РНК-вирус
Части этой статьи (относящиеся к таксономии в разделах Балтимора [выпуск ICTV 2018b → 2019]) необходимо обновить . ( январь 2021 г. ) |

РНК -вирус — это вирус (кроме ретровирусов которого содержит рибонуклеиновую кислоту ( РНК ) ), генетический материал . [1] Нуклеиновая кислота обычно представляет собой одноцепочечную РНК ( оцРНК ), но может быть и двухцепочечной (дцРНК). [2] Известные заболевания человека, вызываемые РНК-вирусами, включают простуду , грипп , атипичную пневмонию , MERS , COVID-19 , вирус Денге , гепатит С , гепатит Е , лихорадку Западного Нила , болезнь, вызванную вирусом Эбола , бешенство , полиомиелит , эпидемический паротит и корь .
Международный комитет по таксономии вирусов (ICTV) классифицирует РНК-вирусы как те, которые принадлежат к группе III , группе IV или группе V балтиморской классификационной системы. Эта категория исключает группу VI , вирусы с генетическим материалом РНК, но использующие промежуточные ДНК в своем жизненном цикле : они называются ретровирусами . [3] включая ВИЧ-1 и ВИЧ-2, которые вызывают СПИД .
По состоянию на май 2020 года считается, что все известные РНК-вирусы, кодирующие РНК-направленную РНК-полимеразу, образуют монофилетическую группу, известную как царство рибовирий . [4] Большинство таких РНК-вирусов относятся к царству Orthornavirae , а положение остальных еще не определено . [5] Область содержит не все РНК-вирусы: Deltavirus , Asunviroidae и Pospiviroidae — таксоны РНК-вирусов, ошибочно включенные в 2019 г. [а] но исправлено в 2020 году. [6]
Характеристики [ править ]
Вирусы с одноцепочечной РНК смысл РНК и
РНК-вирусы можно дополнительно классифицировать в зависимости от смысла или полярности их РНК на РНК-вирусы с отрицательным и положительным смыслом или амбисенсные РНК-вирусы. Вирусная РНК с положительным смыслом аналогична мРНК и, следовательно, может быть немедленно транслирована клеткой-хозяином. Вирусная РНК с отрицательным смыслом комплементарна мРНК и, следовательно, перед трансляцией должна быть преобразована в РНК с положительным смыслом с помощью РНК-зависимой РНК-полимеразы . Очищенная РНК вируса с положительным смыслом может непосредственно вызывать инфекцию, хотя она может быть менее заразной, чем целая вирусная частица. Напротив, очищенная РНК вируса с отрицательным смыслом сама по себе не является инфекционной, поскольку ее необходимо транскрибировать в РНК с положительным смыслом; каждый вирион может быть транскрибирован несколькими РНК с положительным смыслом. Вирусы с амбисенсовой РНК напоминают вирусы с РНК с отрицательным смыслом, за исключением того, что они транслируют гены со своих отрицательных и положительных цепей. [7]
Вирусы с двухцепочечной РНК [ править ]
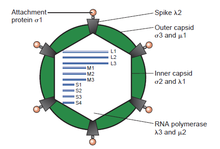
Вирусы с двухцепочечной (дц)РНК представляют собой разнообразную группу вирусов, которые широко различаются в зависимости от круга хозяев (человек, животные, растения, грибы , [б] и бактерий ), количество сегментов генома (от одного до двенадцати) и вириона организацию ( число триангуляции , слои капсида , шипы, турели и т. д.). К представителям этой группы относятся ротавирусы , которые являются наиболее распространенной причиной гастроэнтерита у детей раннего возраста, и пикобирнавирусы , которые являются наиболее частыми вирусами в образцах фекалий людей и животных с признаками диареи или без них. Вирус блютанга является экономически важным возбудителем, поражающим крупный рогатый скот и овец. В последние годы был достигнут прогресс в определении структур с атомным и субнанометровым разрешением ряда ключевых вирусных белков и капсидов вирионов нескольких дцРНК-вирусов, что подчеркивает существенные параллели в структуре и процессах репликации многих из этих вирусов. [2] [ нужна страница ]
Скорость мутаций [ править ]
РНК-вирусы обычно имеют очень высокий уровень мутаций по сравнению с ДНК-вирусами . [9] потому что вирусные РНК-полимеразы не обладают корректировать способностью ДНК-полимеразы . [10] Генетическое разнообразие РНК-вирусов является одной из причин, почему против них трудно создать эффективные вакцины . [11] Ретровирусы также имеют высокую частоту мутаций, даже несмотря на то, что их промежуточная ДНК интегрируется в геном хозяина (и, таким образом, после интеграции подлежит проверке ДНК хозяина), поскольку ошибки во время обратной транскрипции встраиваются в обе цепи ДНК перед интеграцией. [12] Некоторые гены РНК-вируса важны для циклов репликации вируса, и мутации недопустимы. Например, область генома вируса гепатита С , кодирующая основной белок, высококонсервативна . [13] потому что он содержит структуру РНК, вовлеченную во внутренний сайт входа в рибосому . [14]
Сложность последовательности [ править ]
В среднем вирусы дцРНК демонстрируют меньшую избыточность последовательности по сравнению с вирусами оцРНК. Напротив, вирусы дцДНК содержат больше всего избыточных последовательностей генома, а вирусы оцДНК — меньше всего. [15] Было показано, что сложность последовательности вирусов является ключевой характеристикой для точной безреферентной вирусной классификации. [15]
Репликация [ править ]
РНК-вирусы животных классифицируются ICTV. Существует три отдельные группы РНК-вирусов в зависимости от их генома и способа репликации:
- Вирусы с двухцепочечной РНК (группа III) содержат от одной до дюжины различных молекул РНК, каждая из которых кодирует один или несколько вирусных белков.
- Геном вирусов оцРНК с положительным смыслом (группа IV) напрямую используется в виде мРНК, при этом рибосомы хозяина переводят его в один белок, который модифицируется белками хозяина и вируса с образованием различных белков, необходимых для репликации. Одна из них включает РНК-зависимую РНК-полимеразу (РНК-репликазу), которая копирует вирусную РНК с образованием двухцепочечной репликативной формы. В свою очередь, эта дцРНК направляет образование новой вирусной РНК.
- Геном вирусов оцРНК с отрицательным смыслом (группа V) должен быть скопирован с помощью РНК-репликазы для образования РНК с положительным смыслом. Это означает, что вирус должен принести с собой фермент РНК-реплицазу. Молекула РНК с положительным смыслом затем действует как вирусная мРНК, которая транслируется в белки рибосомами хозяина.
Ретровирусы (группа VI) имеют геном с одноцепочечной РНК, но, как правило, не считаются РНК-вирусами, поскольку для репликации они используют промежуточные ДНК. Обратная транскриптаза , вирусный фермент, который выделяется из самого вируса после того, как он лишен оболочки, преобразует вирусную РНК в комплементарную цепь ДНК, которая копируется с образованием двухцепочечной молекулы вирусной ДНК. После того, как эта ДНК интегрируется в геном хозяина с помощью вирусного фермента интегразы , экспрессия закодированных генов может привести к образованию новых вирионов.
Рекомбинация [ править ]
Многочисленные РНК-вирусы способны к генетической рекомбинации по крайней мере два вирусных генома . , когда в одной и той же клетке-хозяине присутствуют [16] Очень редко вирусная РНК может рекомбинировать с РНК хозяина. [17] Рекомбинация РНК, по-видимому, является основной движущей силой в определении архитектуры генома и хода вирусной эволюции среди пикорнавирусов ( (+)ssRNA ), например, полиовируса . [18] У Retroviridae ((+)ssRNA), например ВИЧ , повреждения генома РНК, по-видимому, можно избежать во время обратной транскрипции за счет переключения цепи, формы рекомбинации. [19] [20] [21] Рекомбинация также происходит у Reoviridae (дсРНК), например, у реовирусов; Orthomyxoviridae ((-)ssRNA), например, вирус гриппа ; [21] и Coronaviridae ((+)ssRNA), например SARS . [22] Рекомбинация РНК-вирусов, по-видимому, является адаптацией, позволяющей справиться с повреждением генома. [16] Рекомбинация может происходить нечасто между вирусами животных одного и того же вида, но разных линий. Полученные рекомбинантные вирусы иногда могут вызывать вспышки инфекции у людей. [22]
Классификация [ править ]
Этот раздел может потребовать очистки Википедии , чтобы соответствовать стандартам качества . Конкретная проблема заключается в следующем: устаревшая и излишняя статья о рибовирии , а также более поздний текст; см. ДНК-вирус для чистой интеграции между ICTV высшего порядка и Балтимором. ( январь 2021 г. ) |
Классификация основана главным образом на типе генома (двухцепочечный, отрицательный или положительный одноцепочечный), а также количестве и организации генов. В настоящее время известно 5 отрядов и 47 семейств РНК-вирусов. Есть также много неустановленных видов и родов.
Родственными, но отличными от РНК-вирусов, являются вироиды и РНК-сателлитные вирусы . В настоящее время они не классифицируются как РНК-вирусы и описаны на отдельных страницах.
Исследование нескольких тысяч РНК-вирусов показало наличие как минимум пяти основных таксонов: группы левивирусов и родственников; супергруппа пикорнавирусов; супергруппа альфавирусов плюс супергруппа флавивирусов; вирусы дсРНК; и -пятицепочечные вирусы. [23] Группа лентивирусов , по-видимому, является базовой для всех остальных РНК-вирусов. Следующее крупное подразделение лежит между пикорнасупрагруппой и остальными вирусами. Вирусы с дцРНК, по-видимому, произошли от предка с +ve РНК, а вирусы с -ve РНК произошли от вирусов с дцРНК. Ближайшим родственником вирусов с пятицепочечной РНК является Reoviridae .
РНК-вирусы цепью положительной с
Это самая большая группа РНК-вирусов. [24] и был организован ICTV в типы Kitrinoviricota , Lenarviricota и Pisuviricota в царстве Orthornavirae и царстве Riboviria . [25]
РНК-вирусы с положительной цепью также можно классифицировать на основе РНК-зависимой РНК-полимеразы. Были признаны три группы: [26]
- Бимовирусы, комовирусы, неповирусы, нодавирусы, пикорнавирусы, потивирусы, собемовирусы и подмножество лютеовирусов (вирус западной желтизны свеклы и вирус скручивания листьев картофеля) — пикорноподобная группа (Picornavirata).
- Кармовирусы, диантовирусы, флавивирусы, пестивирусы, статовирусы, томбусвирусы, бактериофаги с одноцепочечной РНК, вирус гепатита С и субпопуляция лютеовирусов (вирус желтой карликовости ячменя) — флавиподобная группа (Flavivirata).
- Альфавирусы, карлавирусы, фуровирусы, хордеивирусы, потексвирусы, рубивирусы, тобравирусы, трикорнавирусы, тимовирусы, вирус хлоротичной пятнистости листьев яблони, вирус желтухи свеклы и вирус гепатита Е — альфа-подобная группа (Rubivirata).
Предложено разделение альфа-подобной (синдбис-подобной) супергруппы на основе нового домена, расположенного вблизи N-концев белков, участвующих в репликации вируса. [27] Предлагаются две группы: группа «альтовирусов» (альфавирусы, фуровирусы, вирус гепатита Е, хордеивирусы, тобамовирусы, тобравирусы, трикорнавирусы и, возможно, рубивирусы); и группа «типовирусов» (вирус хлоротичной пятнистости листьев яблони, карлавирусы, потексвирусы и тимовирусы).
Альфа-подобную супергруппу можно разделить на три клады : рубиподобные, тобамоподобные и тимоподобные вирусы. [28]
Дополнительная работа выявила пять групп РНК-вирусов с положительной цепью, содержащих четыре, три, три, три и один порядок (порядки) соответственно. [29] Эти четырнадцать порядков содержат 31 семейство вирусов (в том числе 17 семейств вирусов растений) и 48 родов (в том числе 30 родов вирусов растений). Этот анализ предполагает, что альфавирусы и флавивирусы можно разделить на два семейства — Togaviridae и Flaviridae соответственно, но предполагает, что другие таксономические отнесения, такие как пестивирусы, вирус гепатита С, рубивирусы, вирус гепатита Е и артеривирусы, могут быть неверными. Коронавирусы и торовирусы представляют собой отдельные семейства, относящиеся к разным порядкам, а не отдельные роды одного и того же семейства, как это классифицируется в настоящее время. Лютеовирусы относятся к двум семействам, а не к одному, а вирус хлоротичной пятнистости листьев яблони, по-видимому, не является клостеровирусом, а представляет собой новый род Potexviridae.
Эволюция [ править ]
Эволюция пикорнавирусов, основанная на анализе их РНК-полимераз и геликаз , по-видимому, восходит к дивергенции эукариот . [30] Их предполагаемые предки включают бактериальные ретроэлементы группы II , семейство протеаз HtrA и ДНК- бактериофаги .
Партитивирусы связаны с предком тотивируса и, возможно, произошли от него. [31]
Гиповирусы и барнавирусы, по-видимому, имеют общее происхождение с линиями потивирусов и собемовирусов соответственно. [31]
Вирусы с двухцепочечной РНК [ править ]
Этот анализ также предполагает, что вирусы дцРНК не являются тесно связанными друг с другом, а вместо этого принадлежат к четырем дополнительным классам — Birnaviridae, Cystoviridae, Partitiviridae и Reoviridae — и одному дополнительному порядку (Totiviridae) одного из классов положительных оцРНК-вирусов в того же подтипа, что и РНК-вирусы с положительной цепью.
Одно исследование показало, что существует две большие клады: одна включает семейства Caliciviridae , Flaviviridae и Picornaviridae , а вторая включает семейства Alphatetraviridae , Birnaviridae , Cystoviridae , Nodaviridae и Permutotretraviridae . [32]
цепью РНК отрицательной с Вирусы
Эти вирусы имеют несколько типов генома: от одной молекулы РНК до восьми сегментов. Несмотря на их разнообразие, похоже, что они произошли от членистоногих и оттуда развились. [33]
Спутниковые вирусы [ править ]
Также известен ряд вирусов-сателлитов — вирусов, которым для завершения жизненного цикла требуется помощь другого вируса. Их таксономия еще не определена. Следующие четыре рода были предложены для сателлитных вирусов с одноцепочечной РНК с положительным смыслом, которые заражают растения: Albetovirus , Aumaivirus , Papanivirus и Virtovirus . [34] Семейство Sarthroviridae , включающее род Macronovirus , было предложено для сателлитных вирусов с одноцепочечной РНК с положительным смыслом, которые заражают членистоногих .
Группа III дцРНК – вирусы
В эту группу входят двенадцать семейств и ряд неназначенных родов и видов. [10]
- Семейство Амальгавирусиды.
- Семейство Birnaviridae
- Семейство Chrysoviridae
- Семейство Cystoviridae
- Семейство Эндорнавирусиды
- Семейство гиповирусов
- Семейство Мегабирнавирусиды.
- Семейство Partitiviridae
- Семейство Picobirnaviridae.
- Семейство Reoviridae - включает ротавирус.
- Семейство Тотивирусиды
- Семейство Quadriviridae
- Род Ботибирнавирус.
- Неназначенные виды
IV – вирусы оцРНК с положительным смыслом . Группа
В этой группе признаны три отряда и 34 семейства. Кроме того, существует ряд неклассифицированных видов и родов.
- Заказать Нидовиралеса
- Семейство Arteriviridae
- Семейство Coronaviridae - включает коронавирус человека (вирусы простуды HCoV-229E , HCoV-HKU1 , HCoV-NL63 и HCoV-OC43 ), MERS-CoV , SARS-CoV-1 и SARS-CoV-2.
- Семейство Мезонивирусиды
- Семейство Ронивирусиды
- Заказать Пикорнавирусы
- Семейство дицистровириды.
- Семейство Ифлавирусиды
- Семейство Марнавирусиды.
- Семейство Picornaviridae – включает полиовирус , риновирус (вирус простуды), гепатита А. вирус
- Семейство Secoviridae включает подсемейство Comovirinae.
- Род Bacillariornavirus
- Вид Вирус мухи ламинарии
- Заказать Тимовирал
- Семейство Alphaflexiviridae
- Семейство Betaflexiviridae.
- Семейство Gammaflexiviridae.
- Семейство Tymoviridae
- Неназначенный
- Семейство Alphatetraviridae
- Семейство Альвернавирусиды.
- Семейство Astroviridae
- Семейство Barnaviridae.
- Семейство Бенивирусиды
- Семейство Ботурмиавирусиды.
- Семейство Бромовирусиды
- Семейство Caliciviridae - включает вирус Норуолк.
- Семейство Carmotetraviridae.
- Семейство Closteroviridae
- Семейство Flaviviridae – включает вирус желтой лихорадки , вирус Западного Нила , вирус гепатита С , лихорадки Денге вирус , вирус Зика.
- Семейство Fusariviridae.
- Семейство Hepeviridae
- Семейство гиповирусов
- Семейство Левивирусиды.
- Семейство Luteoviridae - включает вирус желтой карликовости ячменя.
- Семейство Polycipiviridae
- Семейство Нарнавирусиды.
- Семейство Нодавирусиды
- Семейство Permutotetraviridae
- Семейство Потивирусиды
- Семейство Сартровирусиды.
- Семейный статовирус
- Семейство Togaviridae – включает вирус краснухи , вирус Росс-Ривер , вирус Синдбис , вирус Чикунгунья.
- Семейство Tombusviridae
- Семейство Virgaviridae [35]
- Неназначенные роды
- Род Блюнервирус
- Род Цилевирус
- Род Хигревирус
- Род Идеовирус
- Род Негевирус
- Род Ourmiavirus
- Род Полемовирус
- Род Синайвирус
- Род Собемовирус
- Неназначенные виды
- Вирус Acyrthosiphon pisum
- Бастровирус
- Вирус Блэкфорда
- Вирус некротической кольцевой пятнистости черники
- Кадисистровирус
- Вирус Chara australis
- Очень маленький вирус
- Вирус хлороза ягод годжи
- Вирус Harmonia axyridis 1
- Гепеливирус
- Клещевой вирус Цзинмэнь
- Белый вирус
- Недицистровирус
- Вирус Nesidiocorus тонкий 1
- Нифлавирус
- Вирус желтой ниландерии 1
- Вирус Орсе
- РНК-вирус 1 Osedax japonicus
- Пикаливирус
- Нидовирус секреторных клеток планарий
- Вирус Plasmopara halstedii
- Розеллиния некатрикс фузаривирус 1
- Вирус Сантей
- секаливирус
- Вирус Solenopsis invicta 3
- Уханьский вирус круглых червей крупных свиней
Спутниковые вирусы
- Семейство Сартровирусиды.
- Род Альбетовирус
- Род Аумаивирус
- Род Папанивирус
- Род Виртовирус
- Вирус хронического паралича пчел
Также был описан неклассифицированный вирус, подобный астровирусу/гепевирусу. [36]
Группа V – вирусы оцРНК с смыслом . отрицательным
За исключением вируса гепатита D , эту группу вирусов отнесли к одному типу — Negarnaviricota . Этот тип разделен на два подтипа — Haploviricotina и Polyploviricotina . В подтипе Haploviricotina в настоящее время выделяются четыре класса: Chunqiuviricetes , Milneviricetes , Monjiviricetes и Yunchangviricetes . В подтипе Polyploviricotina выделяют два класса: Ellioviricetes и Insthoviricetes .
В настоящее время в этой группе признаны шесть классов, семь орденов и двадцать четыре семейства. Ряд неустановленных видов и родов еще предстоит классифицировать. [10]
- Тип Негарнавирикота [37]
- Подтип Haploviricotina
- Класс Chunqiuviricetes
- Заказать Мувиралес
- Семейство Qinviridae
- Заказать Мувиралес
- Класс Милневирицеты
- Заказать Серпентовирусы
- Семейство Aspiviridae
- Заказать Серпентовирусы
- Класс Мондживирицетес
- Заказать Jingchuvirales
- Семейство Чувириды.
- Заказать мононегавирусы
- Семейство Bornaviridae - вирус болезни Борна.
- Семейство Filoviridae - включает Эбола вирус , вирус Марбург.
- Семейство Mymonaviridae
- Семейство Нямивириды. [38]
- Семейство Paramyxoviridae – включает вирус кори , вирус паротита , вирус Нипах , вирус Хендра и вирус NDV.
- Семейство Pneumoviridae - включает RSV и метапневмовирус.
- Семейство Rhabdoviridae - включает бешенства. вирус
- Семейство Sunviridae
- Род Анфевирус
- Род Арливирус
- Род Ченгтивирус
- Род Круставирус
- Род Вастривирус
- Заказать Jingchuvirales
- Класс Юнчанвирицетес
- Заказать Goujianvirales
- Семейство Юэвириды
- Заказать Goujianvirales
- Класс Chunqiuviricetes
- Подтип Polyploviricotina
- Класс Эллиовирицетес
- Заказать буньявирусы
- Семейство Arenaviridae - включает вирус Ласса.
- Семейство Cruliviridae
- Семейство Feraviridae
- Семейство Фимовирусиды.
- Семейство Хантавирусиды
- Семейство Jonviridae
- Семейство Nairoviridae
- Семейство Peribunyaviridae.
- Семейство фазмавирусов.
- Семейство фенуивириды.
- Семейство Тосповирусиды
- Род Тилапиневирусиды.
- Заказать буньявирусы
- Класс Insthoviricetes
- Заказать Articulavirales
- Семейство Amnoonviridae - включает вирус Тааструп.
- Семейство Orthomyxoviridae - включает гриппа . вирусы
- Заказать Articulavirales
- Класс Эллиовирицетес
- Подтип Haploviricotina
- Неназначенные роды:
- Род Deltavirus – включает вирус гепатита D (не настоящий вирус, а субвирусный агент ).
Галерея [ править ]






 Корь ( Парамиксовирусы )
Корь ( Парамиксовирусы )
















См. также [ править ]
- Классификация вирусов
- Список вирусов
- Репликация вируса
- Позитивный/отрицательный смысл
- Вирусы животных
- Вирусы с двухцепочечной РНК
- Ретровирус
- ДНК-вирусы
- Репликационный элемент цис-действия норовируса
- Вироид
Примечания [ править ]
- ^ Это включение произошло из-за TaxoProp 2017.006G , в котором предлагалась рибовирия . Путаница может возникнуть из-за ссылки TaxoProp на «монофилию всех РНК-вирусов», неправильно названную, поскольку она была продемонстрирована только с помощью RdRP. С другой стороны, в предложенном определении рибовирии правильно упоминается RdRP..
- ^ Большинство грибковых вирусов представляют собой вирусы с двухцепочечной РНК. Описано небольшое количество РНК-вирусов с положительной цепью. В одном отчете высказывается предположение о возможности существования вируса с отрицательной цепью. [8]
Ссылки [ править ]
- ^ Вагнер, Эдвард К.; Хьюлетт, Мартинес Дж. (1999). Базовая вирусология . Молден, Массачусетс: Blackwell Science, Inc., с. 249. ИСБН 0-632-04299-0 . Проверено 30 марта 2020 г.
- ^ «Список в таксономическом порядке - указатель списков видов ICTV» . Проверено 11 апреля 2008 г.
- ^ Исполнительный комитет Международного комитета по таксономии вирусов (май 2020 г.). «Новая сфера таксономии вирусов: разделение виросферы на 15 иерархических рангов» . Природная микробиология . 5 (5): 668–674. дои : 10.1038/s41564-020-0709-x . ПМК 7186216 . ПМИД 32341570 .
- ^ TaxoProp 2019.006G
- ^ TaxoProp 2019.009G
- ^ Нгуен М., Хэнни А.Л. (июнь 2003 г.). «Стратегии экспрессии амбисенс-вирусов». Вирусные исследования . 93 (2): 141–50. дои : 10.1016/S0168-1702(03)00094-7 . ПМИД 12782362 .
- ^ Кондо Х., Тиба С., Тойода К., Сузуки Н. (январь 2013 г.). «Доказательства заражения грибов вирусом с отрицательной цепью РНК» . Вирусология . 435 (2): 201–09. дои : 10.1016/j.virol.2012.10.002 . ПМИД 23099204 .
- ^ Санхуан Р., Небот М.Р., Кирико Н., Мански Л.М., Белшоу Р. (октябрь 2010 г.). «Темпы вирусных мутаций» . Журнал вирусологии . 84 (19): 9733–48. дои : 10.1128/JVI.00694-10 . ПМЦ 2937809 . ПМИД 20660197 .
- ↑ Перейти обратно: Перейти обратно: а б с Кляйн Д.В., Прескотт Л.М., Харли Дж. (1993). Микробиология . Дубьюк, Айова: Wm. К. Браун. ISBN 978-0-697-01372-9 .
- ^ Штайнхауэр Д.А., Холланд Дж.Дж. (1987). «Быстрая эволюция РНК-вирусов». Ежегодный обзор микробиологии . 41 : 409–33. дои : 10.1146/annurev.mi.41.100187.002205 . ПМИД 3318675 .
- ^ Бутвелл К.Л., Роллан М.М., Хербек Дж.Т., Маллинз Дж.И., Аллен Т.М. (октябрь 2010 г.). «Эволюция вируса и его побег во время острой инфекции ВИЧ-1» . Журнал инфекционных болезней . 202 (Приложение 2): S309–14. дои : 10.1086/655653 . ПМЦ 2945609 . ПМИД 20846038 .
- ^ Бух Дж., Перселл Р.Х., Миллер Р.Х. (август 1994 г.). «Анализ последовательности основного гена 14 генотипов вируса гепатита С» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (17): 8239–43. Бибкод : 1994PNAS...91.8239B . дои : 10.1073/pnas.91.17.8239 . ПМК 44581 . ПМИД 8058787 .
- ^ Туплин А., Эванс DJ, Симмондс П. (октябрь 2004 г.). «Детальное картирование вторичных структур РНК в последовательностях ядра и области, кодирующей NS5B, вируса гепатита С путем расщепления РНКазой и новых методов биоинформатического прогнозирования» . Журнал общей вирусологии . 85 (Часть 10): 3037–47. дои : 10.1099/vir.0.80141-0 . ПМИД 15448367 .
- ↑ Перейти обратно: Перейти обратно: а б Сильва Х.М., Пратас Д., Каэтано Т., Матос Д. (август 2022 г.). «Сложность вирусных геномов» . ГигаСайенс . 11 : 1–16. doi : 10.1093/gigascience/giac079 . ПМЦ 9366995 . ПМИД 35950839 .
- ↑ Перейти обратно: Перейти обратно: а б Барр Дж. Н., Фернс Р. (июнь 2010 г.). «Как РНК-вирусы сохраняют целостность своего генома» . Журнал общей вирусологии . 91 (Часть 6): 1373–87. дои : 10.1099/vir.0.020818-0 . ПМИД 20335491 .
- ^ Стедман, Кеннет М. (2015). «Глубокая рекомбинация: гены вируса РНК и оцДНК в ДНК-вирусе и геномах хозяина» . Ежегодный обзор вирусологии . 2 (1): 203–217. doi : 10.1146/annurev-virology-100114-055127 . ISSN 2327-0578 . ПМИД 26958913 . S2CID 207745438 .
- ^ Муслин С., Мак Кейн А., Бессо М., Блондель Б., Дельпейру Ф. (сентябрь 2019 г.). «Рекомбинация энтеровирусов: многоэтапный модульный эволюционный процесс» . Вирусы . 11 (9): 859. дои : 10.3390/v11090859 . ПМК 6784155 . ПМИД 31540135 .
- ^ Ху В.С., Темин Х.М. (ноябрь 1990 г.). «Ретровирусная рекомбинация и обратная транскрипция». Наука . 250 (4985): 1227–33. Бибкод : 1990Sci...250.1227H . дои : 10.1126/science.1700865 . ПМИД 1700865 .
- ^ Роусон Дж.М., Николаичик О.А., Кил Б.Ф., Патак В.К., Ху В.С. (ноябрь 2018 г.). «Рекомбинация необходима для эффективной репликации ВИЧ-1 и поддержания целостности вирусного генома» . Исследования нуклеиновых кислот . 46 (20): 10535–45. дои : 10.1093/nar/gky910 . ПМК 6237782 . ПМИД 30307534 .
- ↑ Перейти обратно: Перейти обратно: а б Бернштейн Х., Бернштейн К., Мишод Р.Э. (январь 2018 г.). «Пол у микробных патогенов» . Инфекция, генетика и эволюция . 57 : 8–25. дои : 10.1016/j.meegid.2017.10.024 . ПМИД 29111273 .
- ↑ Перейти обратно: Перейти обратно: а б Су С., Вонг Г., Ши В., Лю Дж., Лай А.С., Чжоу Дж. и др. (июнь 2016 г.). «Эпидемиология, генетическая рекомбинация и патогенез коронавирусов» . Тенденции в микробиологии . 24 (6): 490–502. дои : 10.1016/j.tim.2016.03.003 . ПМК 7125511 . ПМИД 27012512 .
- ^ Вольф Ю.И., Казлаускас Д., Иранзо Дж., Люсия-Санс А., Кун Дж.Х., Крупович М., Доля В.В., Кунин Е.В. (ноябрь 2018 г.). «Происхождение и эволюция глобального РНК-вирома» . мБио 9 (6). дои : 10.1128/mBio.02329-18 . ПМК 6282212 . ПМИД 30482837 .
- ^ Франки Р.И., Фоке С.М., Кнудсон Д.Л., Браун Ф. (1991). Классификация и номенклатура вирусов. Пятый отчет Международного комитета по таксономии вирусов, Архивы вирусологии (Приложение 2) . Спрингер. ISBN 978-3-7091-9163-7 .
- ^ «Текущий выпуск таксономии ICTV | ICTV» . ictv.global . Проверено 3 апреля 2023 г.
- ^ Кунин Е.В. (сентябрь 1991 г.). «Филогения РНК-зависимых РНК-полимераз РНК-вирусов с положительной цепью» . Журнал общей вирусологии . 72 (Часть 9): 2197–206. дои : 10.1099/0022-1317-72-9-2197 . ПМИД 1895057 .
- ^ Розанов М.Н., Кунин Е.В., Горбаленя А.Е. (август 1992 г.). «Консервация предполагаемого домена метилтрансферазы: отличительная черта «синдбис-подобной» супергруппы РНК-вирусов с положительной цепью». Журнал общей вирусологии . 73 (Часть 8): 2129–34. CiteSeerX 10.1.1.532.7367 . дои : 10.1099/0022-1317-73-8-2129 . ПМИД 1645151 .
- ^ Кунин Е.В., Доля В.В. (1993). «Эволюция и таксономия РНК-вирусов с положительной цепью: последствия сравнительного анализа аминокислотных последовательностей». Критические обзоры по биохимии и молекулярной биологии . 28 (5): 375–430. дои : 10.3109/10409239309078440 . ПМИД 8269709 .
- ^ Уорд CW (1993). «Прогресс к более высокой систематике вирусов» . Исследования в области вирусологии . 144 (6): 419–53. дои : 10.1016/S0923-2516(06)80059-2 . ПМЦ 7135741 . ПМИД 8140287 .
- ^ Кунин Е.В., Вольф Ю.И., Нагасаки К., Доля В.В. (декабрь 2008 г.). «Большой взрыв эволюции пикорноподобных вирусов предшествовал появлению супергрупп эукариот» . Обзоры природы. Микробиология . 6 (12): 925–39. дои : 10.1038/nrmicro2030 . ПМИД 18997823 .
- ↑ Перейти обратно: Перейти обратно: а б Габриал С.А. (1998). «Происхождение, адаптация и пути эволюции грибных вирусов» . Гены вирусов . 16 (1): 119–31. дои : 10.1023/а:1007966229595 . ПМК 7089520 . ПМИД 9562896 .
- ^ Жибрат Ж.Ф., Мариадассу М., Будино П., Дельмас Б. (июль 2013 г.). «Анализ излучения бирнавирусов из различных типов хозяев и их эволюционного сходства с другими вирусами с двухцепочечной РНК и положительной РНК-цепью с использованием надежного структурного выравнивания множественных последовательностей и передовых филогенетических методов» . Эволюционная биология BMC . 13 (1): 154. Бибкод : 2013BMCEE..13..154G . дои : 10.1186/1471-2148-13-154 . ПМЦ 3724706 . ПМИД 23865988 .
- ^ Ли CX, Ши М, Тянь Дж.Х., Лин XD, Кан Ю.Дж., Чен Л.Дж. и др. (январь 2015 г.). «Беспрецедентное геномное разнообразие РНК-вирусов у членистоногих указывает на происхождение РНК-вирусов с отрицательным смыслом» . электронная жизнь . 4 . doi : 10.7554/eLife.05378 . ПМЦ 4384744 . ПМИД 25633976 .
- ^ Крупович М., Кун Дж.Х., Фишер М.Г. (январь 2016 г.). «Система классификации вирофагов и сателлитных вирусов» . Архив вирусологии . 161 (1): 233–47. дои : 10.1007/s00705-015-2622-9 . hdl : 11858/00-001M-0000-0028-DC34-F . ПМИД 26446887 .
- ^ Адамс М.Ю., Антонив Дж.Ф., Кройце Дж. (2009). «Virgaviridae: новое семейство палочковидных вирусов растений» . Архив вирусологии . 154 (12): 1967–72. дои : 10.1007/s00705-009-0506-6 . ПМИД 19862474 .
- ^ Панкович П., Борос А., Кисс Т., Энгельманн П., Рейтер Г. (2019) Генетически сильно дивергентный РНК-вирус с астровирусоподобной (5'-конец) и гепевирусоподобной (3'-конец) организацией генома у плотоядных птиц, европейцы. валик ( Coracias garrulus ). Заразить Генет Эвол
- ^ «Таксономия вирусов: выпуск 2018 г.» . Международный комитет по таксономии вирусов . Проверено 13 ноября 2018 г.
- ^ Михиндукуласурия К.А., Нгуен Н.Л., Ву Г, Хуанг Х.В., да Роза А.П., Попов В.Л. и др. (май 2009 г.). «Маленькие и промежуточные вирусы определяют новый таксон РНК-вирусов отряда Mononegavirales» . Журнал вирусологии . 83 (10): 5109–16. дои : 10.1128/JVI.02667-08 . ПМК 2682064 . ПМИД 19279111 .
Внешние ссылки [ править ]
- РНК-вирусы Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- Вирусы животных
