытий в вирусных геномах, скорее всего, отражает оверпринтинг как эволюционное происхождение перекрывающихся вирусных генов. [36] Оверпринтинг является распространенным источником генов de novo в вирусах. [28]
Доля вирусов с перекрывающимися кодирующими последовательностями в геномах варьируется. [2] с двухцепочечной Вирусы РНК содержат менее четверти вирусов, в то время как почти три четверти ретровирусов и вирусов с геномами одноцепочечной ДНК содержат перекрывающиеся кодирующие последовательности. [37] В частности, сегментированные вирусы или вирусы, геном которых разделен на отдельные части и упакованы либо в один и тот же капсид , либо в отдельные капсиды, с большей вероятностью будут содержать перекрывающуюся последовательность, чем несегментированные вирусы. [37] РНК-вирусы имеют меньше перекрывающихся генов, чем ДНК-вирусы, которые обладают более низкой частотой мутаций и менее ограничительными размерами генома. [37] [38] Более низкая частота мутаций ДНК-вирусов способствует большей новизне генома и эволюционным исследованиям внутри структурно ограниченного генома и может быть основной движущей силой эволюции перекрывающихся генов. [39] [40]
Исследования вирусных генов с надпечаткой показывают, что их белковые продукты, как правило, являются вспомогательными белками, которые не важны для вирусной пролиферации, но способствуют патогенности . Белки с надпечаткой часто имеют необычное распределение аминокислот и высокий уровень внутренней неупорядоченности . [41] В некоторых случаях белки с надпечаткой действительно имеют четко определенные, но новые трехмерные структуры; [42] Одним из примеров является супрессор сайленсинга РНК p19, обнаруженный у Tombusviruses , который имеет как новую структуру белка , так и новый способ связывания при распознавании siRNA . [28] [30] [43]
Прокариоты
[ редактировать ]Оценки перекрытия генов в бактериальных геномах обычно показывают, что около трети бактериальных генов перекрываются, хотя обычно только на несколько пар оснований. [12] [44] [45] Большинство исследований перекрытия в бактериальных геномах обнаруживают доказательства того, что перекрытие выполняет функцию регуляции генов , позволяя перекрывающимся генам совместно регулировать транскрипцию и трансляцию . [12] [23] В геномах прокариот наиболее распространены однонаправленные перекрытия, возможно, из-за тенденции соседних прокариотических генов иметь общую ориентацию. [12] [14] [11] Среди однонаправленных перекрытий длинные перекрытия чаще считываются со смещением в один нуклеотид в рамке считывания (т. е. фаза 1), а короткие перекрытия чаще читаются в фазе 2. [45] [46] Длинные перекрытия, составляющие более 60 пар оснований , более характерны для конвергентных генов; однако предполагаемые длинные перекрытия имеют очень высокий уровень ошибочных аннотаций . [47] Надежно подтвержденные примеры длительного перекрытия бактериальных геномов редки; в хорошо изученном модельном организме Escherichia coli только четыре пары генов хорошо подтверждены как имеющие длинные наложенные перекрытия. [48]
Эукариоты
[ редактировать ]По сравнению с геномами прокариот, геномы эукариот часто плохо аннотированы, и поэтому выявление реальных совпадений является относительно сложной задачей. [28] Однако примеры подтвержденного перекрытия генов были зарегистрированы у различных эукариотических организмов, включая млекопитающих, таких как мыши и люди. [49] [50] [51] [52] Эукариоты отличаются от прокариотов распределением типов перекрытия: хотя однонаправленные (т. е. одноцепочечные) перекрытия наиболее распространены у прокариот, у эукариот чаще встречаются перекрытия противоположных или антипараллельных цепей. Среди перекрытий противоположных цепей наиболее распространена конвергентная ориентация. [50] Большинство исследований перекрытия генов эукариот показали, что перекрывающиеся гены широко подвержены геномной реорганизации даже у близкородственных видов, и, таким образом, наличие перекрытия не всегда хорошо сохраняется. [51] [53] Перекрытие со старыми или менее таксономически ограниченными генами также является общей чертой генов, которые, вероятно, возникли de novo в данной эукариотической линии. [51] [54] [55]
Функция
[ редактировать ]Точные функции перекрывающихся генов, по-видимому, различаются в разных сферах жизни, но несколько экспериментов показали, что они важны для жизненных циклов вируса посредством правильной экспрессии белка и стехиометрии. [56] а также играет роль в правильном сворачивании белка. [57] версия бактериофага ΦX174 , в которой были удалены все перекрытия генов. Также была создана [58] доказав, что они не нужны для репликации.
Сохранение и эволюция перекрывающихся генов внутри вирусов также могут быть связаны с ограничениями размера капсида . [59] Резкая потеря жизнеспособности наблюдалась у вирусов, чьи геномы были длиннее генома дикого типа. [60] Увеличение длины генома одноцепочечной ДНК ΦX174 на >1% приводит к почти полной потере инфекционности , что, как полагают, является результатом строгих физических ограничений, налагаемых конечным объемом капсида. [61] Исследования аденоассоциированных вирусов в качестве доставки генов векторов показали, что вирусная упаковка ограничена пределами размера генетического груза, что требует использования нескольких векторов для доставки крупных человеческих генов, таких как CFTR81. [62] [63] Таким образом, предполагается, что перекрывающиеся гены возникли как средство преодоления этих физических ограничений, увеличивая генетическое разнообразие за счет использования только существующей последовательности, а не увеличения длины генома.
Методы идентификации перекрывающихся генов и ORF
[ редактировать ]Стандартизированные методы, такие как аннотация генома, могут быть неподходящими для обнаружения перекрывающихся генов, поскольку они полагаются на уже отобранные гены, в то время как перекрывающиеся гены обычно не учитываются и содержат атипичный состав последовательностей. [2] [64] [65] [66] Стандарты аннотации генома также часто предвзяты в отношении перекрытия признаков, например, когда гены полностью содержатся внутри другого гена. [67] Более того, некоторые конвейеры биоинформатики, такие как конвейер RAST, заметно наказывают за перекрытие между прогнозируемыми ORF. [68] Однако быстрое развитие инструментов измерения белков и РНК в масштабе генома, а также все более совершенных алгоритмов прогнозирования выявило лавину перекрывающихся генов и ORF внутри многочисленных геномов. [2] Протеогеномные методы сыграли важную роль в обнаружении многочисленных перекрывающихся генов и включают комбинацию таких методов, как восходящая протеомика , профилирование рибосом , секвенирование ДНК и возмущения . Секвенирование РНК также используется для идентификации областей генома, содержащих перекрывающиеся транскрипты. Он был использован для идентификации 180 000 альтернативных ORF в ранее аннотированных кодирующих областях, обнаруженных у людей. [69] Недавно обнаруженные ORF, подобные этим, проверяются с использованием различных методов обратной генетики , таких как CRISPR-Cas9 и разрушение каталитически мертвого Cas9 (dCas9) . [70] [71] [72] Также предпринимаются попытки доказательства путем синтеза, чтобы вне всякого сомнения доказать отсутствие каких-либо неоткрытых перекрывающихся генов. [73]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Нельсон, Чейз В. и др. (1 октября 2020 г.). «Динамически развивающийся новый перекрывающийся ген как фактор пандемии SARS-CoV-2» . электронная жизнь . 9 . doi : 10.7554/eLife.59633 . ПМЦ 7655111 . ПМИД 33001029 .
- ^ Jump up to: а б с д и ж Райт Б.В., Моллой, член парламента, Яшке П.Р. (5 октября 2021 г.). «Перекрывающиеся гены в природных и искусственно созданных геномах» . Обзоры природы Генетика . 23 (3): 154–168. дои : 10.1038/s41576-021-00417-w . ISSN 1471-0064 . ПМЦ 8490965 . ПМИД 34611352 .
- ^ Jump up to: а б с Ю. Фукуда, М. Томита и Т. Васио (1999). «Сравнительное изучение перекрывающихся генов в геномах Mycoplasmagentium и Mycoplasma pneumoniae » . Нуклеиновые кислоты Рез . 27 (8): 1847–1853. дои : 10.1093/нар/27.8.1847 . ПМК 148392 . ПМИД 10101192 .
- ^ Jump up to: а б с д Павези А (26 мая 2021 г.). «Происхождение, эволюция и стабильность перекрывающихся генов в вирусах: систематический обзор» . Гены . 12 (6): 809. doi : 10.3390/genes12060809 . ISSN 2073-4425 . ПМЦ 8227390 . ПМИД 34073395 .
- ^ Нормарк С., Бергстрем С., Эдлунд Т., Грундстрем Т., Яурин Б., Линдберг Ф.П., Олссон О (декабрь 1983 г.). «Перекрывающиеся гены» . Ежегодный обзор генетики . 17 (1): 499–525. дои : 10.1146/annurev.ge.17.120183.002435 . ISSN 0066-4197 . ПМИД 6198955 .
- ^ Jump up to: а б Киз П.К., Гиббс А. (15 октября 1992 г.). «Происхождение генов: «большой взрыв» или непрерывное творение?» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (20): 9489–93. Бибкод : 1992PNAS...89.9489K . дои : 10.1073/pnas.89.20.9489 . ПМК 50157 . ПМИД 1329098 .
- ^ Киз П.К., Гиббс А. (15 октября 1992 г.). «Происхождение генов: «большой взрыв» или непрерывное творение?» . Труды Национальной академии наук . 89 (20): 9489–9493. Бибкод : 1992PNAS...89.9489K . дои : 10.1073/pnas.89.20.9489 . ISSN 0027-8424 . ПМК 50157 . ПМИД 1329098 .
- ^ Гиббс А., Киз П.К. (19 октября 1995 г.), «В поисках происхождения вирусных генов» , «Молекулярные основы эволюции вирусов » , Cambridge University Press, стр. 76–90, doi : 10.1017/cbo9780511661686.008 , ISBN 978-0-521-45533-6 , получено 3 декабря 2021 г.
- ^ Jump up to: а б с д Павеси А., Магиоркинис Г., Карлин Д.Г., Уилке СО (15 августа 2013 г.). «Вирусные белки, возникшие de Novo путем надпечатки, можно идентифицировать по использованию кодонов: применение к «генному питомнику» дельтаретровирусов» . PLOS Вычислительная биология . 9 (8): e1003162. Бибкод : 2013PLSCB...9E3162P . дои : 10.1371/journal.pcbi.1003162 . ПМЦ 3744397 . ПМИД 23966842 .
- ^ Андерсон С., Банкир А.Т., Баррелл Б.Г., де Брёйн М.Х., Коулсон А.Р., Друэн Дж., Эперон И.С., Нирлих Д.П., Роу Б.А., Сэнгер Ф., Шрайер П.Х., Смит А.Дж., Стаден Р., Янг И.Г. (апрель 1981 г.). «Последовательность и организация митохондриального генома человека». Природа . 290 (5806): 457–465. Бибкод : 1981Natur.290..457A . дои : 10.1038/290457a0 . ПМИД 7219534 . S2CID 4355527 .
- ^ Jump up to: а б с Фукуда Ю., Накаяма Ю., Томита М. (декабрь 2003 г.). «О динамике перекрывающихся генов в бактериальных геномах». Джин . 323 : 181–187. дои : 10.1016/j.gene.2003.09.021 . ПМИД 14659892 .
- ^ Jump up to: а б с д и ж Джонсон З., Чисхолм С. (2004). «Свойства перекрывающихся генов сохраняются во всех микробных геномах» . Геном Рез . 14 (11): 2268–72. дои : 10.1101/гр.2433104 . ПМК 525685 . ПМИД 15520290 .
- ^ Jump up to: а б Нормарк С., Бергстром С., Эдлунд Т., Грундстрем Т., Яурин Б., Линдберг Ф.П., Олссон О. (1983). «Перекрывающиеся гены». Ежегодный обзор генетики . 17 : 499–525. дои : 10.1146/annurev.ge.17.120183.002435 . ПМИД 6198955 .
- ^ Jump up to: а б с д Рогозин И.Б., Спиридонов А.Н., Сорокин А.В., Вольф Ю.И., Йордан И., Татусов Р.Л., Кунин Е.В. (май 2002 г.). «Очистка и направленный отбор перекрывающихся прокариотических генов». Тенденции в генетике . 18 (5): 228–232. дои : 10.1016/S0168-9525(02)02649-5 . ПМИД 12047938 .
- ^ Jump up to: а б Мизоками М., Орито Э., Оба Ки, Икео К., Лау Дж. Я., Годобори Т. (январь 1997 г.). «Ограниченная эволюция в отношении перекрытия генов вируса гепатита B» . Журнал молекулярной эволюции . 44 (С1): С83–С90. Бибкод : 1997JMolE..44S..83M . дои : 10.1007/pl00000061 . ISSN 0022-2844 . ПМИД 9071016 . S2CID 22644652 .
- ^ Эллисон-младший, Лехнер М., Хоеппнер, член парламента, Пул А.М. (12 февраля 2016 г.). «Положительный отбор или свобода варьирования? Оценка функционального значения изменения последовательности с помощью молекулярной динамики» . ПЛОС ОДИН . 11 (2): e0147619. Бибкод : 2016PLoSO..1147619A . дои : 10.1371/journal.pone.0147619 . ISSN 1932-6203 . ПМЦ 4752228 . ПМИД 26871901 .
- ^ Фуджи И., Киётани К., Ёсида Т., Сакагути Т. (2001). «Консервативные и неконсервативные области генома вируса Сендай: эволюция гена, обладающего перекрывающимися рамками считывания» . Гены вирусов . 22 (1): 47–52. дои : 10.1023/а:1008130318633 . ISSN 0920-8569 . ПМИД 11210938 . S2CID 12869504 .
- ^ Гаядер С., Дюкре Д.Г. (1 июля 2002 г.). «Анализ последовательностей изолятов вируса скручивания листьев картофеля выявляет генетическую стабильность, основные эволюционные события и дифференциальное давление отбора между перекрывающимися продуктами рамки считывания» . Журнал общей вирусологии . 83 (7): 1799–1807. дои : 10.1099/0022-1317-83-7-1799 . ISSN 0022-1317 . ПМИД 12075102 .
- ^ Стаменкович Г.Г., Чиркович В.С., Шилич М.М., Благоевич Й.В., Кнежевич А.М., Йоксич И.Д., Станоевич М.П. (24 октября 2016 г.). «Степень замещения и естественный отбор в парвовирусе B19» . Научные отчеты . 6 (1): 35759. Бибкод : 2016NatSR...635759S . дои : 10.1038/srep35759 . ISSN 2045-2322 . ПМК 5075947 . ПМИД 27775080 .
- ^ Кракауэр, округ Колумбия (июнь 2000 г.). «Стабильность и эволюция перекрывающихся генов» . Эволюция . 54 (3): 731–739. дои : 10.1111/j.0014-3820.2000.tb00075.x . ПМИД 10937248 . S2CID 8818055 .
- ^ Делэй Л., ДеЛуна А., Ласкано А., Бесерра А. (2008). «Происхождение нового гена посредством надпечатки в Escherichia coli» . Эволюционная биология BMC . 8 (1): 31. Бибкод : 2008BMCEE...8...31D . дои : 10.1186/1471-2148-8-31 . ПМК 2268670 . ПМИД 18226237 .
- ^ Саха Д., Поддер С., Панда А., Гош Т.С. (май 2016 г.). «Перекрывающиеся гены: значительный геномный коррелят темпов роста прокариот». Джин . 582 (2): 143–147. дои : 10.1016/j.gene.2016.02.002 . ПМИД 26853049 .
- ^ Jump up to: а б Луо Й, Баттистуцци Ф, Лин К, Гибас С (29 ноября 2013 г.). «Эволюционная динамика перекрывающихся генов сальмонеллы» . ПЛОС ОДИН . 8 (11): е81016. Бибкод : 2013PLoSO...881016L . дои : 10.1371/journal.pone.0081016 . ПМЦ 3843671 . ПМИД 24312259 .
- ^ Вэй Икс, Чжан Дж (31 декабря 2014 г.). «Простой метод оценки силы естественного отбора по перекрывающимся генам» . Геномная биология и эволюция . 7 (1): 381–390. дои : 10.1093/gbe/evu294 . ПМЦ 4316641 . ПМИД 25552532 .
- ^ Грассе ПП (1977). Эволюция живых организмов: доказательства новой теории трансформации . Академическая пресса. ISBN 978-1-4832-7409-6 .
- ^ Оно С. (апрель 1984 г.). «Рождение уникального фермента из альтернативной рамки считывания ранее существовавшей, внутренне повторяющейся кодирующей последовательности» . Труды Национальной академии наук Соединенных Штатов Америки . 81 (8): 2421–5. Бибкод : 1984PNAS...81.2421O . дои : 10.1073/pnas.81.8.2421 . ПМК 345072 . ПМИД 6585807 .
- ^ Jump up to: а б с Уиллис С., Мэйсел Дж. (19 июля 2018 г.). «Рождение генов способствует структурному расстройству, кодируемому перекрывающимися генами» . Генетика . 210 (1): 303–313. дои : 10.1534/genetics.118.301249 . ПМК 6116962 . ПМИД 30026186 .
- ^ Jump up to: а б с д Сабат Н., Вагнер А., Карлин Д. (19 июля 2012 г.). «Эволюция вирусных белков, возникших de Novo путем надпечатки» . Молекулярная биология и эволюция . 29 (12): 3767–3780. дои : 10.1093/molbev/mss179 . ПМЦ 3494269 . ПМИД 22821011 .
- ^ Jump up to: а б с Сэнгер Ф., генеральный директор Air, Баррелл Б.Г., Браун Н.Л., Коулсон А.Р., Фиддес Дж.К., Хатчисон К.А., Слокомб П.М., Смит М. (1977). «Нуклеотидная последовательность ДНК бактериофага ΦX174». Природа . 265 (5596): 687–95. Бибкод : 1977Natur.265..687S . дои : 10.1038/265687a0 . ПМИД 870828 . S2CID 4206886 .
- ^ Jump up to: а б Е.К., Малинина Л., Патель DJ (3 декабря 2003 г.). «Распознавание малых интерферирующих РНК вирусным супрессором молчания РНК» . Природа . 426 (6968): 874–878. Бибкод : 2003Natur.426..874Y . дои : 10.1038/nature02213 . ПМЦ 4694583 . ПМИД 14661029 .
- ^ Jump up to: а б Баррелл Б.Г., генеральный директор Air, Хатчисон, Калифорния (ноябрь 1976 г.). «Перекрывающиеся гены в бактериофаге φX174» . Природа . 264 (5581): 34–41. Бибкод : 1976Natur.264...34B . дои : 10.1038/264034a0 . ISSN 1476-4687 . ПМИД 1004533 . S2CID 4264796 .
- ^ ЛИННИ Э., ХАЯШИ М. (май 1974 г.). «Внутригенная регуляция синтеза белков гена А ΦX174» . Природа . 249 (5455): 345–348. Бибкод : 1974Natur.249..345L . дои : 10.1038/249345a0 . ISSN 0028-0836 . ПМИД 4601823 . S2CID 4175651 .
- ^ Рожновский А.П., Дор С.М., Кемп С.З., Фейн Б.А. (6 января 2020 г.). «Наконец, роль, достойная звезды: сильно консервативные, несущественные белки микровируса А* обеспечивают точность реакций упаковки продукта» . Журнал вирусологии . 94 (2). дои : 10.1128/jvi.01593-19 . ISSN 0022-538X . ПМК 6955274 . PMID 31666371 .
- ^ Докрилл П (11 ноября 2020 г.). «Ученые только что обнаружили загадочно скрытый «ген внутри гена» SARS-CoV-2» . НаукаАлерт . Проверено 11 ноября 2020 г.
- ^ Кирико Н., Вианелли А., Белшоу Р. (7 июля 2010 г.). «Почему гены у вирусов перекрываются» . Труды Королевского общества B: Биологические науки . 277 (1701): 3809–3817. дои : 10.1098/rspb.2010.1052 . ПМЦ 2992710 . ПМИД 20610432 .
- ^ Брандес Н., Линиал М. (21 мая 2016 г.). «Перекрытие генов и ограничения размера в вирусном мире» . Биология Директ . 11 (1): 26. дои : 10.1186/s13062-016-0128-3 . ПМЦ 4875738 . ПМИД 27209091 .
- ^ Jump up to: а б с Schlub TE, Holmes EC (1 января 2020 г.). «Свойства и количество перекрывающихся генов у вирусов» . Эволюция вирусов . 6 (1): veaa009. дои : 10.1093/ve/veaa009 . ISSN 2057-1577 . ПМК 7017920 . ПМИД 32071766 .
- ^ Кирико Н., Вианелли А., Белшоу Р. (7 июля 2010 г.). «Почему гены у вирусов перекрываются» . Труды Королевского общества B: Биологические науки . 277 (1701): 3809–3817. дои : 10.1098/rspb.2010.1052 . ISSN 0962-8452 . ПМЦ 2992710 . ПМИД 20610432 .
- ^ Брандес Н., Линиал М. (21 мая 2016 г.). «Перекрытие генов и ограничения размера в вирусном мире» . Биология Директ . 11 (1): 26. дои : 10.1186/s13062-016-0128-3 . ISSN 1745-6150 . ПМЦ 4875738 . ПМИД 27209091 .
- ^ Павеси А (июль 2020 г.). «Новое понимание эволюционных особенностей вирусных перекрывающихся генов с помощью дискриминантного анализа» . Вирусология . 546 : 51–66. дои : 10.1016/j.virol.2020.03.007 . ISSN 0042-6822 . ПМЦ 7157939 . ПМИД 32452417 .
- ^ Ранкурел С., Хосрави М., Данкер А.К., Ромеро П.Р., Карлин Д. (29 июля 2009 г.). «Перекрывающиеся гены производят белки с необычными свойствами последовательности и дают представление о создании белков De Novo» . Журнал вирусологии . 83 (20): 10719–10736. дои : 10.1128/JVI.00595-09 . ПМК 2753099 . ПМИД 19640978 .
- ^ Аброй А (1 декабря 2015 г.). «Взгляд на отношения виросфера-хозяин, основанный на белковых доменах». Биохимия . 119 : 231–243. дои : 10.1016/j.biochi.2015.08.008 . ПМИД 26296474 .
- ^ Варгасон Дж. М., Ситтия Г., Бургян Дж., Холл Т. М. (декабрь 2003 г.). «Избирательное по размеру распознавание миРНК супрессором молчания РНК» . Клетка . 115 (7): 799–811. дои : 10.1016/S0092-8674(03)00984-X . ПМИД 14697199 . S2CID 12993441 .
- ^ Хувет М., депутат Штумпфа (1 января 2014 г.). «Перекрывающиеся гены: окно в эволюционность генов» . БМК Геномика . 15 (1): 721. дои : 10.1186/1471-2164-15-721 . ISSN 1471-2164 . ПМК 4161906 . ПМИД 25159814 .
- ^ Jump up to: а б Кок Пи Джей, Уитворт Д.Э. (19 марта 2007 г.). «Эволюция перекрытий генов: относительное смещение рамки считывания в генах прокариотической двухкомпонентной системы». Журнал молекулярной эволюции . 64 (4): 457–462. Бибкод : 2007JMolE..64..457C . дои : 10.1007/s00239-006-0180-1 . ПМИД 17479344 . S2CID 21612308 .
- ^ Фонсека М.М., Харрис DJ, Посада Д. (5 ноября 2013 г.). «Происхождение и распределение длины однонаправленных перекрывающихся генов прокариот» . G3: Гены, геномы, генетика . 4 (1): 19–27. дои : 10.1534/g3.113.005652 . ПМЦ 3887535 . ПМИД 24192837 .
- ^ Пальеха А., Харрингтон Э.Д., Борк П. (2008). «Большие перекрытия генов в геномах прокариот: результат функциональных ограничений или неверных прогнозов?» . БМК Геномика . 9 (1): 335. дои : 10.1186/1471-2164-9-335 . ПМЦ 2478687 . ПМИД 18627618 .
- ^ Феллнер Л., Саймон С., Шерлинг С., Виттинг М., Шобер С., Полте С., Шмитт-Копплин П., Кейм Д.А., Шерер С., Нойхаус К. (18 декабря 2015 г.). «Доказательства недавнего происхождения кодирующего бактериальный белок перекрывающегося гена-сироты в результате эволюционной надпечатки» . Эволюционная биология BMC . 15 (1): 283. Бибкод : 2015BMCEE..15..283F . дои : 10.1186/s12862-015-0558-z . ПМЦ 4683798 . ПМИД 26677845 .
- ^ МакЛисахт А., Герцони Д. (31 августа 2015 г.). «Новые гены из некодирующей последовательности: роль генов, кодирующих белки de novo, в эволюционных инновациях эукариот» . Философские труды Королевского общества B: Биологические науки . 370 (1678): 20140332. doi : 10.1098/rstb.2014.0332 . ПМЦ 4571571 . ПМИД 26323763 .
- ^ Jump up to: а б К. Санна, В. Ли и Л. Чжан (2008). «Перекрывающиеся гены в геномах человека и мыши» . БМК Геномика . 9 (169): 169. дои : 10.1186/1471-2164-9-169 . ПМК 2335118 . ПМИД 18410680 .
- ^ Jump up to: а б с Макаловска И., Лин К.Ф., Эрнандес К. (2007). «Рождение и смерть перекрывающихся генов у позвоночных» . Эволюционная биология BMC . 7 (1): 193. Бибкод : 2007BMCEE...7..193M . дои : 10.1186/1471-2148-7-193 . ПМК 2151771 . ПМИД 17939861 .
- ^ Вирамачанени V (1 февраля 2004 г.). «Перекрывающиеся гены млекопитающих: сравнительная перспектива» . Геномные исследования . 14 (2): 280–286. дои : 10.1101/гр.1590904 . ПМК 327103 . ПМИД 14762064 .
- ^ Бехура С.К., Северсон Д.В. (2013). «Перекрывающиеся гены Aedes aegypti: эволюционные последствия сравнения с ортологами Anopheles gambiae и другими насекомыми» . Эволюционная биология BMC . 13 (1): 124. Бибкод : 2013BMCEE..13..124B . дои : 10.1186/1471-2148-13-124 . ПМЦ 3689595 . ПМИД 23777277 .
- ^ Мерфи Д.Н., МакЛисахт А., Кармел Л. (21 ноября 2012 г.). «De Novo Происхождение генов, кодирующих белки, у мышевидных грызунов» . ПЛОС ОДИН . 7 (11): е48650. Бибкод : 2012PLoSO...748650M . дои : 10.1371/journal.pone.0048650 . ПМК 3504067 . ПМИД 23185269 .
- ^ Ноулз Д.Г., Маклизат А. (2 сентября 2009 г.). «Недавнее происхождение генов, кодирующих человеческие белки», de novo . Геномные исследования . 19 (10): 1752–1759. дои : 10.1101/гр.095026.109 . ПМЦ 2765279 . ПМИД 19726446 .
- ^ Райт Б.В., Руан Дж., Моллой, член парламента, Яшке П.Р. (20 ноября 2020 г.). «Модуляризация генома показывает, что перекрывающаяся топология генов необходима для эффективного воспроизводства вирусов» . ACS Синтетическая биология . 9 (11): 3079–3090. doi : 10.1021/acsynbio.0c00323 . ISSN 2161-5063 . ПМИД 33044064 . S2CID 222300240 .
- ^ Прадхан П., Ли В., Каур П. (январь 2009 г.). «Трансляционное соединение контролирует экспрессию и функцию откачивающего насоса лекарства DrrAB» . Журнал молекулярной биологии . 385 (3): 831–842. дои : 10.1016/j.jmb.2008.11.027 . ПМИД 19063901 .
- ^ Яшке П.Р., Либерман Э.К., Родригес Дж., Сьерра А., Энди Д. (декабрь 2012 г.). «Полностью декомпрессированный синтетический геном бактериофага øX174, собранный и заархивированный в дрожжах» . Вирусология . 434 (2): 278–284. дои : 10.1016/j.virol.2012.09.020 . ISSN 0042-6822 . ПМИД 23079106 .
- ^ Кракауэр Д.С., Плоткин Дж.Б. (29 января 2002 г.). «Избыточность, антиизбыточность и надежность геномов» . Труды Национальной академии наук . 99 (3): 1405–1409. Бибкод : 2002PNAS...99.1405K . дои : 10.1073/pnas.032668599 . ISSN 0027-8424 . ПМК 122203 . ПМИД 11818563 .
- ^ Фейсс М., Фишер Р., Крейтон М., Эгнер С. (март 1977 г.). «Упаковка хромосомы бактериофага λ: влияние длины хромосомы» . Вирусология . 77 (1): 281–293. дои : 10.1016/0042-6822(77)90425-1 . ISSN 0042-6822 . ПМИД 841861 .
- ^ Аояма А., Хаяси М. (сентябрь 1985 г.). «Влияние размера генома на упаковку ДНК бактериофага phi X174 in vitro» . Журнал биологической химии . 260 (20): 11033–11038. дои : 10.1016/s0021-9258(17)39144-5 . ISSN 0021-9258 . ПМИД 3161888 . S2CID 32443408 .
- ^ Ву Цзы, Ян Х., Колози П. (январь 2010 г.). «Влияние размера генома на упаковку вектора AAV» . Молекулярная терапия . 18 (1): 80–86. дои : 10.1038/mt.2009.255 . ISSN 1525-0016 . ПМК 2839202 . ПМИД 19904234 .
- ^ Вайдьянатан С., Байк Р., Чен Л., Браво Д.Т., Суарес С.Дж., Абазари С.М., Салахудин А.А., Дудек А.М., Теран К.А., Дэвис Т.Х., Ли СМ (март 2021 г.). «Целевая замена полноразмерного CFTR в стволовых клетках дыхательных путей человека на CRISPR-Cas9 для коррекции панмутации в эндогенном локусе» . Молекулярная терапия . 30 (1): 223–237. дои : 10.1016/j.ymthe.2021.03.023 . ПМЦ 8753290 . ПМИД 33794364 . S2CID 232761334 .
- ^ Уиллис С., Мэйсел Дж. (19 июля 2018 г.). «Рождение генов способствует структурному расстройству, кодируемому перекрывающимися генами» . Генетика . 210 (1): 303–313. дои : 10.1534/genetics.118.301249 . ISSN 1943-2631 . ПМК 6116962 . ПМИД 30026186 .
- ^ Павеси А., Вианелли А., Кирико Н., Бао Ю., Блинкова О., Белшоу Р., Ферт А., Карлин Д. (19 октября 2018 г.). «Перекрывающиеся гены и белки, которые они кодируют, значительно отличаются по составу последовательностей от неперекрывающихся генов» . ПЛОС ОДИН . 13 (10): e0202513. Бибкод : 2018PLoSO..1302513P . дои : 10.1371/journal.pone.0202513 . ISSN 1932-6203 . ПМК 6195259 . ПМИД 30339683 .
- ^ Павеси А., Магиоркинис Г., Карлин Д.Г. (15 августа 2013 г.). «Вирусные белки, возникшие de Novo путем надпечатки, можно идентифицировать по использованию кодонов: применение к «генному питомнику» дельтаретровирусов» . PLOS Вычислительная биология . 9 (8): e1003162. Бибкод : 2013PLSCB...9E3162P . дои : 10.1371/journal.pcbi.1003162 . ISSN 1553-7358 . ПМЦ 3744397 . ПМИД 23966842 .
- ^ «Дополнительная информация 2: Информация о доступе к базе данных генома NCBI (файл PDF)» . дои : 10.7717/peerj.6447/supp-2 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Ахмед Н. (27 марта 2009 г.). «Рекомендации профессорско-преподавательского состава по серверу RAST: быстрые аннотации с использованием технологии подсистем» . дои : 10.3410/f.1157743.618965 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Бен-Тал Н, изд. (23 июня 2017 г.). «Письмо о решении: Глубокая аннотация транскриптома позволяет обнаруживать и функциональную характеристику загадочных малых белков» . дои : 10.7554/elife.27860.082 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Баззини А., Ву Ц (6 марта 2020 г.). «Рекомендации мнений преподавателей о всеобъемлющем функциональном переводе неканонических человеческих открытых рамок чтения» . дои : 10.3410/f.737484924.793572056 . S2CID 215850701 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Пренснер Дж.Р., Эначе О.М., Лурия В., Круг К., Клаузер К.Р., Демпстер Дж.М., Каргер А., Ван Л., Стумбрайт К., Ван В.М., Ботта Г. (28 января 2021 г.). «Неканонические открытые рамки считывания кодируют функциональные белки, необходимые для выживания раковых клеток» . Природная биотехнология . 39 (6): 697–704. дои : 10.1038/s41587-020-00806-2 . ISSN 1087-0156 . ПМЦ 8195866 . ПМИД 33510483 .
- ^ Цао X, Хитун А, Луо Й, На З, Пхудокмай Т, Саппахав К, Олатунджи Е, Уттамапинант С, Славофф С.А. (5 марта 2020 г.). «Alt-RPL36 подавляет сигнальный путь PI3K-AKT-mTOR путем взаимодействия с TMEM24» . Природные коммуникации . 12 (1): 508. bioRxiv 10.1101/2020.03.04.977314 . дои : 10.1038/s41467-020-20841-6 . ПМК 7820019 . ПМИД 33479206 .
- ^ Яшке П.Р., Дотсон Г.А., Хунг К.С., Лю Д., Энди Д. (12 ноября 2019 г.). «Окончательная демонстрация путем синтеза полноты аннотации генома» . Труды Национальной академии наук . 116 (48): 24206–24213. Бибкод : 2019PNAS..11624206J . дои : 10.1073/pnas.1905990116 . ISSN 0027-8424 . ПМК 6883844 . ПМИД 31719208 .