Полиомавирусы
| Полиомавирусы | |
|---|---|
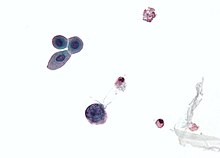 | |
| Микрофотография , показывающая клетку, инфицированную полиомавирусом : большая (синяя) клетка внизу по центру слева. Цитологический образец мочи. | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Моноднавирия |
| Королевство: | Шотокувираэ |
| Тип: | Коссавирикота |
| Сорт: | Паповавирицетес |
| Заказ: | Сеполивирусы |
| Семья: | Полиомавирусы |
| Роды | |
Polyomaviridae — семейство вирусов , естественными хозяевами которых являются преимущественно млекопитающие и птицы. [1] [2] По состоянию на 2024 год существует восемь признанных родов. [3] Известно, что 14 видов заражают человека, тогда как другие, такие как обезьяний вирус 40 , выявляются у людей в меньшей степени. [4] [5] Большинство этих вирусов очень распространены и обычно протекают бессимптомно в большинстве изученных популяций человека. [6] [7] Вирус BK связан с нефропатией у пациентов с трансплантацией почек и пациентов с трансплантацией твердых органов без почек. [8] [9] JC-вирус с прогрессирующей мультифокальной лейкоэнцефалопатией , [10] и вирус клеток Меркеля с раком клеток Меркеля . [11]
Структура и геном
[ редактировать ]
Полиомавирусы представляют собой безоболочечные двухцепочечные ДНК- вирусы с кольцевыми геномами длиной около 5000 пар оснований . Геном упакован в вирусный капсид диаметром около 40-50 нанометров , имеющий икосаэдрическую форму (симметрия Т=7). [2] [12] Капсид состоит из 72 пентамерных капсомеров белка VP1 , который способен к самосборке в закрытый икосаэдр; [13] каждый пентамер VP1 связан с одной молекулой одного из двух других капсидных белков, VP2 или VP3 . [5]

Геном типичного полиомавируса кодирует от 5 до 9 белков , разделенных на две области транскрипции, называемые ранними и поздними областями, в зависимости от времени, в течение которого они транскрибируются. Каждая область транскрибируется РНК-полимеразой II клетки-хозяина как одна пре-мессенджерная РНК, содержащая несколько генов. Ранняя область обычно кодирует два белка, малый и большой опухолевые антигены, образующиеся путем альтернативного сплайсинга . Поздняя область содержит три структурных белка капсида VP1, VP2 и VP3, продуцируемые альтернативными сайтами начала трансляции . Дополнительные гены и другие вариации на эту тему присутствуют в некоторых вирусах: например, полиомавирусы грызунов имеют третий белок, называемый средним опухолевым антигеном, в ранней области, который чрезвычайно эффективен в индукции клеточной трансформации ; SV40 имеет дополнительный капсидный белок VP4; в некоторых примерах имеется дополнительный регуляторный белок, называемый агнопротеином, экспрессируемый из поздней области. Геном также содержит некодирующая ранней и поздней областей контрольная или регуляторная область, содержащая промоторы , сайты начала транскрипции и точку начала репликации . [2] [12] [5] [15]
Репликация и жизненный цикл
[ редактировать ]
Жизненный цикл полиомавируса начинается с проникновения в клетку-хозяина . Клеточные рецепторы полиомавирусов представляют собой сиаловых кислот остатки гликанов , обычно ганглиозидов . Прикрепление полиомавирусов к клеткам-хозяевам опосредуется связыванием VP1 с сиалилированными гликанами на поверхности клетки. [2] [12] [15] [16] В некоторых конкретных вирусах происходят дополнительные взаимодействия клетки с поверхностью; например, вирус JC считается, что требует взаимодействия с рецептором 5HT2A , а вирус клеток Меркеля - с гепарансульфатом . [15] [17] Однако в целом взаимодействия вирус-клетка опосредуются часто встречающимися молекулами на поверхности клетки и, следовательно, вероятно, не вносят основного вклада в наблюдаемый тропизм клеточного типа отдельных вирусов . [15] После связывания с молекулами на поверхности клетки вирион подвергается эндоцитозу и попадает в эндоплазматический ретикулум – поведение, уникальное среди известных вирусов без оболочки. [18] - где структура вирусного капсида может быть нарушена действием ферментов дисульфид-изомеразы клетки-хозяина . [2] [12] [19]
Детали транзита в ядро неясны и могут различаться у разных полиомавирусов. Часто сообщалось, что неповрежденная, хотя и искаженная, частица вириона высвобождается из эндоплазматического ретикулума в цитоплазму клетки, где геном высвобождается из капсида, возможно, из-за низкой концентрации кальция в цитоплазме. [18] Как экспрессия вирусных генов, так и репликация вирусного генома происходят в ядре с использованием механизмов клетки-хозяина. Ранние гены, включающие как минимум малый опухолевый антиген (ST) и большой опухолевый антиген (LT), экспрессируются первыми из одной альтернативно сращенной цепи информационной РНК . хозяина Эти белки служат для управления клеточным циклом , нарушая регуляцию перехода от фазы G1 к фазе S , когда геном клетки-хозяина реплицируется, поскольку для репликации вирусного генома необходим механизм репликации ДНК клетки-хозяина. [2] [12] [15] Точный механизм этого нарушения регуляции зависит от вируса; например, SV40 LT может напрямую связывать p53 клетки-хозяина , но LT мышиного полиомавируса этого не делает. [20] LT индуцирует репликацию ДНК из некодирующей контрольной области вирусного генома (NCCR), после чего экспрессия ранней мРНК снижается и начинается экспрессия поздней мРНК, кодирующей белки вирусного капсида. [19] Когда эти взаимодействия начинаются, LT, принадлежащие нескольким полиомавирусам, включая полиомавирус клеток Меркеля , представляют онкогенный потенциал. [21] Описано несколько механизмов регуляции перехода от ранней к поздней экспрессии генов, включая участие белка LT в репрессии раннего промотора. [19] экспрессия нетерминированных поздних мРНК с расширениями, комплементарными ранней мРНК, [15] и экспрессия регуляторных микроРНК . [15] Экспрессия поздних генов приводит к накоплению белков вирусного капсида в цитоплазме клетки-хозяина. Компоненты капсида проникают в ядро, чтобы инкапсидировать новую вирусную геномную ДНК. Новые вирионы могут быть собраны на вирусных фабриках . [2] [12] Механизм высвобождения вируса из клетки-хозяина различается у разных полиомавирусов; некоторые экспрессируют белки, которые облегчают выход из клетки, такие как агнопротеин или VP4 . [19] В некоторых случаях высокие уровни инкапсидированного вируса приводят к лизису клеток с высвобождением вирионов. [15]
Вирусные белки
[ редактировать ]Опухолевые антигены
[ редактировать ]Большой опухолевый антиген играет ключевую роль в регулировании жизненного цикла вируса, связываясь с вирусным источником репликации ДНК, где он способствует синтезу ДНК. Кроме того, поскольку полиомавирус полагается на механизмы клетки-хозяина для репликации, клетка-хозяин должна находиться в s-фазе, чтобы это началось. В связи с этим большой Т-антиген также модулирует клеточные сигнальные пути, стимулируя прогресс клеточного цикла путем связывания с рядом белков клеточного контроля. [22] Это достигается за счет двухстороннего воздействия: ингибирования генов, подавляющих опухоль, p53 и членов семейства ретинобластомы (pRB). [23] и стимуляцию путей роста клеток путем связывания клеточной ДНК, ассоциации АТФазы-хеликазы, ДНК-полимеразы α и связывания факторов комплекса преинициации транскрипции. [24] Эта аномальная стимуляция клеточного цикла является мощной силой для онкогенной трансформации. [ нужна ссылка ]
Небольшой опухолевый антигенный белок также способен активировать несколько клеточных путей, которые стимулируют пролиферацию клеток. Малые Т-антигены полиомавируса обычно нацелены на протеинфосфатазу 2А ( PP2A ), [25] ключевой мультисубъединичный регулятор нескольких путей, включая Akt , путь митоген-активируемой протеинкиназы (MAPK) и путь активируемой стрессом протеинкиназы (SAPK). [26] [27] Малый Т-антиген полиомавируса клеток Меркеля кодирует уникальный домен, называемый доменом LT-стабилизации (LSD), который связывается и ингибирует FBXW7, лигазу E3 регулирующую как клеточные, так и вирусные онкопротеины. [28] В отличие от SV40, малый Т-антиген MCV напрямую трансформирует клетки грызунов in vitro. [29]
Средний опухолевый антиген используется в модельных организмах, разработанных для изучения рака, таких как система MMTV-PyMT , где средний Т связан с MMTV промотором . Там он функционирует как онкоген , а ткань, в которой развивается опухоль, определяется промотором MMTV. [ нужна ссылка ]
Капсидные белки
[ редактировать ]Капсид полиомавируса состоит из одного основного компонента, основного капсидного белка VP1 , и одного или двух второстепенных компонентов, минорных капсидных белков VP2 и VP3 . VP1 Пентамеры образуют закрытый икосаэдрический вирусный капсид , а внутри капсида каждый пентамер связан с одной молекулой либо VP2, либо VP3. [5] [30] Некоторые полиомавирусы, такие как полиомавирус клеток Меркеля , не кодируют и не экспрессируют VP3. [31] Белки капсида экспрессируются из поздней области генома. [5]
Агнопротеин
[ редактировать ]Агнопротеин представляет собой небольшой многофункциональный фосфопротеин , обнаруженный в поздней кодирующей части генома некоторых полиомавирусов, особенно вируса BK , вируса JC и SV40 . Он важен для пролиферации вирусов, которые его экспрессируют, и считается, что он участвует в регуляции жизненного цикла вируса, особенно репликации и выхода вируса из клетки-хозяина, но точные механизмы неясны. [32] [33]
Таксономия
[ редактировать ]Полиомавирусы относятся к группе I (дцДНК-вирусы). Классификация полиомавирусов была предметом нескольких предлагаемых пересмотров по мере обнаружения новых членов группы. Раньше полиомавирусы и папилломавирусы , которые имели много общих структурных особенностей, но имели очень разные геномные организации, были классифицированы вместе в ныне устаревшем семействе Papovaviridae . [34] (Название Papovaviridae происходит от трех аббревиатур: Pa — папилломавирус , Po — полиомавирус и Va — «вакуолирующий».) [35] Полиомавирусы были разделены на три основные клады (то есть генетически родственные группы): кладу SV40, кладу птичьего вируса и кладу мышиного полиомавируса. [36] Последующая реклассификация, предложенная Международным комитетом по таксономии вирусов (ICTV), рекомендовала разделить семейство Polyomaviridae на три рода: [37]
- Род Orthopolyomavirus (типовой вид SV40 ).
- Род Wukipolyomavirus (типовой вид полиомавируса KI )
- Род Avipolyomavirus (типовой вид Avianpolyomavirus )
Текущая система классификации ICTV признает шесть родов и 117 видов, из которых пяти нельзя отнести к роду. Эта система сохраняет различие между вирусами птиц и млекопитающих, группируя птичью подгруппу в род Gammapolyomavirus . Шесть родов: [38]
- Альфаполиомавирус
- Бетаполиомавирус
- Дельтаполиомавирус
- Эпсилонполиомавирус
- Гаммаполиомавирус
- Зетаполиомавирус
Следующие виды не отнесены к роду: [38]
- Полиомавирус Centropristis striata 1
- Полиомавирус Rhynchobatus jiddensis 1
- Полиомавирус Sparus aurata 1
- Полиомавирус Trematomus Bernacchii 1
- Полиомавирус Trematomus pennellii 1
Описание дополнительных вирусов продолжается. К ним относятся полиомавирус калана 1. [39] и полиомавирус Альпака [40] Другой вирус — полиомавирус гигантской панды 1. [41] Другой вирус был описан у сигмодонтных грызунов. [42] Другой - полиомавирус древесной землеройки 1 - был описан у землероек. [43]
Полиомавирусы человека
[ редактировать ]Большинство полиомавирусов не заражают человека. Из полиомавирусов, каталогизированных по состоянию на 2017 год, в общей сложности было известно 14 с человеческими хозяевами. [4] Однако некоторые полиомавирусы связаны с заболеваниями человека, особенно у с ослабленным иммунитетом лиц . MCV сильно отличается от других полиомавирусов человека и наиболее тесно связан с мышиным полиомавирусом. Полиомавирус, ассоциированный с триходисплазией , имеет отдаленное родство с MCV. Два вируса — HPyV6 и HPyV7 — наиболее тесно связаны с вирусами KI и WU, тогда как HPyV9 наиболее тесно связан с лимфотропным полиомавирусом африканской зеленой обезьяны (LPV). [ нужна ссылка ]
Описан четырнадцатый вирус. [44] Полиомавирус Lyon IARC родственен полиомавирусу енота. [ нужна ссылка ]
Список полиомавирусов человека
[ редактировать ]По состоянию на 2017 год были идентифицированы следующие 14 полиомавирусов с человеческими хозяевами, и их геномы секвенированы: [4]
| Разновидность | Предлагаемый род | Имя вируса | Аббревиатура | NCBI RefSeq | Год открытия | Клинический коррелят (если есть) | Ссылки |
|---|---|---|---|---|---|---|---|
| Полиомавирус человека 5 | Альфа | Полиомавирус клеток Меркеля | MCPyV | NC_010277 | 2008 | Рак клеток Меркеля [5] | [45] [11] [46] |
| Полиомавирус человека 8 | Альфа | Полиомавирус триходисплазии спинулезной | ЦПыВ | NC_014361 | 2010 | Триходисплазия шиповатая [5] | [47] [48] |
| Полиомавирус человека 9 | Альфа | Полиомавирус человека 9 | HPyV9 | NC_015150 | 2011 | Ничего не известно | [49] |
| Полиомавирус человека 12 | Альфа | Полиомавирус человека 12 | HPyV12 | NC_020890 | 2013 | Ничего не известно | [50] |
| Полиомавирус человека 13 | Альфа | Полиомавирус Нью-Джерси | НДжПиВ | NC_024118 | 2014 | Ничего не известно | [51] |
| Полиомавирус человека 1 | Бета | БК полиомавирус | БКПыВ | NC_001538 | 1971 | полиомавирус-ассоциированная нефропатия ; геморрагический цистит [5] | [52] |
| Полиомавирус человека 2 | Бета | JC полиомавирус | JCPyV | NC_001699 | 1971 | Прогрессирующая мультифокальная лейкоэнцефалопатия [5] | [53] |
| Полиомавирус человека 3 | Бета | КИ полиомавирус | КИПыВ | NC_009238 | 2007 | Ничего не известно | [54] |
| Полиомавирус человека 4 | Бета | ВУ полиомавирус | ВУПыВ | NC_009539 | 2007 | Ничего не известно | [14] |
| Полиомавирус человека 6 | Дельта | Полиомавирус человека 6 | HPyV6 | NC_014406 | 2010 | HPyV6-ассоциированный зудящий и дискератотический дерматоз (H6PD) [55] | [31] |
| Полиомавирус человека 7 | Дельта | Полиомавирус человека 7 | HPyV7 | NC_014407 | 2010 | HPyV7-ассоциированная эпителиальная гиперплазия [55] [56] [57] | [31] |
| Полиомавирус человека 10 | Дельта | полиомавирус MW | МВПиВ | NC_018102 | 2012 | Ничего не известно | [58] [59] [60] |
| Полиомавирус человека 11 | Дельта | STL полиомавирус | СТЛПиВ | NC_020106 | 2013 | Ничего не известно | [61] |
| Полиомавирус человека 14 | Альфа | Полиомавирус Лиона IARC | ЛИПыВ | NC_034253.1 | 2017 | Ничего не известно | [62] [63] |
Дельтаполиомавирус содержит только четыре человеческих вируса, показанных в таблице выше. Группы Альфа и Бета содержат вирусы, поражающие различных млекопитающих. Группа Гамма содержит птичьи вирусы. [4] Клинически значимые ассоциации заболеваний показаны только там, где ожидается причинно-следственная связь. [5] [64]
У людей были обнаружены антитела к лимфотропному полиомавирусу обезьян, что позволяет предположить, что этот вирус или близкородственный вирус могут заражать людей. [65]
Клиническая значимость
[ редактировать ]Все полиомавирусы являются широко распространенными инфекциями среди детей и молодых взрослых. [66] Большинство этих инфекций, по-видимому, практически не вызывают симптомов. Эти вирусы, вероятно, сохраняются на протяжении всей жизни почти у всех взрослых. Заболевания, вызванные полиомавирусной инфекцией человека, наиболее распространены среди с ослабленным иммунитетом людей ; ассоциации заболеваний включают вирус BK с нефропатией у пациентов с трансплантацией почки и пациентов с трансплантацией твердых органов без почки, [8] [9] JC-вирус с прогрессирующей мультифокальной лейкоэнцефалопатией , [10] и вирус клеток Меркеля (MCV) с раком клеток Меркеля . [11]
СВ40
[ редактировать ]SV40 размножается в почках обезьян , не вызывая заболевания, но может вызывать рак у грызунов в лабораторных условиях. В 1950-х и начале 1960-х годов более 100 миллионов человек могли подвергнуться воздействию SV40 из-за ранее необнаруженного заражения SV40 полиовакцины , что вызвало обеспокоенность по поводу возможности того, что вирус может вызвать заболевание у людей. [67] [68] Хотя сообщалось о его присутствии при некоторых видах рака человека, включая опухоли головного мозга , опухоли костей , мезотелиомы и неходжкинские лимфомы , [69] Точное обнаружение часто затрудняется высоким уровнем перекрестной реактивности SV40 с широко распространенными полиомавирусами человека. [68] Большинство вирусологов отвергают SV40 как причину рака у человека. [67] [70] [71]
Диагностика
[ редактировать ]Диагноз полиомавируса почти всегда ставится после первичной инфекции, поскольку она протекает либо бессимптомно, либо субклинически. Анализы на антитела обычно используются для обнаружения присутствия антител против отдельных вирусов. [72] Конкурентные анализы часто необходимы для различения очень похожих полиомавирусов. [73]
В случаях прогрессирующей мультифокальной лейкоэнцефалопатии (ПМЛ) перекрестно-реактивные антитела к Т-антигену SV40 (обычно Pab419) используются для непосредственного окрашивания тканей на наличие Т-антигена вируса JC. ПЦР можно использовать при биопсии ткани или спинномозговой жидкости для амплификации ДНК полиомавируса. Это позволяет не только обнаружить полиомавирус, но и определить его подтип. [74]
Для диагностики реактивации полиомавируса при полиомавирусной нефропатии (ПВН) используются три основных диагностических метода: цитология мочи, количественное определение вирусной нагрузки как в моче, так и в крови и биопсия почки . [72] Реактивация полиомавируса в почках и мочевыводящих путях приводит к выделению инфицированных клеток, вирионов и/или вирусных белков с мочой. Это позволяет цитологическому анализу мочи исследовать эти клетки, что, если в ядре имеется включение полиомавируса, является диагностическим признаком инфекции. [75] Кроме того, поскольку моча инфицированного человека будет содержать вирионы и/или вирусную ДНК, количественную оценку вирусной нагрузки можно провести с помощью ПЦР. [76] Это справедливо и для крови.
Биопсию почки также можно использовать, если два только что описанных метода не дали результатов или если желательно определить специфическую вирусную нагрузку для почечной ткани. Подобно цитологическому исследованию мочи, почечные клетки исследуют под световой микроскопией на предмет включения полиомавируса в ядро, а также лизис клеток и вирусные частицы во внеклеточной жидкости. Вирусную нагрузку, как и раньше, также измеряют методом ПЦР. [ нужна ссылка ]
Окрашивание тканей с использованием моноклональных антител против Т-антигена MCV показывает полезность для дифференциации клеточной карциномы Меркеля от других мелкоклеточных опухолей. [77] Были разработаны анализы крови для выявления антител к MCV, которые показывают, что заражение вирусом широко распространено, хотя у пациентов с карциномой Меркеля наблюдаются исключительно более высокие реакции антител, чем у людей, инфицированных бессимптомно. [7] [78] [79] [80]
Использование для отслеживания миграции людей.
[ редактировать ]Вирус JC представляет собой многообещающий генетический маркер эволюции и миграции человека. [81] Он переносится 70–90 процентами людей и обычно передается от родителей потомству. Этот метод не кажется надежным для отслеживания недавнего африканского происхождения современных людей . [ нужна ссылка ]
История
[ редактировать ]Мышиный полиомавирус сообщил был первым открытым полиомавирусом, о котором Людвик Гросс в 1953 году как экстракт мышиных лейкозов, способный вызывать опухоли околоушной железы . [82] Возбудитель был идентифицирован как вирус Сарой Стюарт и Бернис Эдди , в честь которых его когда-то назвали «полиомой SE». [83] [84] [85] Термин «полиома» относится к способности вирусов образовывать множественные (поли-) опухоли (-ома) при определенных условиях. Это название подверглось критике как «лингвистический сэндвич без мяса» («постный», потому что обе морфемы в слове «полиома» являются аффиксами), дающий мало понимания биологии вирусов; Фактически, последующие исследования показали, что большинство полиомавирусов редко вызывают клинически значимые заболевания в организмах-хозяевах в естественных условиях. [86]
По состоянию на 2017 год были идентифицированы и секвенированы десятки полиомавирусов, поражающих в основном птиц и млекопитающих. Известно, что два полиомавируса поражают рыбу, черноморского окуня. [87] и дорада морской лещ . [88] Всего известно четырнадцать полиомавирусов, которые заражают человека. [4]
Ссылки
[ редактировать ]- ^ Моенс Ю, Кальвиньяк-Спенсер С, Лаубер С, Рамквист Т, Фельткамп MC, Догерти MD, Вершур Э.Дж., Элерс Б (июнь 2017 г.). «Профиль таксономии вирусов ICTV: полиомавирусы» . Журнал общей вирусологии . 98 (6): 1159–1160. дои : 10.1099/jgv.0.000839 . ПМЦ 5656788 . ПМИД 28640744 .
- ^ Перейти обратно: а б с д и ж г «Отчет ICTV о полиомавирусах» .
- ^ таксономия. «Браузер таксономии (Polyomaviridae)» . www.ncbi.nlm.nih.gov . Проверено 19 марта 2024 г.
- ^ Перейти обратно: а б с д и Кальвиньяк-Спенсер С., Фельткамп М.К., Догерти М.Д., Моенс У., Рамквист Т., Джон Р., Элерс Б. (июнь 2016 г.). «Обновление таксономии семейства Polyomaviridae» . Архив вирусологии . 161 (6): 1739–50. дои : 10.1007/s00705-016-2794-y . hdl : 10037/13151 . ПМИД 26923930 .
- ^ Перейти обратно: а б с д и ж г час я дж ДеКаприо Дж.А., Гарсиа Р.Л. (апрель 2013 г.). «Рог изобилия полиомавирусов человека» . Обзоры природы. Микробиология . 11 (4): 264–76. дои : 10.1038/nrmicro2992 . ПМЦ 3928796 . ПМИД 23474680 .
- ^ Госсай А., Уотербур Т., Нельсон Х.Х., Мишель А., Уиллхок-Флекенштейн М., Фарзан С.Ф., Хоэн А.Г., Кристенсен Б.С., Келси К.Т., Марсит С.Дж., Павлита М., Карагас М.Р. (январь 2016 г.). «Сероэпидемиология полиомавирусов человека в популяции США» . Американский журнал эпидемиологии . 183 (1): 61–9. дои : 10.1093/aje/kwv155 . ПМК 5006224 . ПМИД 26667254 .
- ^ Перейти обратно: а б Кин Дж. М., Рао С., Ван М., Гарсиа Р.Л. (март 2009 г.). «Сероэпидемиология полиомавирусов человека» . ПЛОС Патогены . 5 (3): e1000363. дои : 10.1371/journal.ppat.1000363 . ПМЦ 2655709 . ПМИД 19325891 .
- ^ Перейти обратно: а б Джамботи Дж.С. (август 2016 г.). «ВК-вирусная нефропатия у реципиентов почечного трансплантата» . Нефрология . 21 (8): 647–54. дои : 10.1111/nep.12728 . ПМИД 26780694 .
- ^ Перейти обратно: а б Куппачи С., Каур Д., Холанда Д.Г., Томас К.П. (апрель 2016 г.). «Вирусная инфекция полиомы BK и заболевание почек при трансплантации твердых органов без почек» . Клинический журнал почек . 9 (2): 310–8. дои : 10.1093/ckj/sfv143 . ПМЦ 4792618 . ПМИД 26985385 .
- ^ Перейти обратно: а б Аданг Л., Бергер Дж. (2015). «Прогрессирующая мультифокальная лейкоэнцефалопатия» . F1000Исследования . 4 : 1424. doi : 10.12688/f1000research.7071.1 . ПМК 4754031 . ПМИД 26918152 .
- ^ Перейти обратно: а б с Фэн Х., Шуда М., Чанг Ю, Мур П.С. (февраль 2008 г.). «Клональная интеграция полиомавируса в клеточную карциному Меркеля человека» . Наука . 319 (5866): 1096–100. Бибкод : 2008Sci...319.1096F . дои : 10.1126/science.1152586 . ПМК 2740911 . ПМИД 18202256 .
- ^ Салунке Д.М., Каспар Д.Л., Гарсеа Р.Л. (сентябрь 1986 г.). «Самосборка очищенного капсидного белка полиомавируса VP1». Клетка . 46 (6): 895–904. дои : 10.1016/0092-8674(86)90071-1 . ПМИД 3019556 . S2CID 25800023 .
- ^ Перейти обратно: а б Гейнор А.М., Ниссен М.Д., Уилли Д.М., Маккей И.М., Ламберт С.Б., Ву Дж., Бреннан Д.С., Сторч Г.А., Слотс Т.П., Ван Д. (май 2007 г.). «Идентификация нового полиомавируса у больных острыми инфекциями дыхательных путей» . ПЛОС Патогены . 3 (5): е64. doi : 10.1371/journal.ppat.0030064 . ПМК 1864993 . ПМИД 17480120 .
- ^ Перейти обратно: а б с д и ж г час Международное агентство по исследованию рака (2013). «Введение в полиомавирусы» (PDF) . Монографии МАИР по оценке канцерогенных рисков для человека . 104 : 121–131.
- ^ Перейти обратно: а б Бух М.Х., Лиачи А.М., О'Хара С.Д., Гарсиа Р.Л., Ной У., Стел Т. (октябрь 2015 г.). «Структурный и функциональный анализ капсидных белков мышиного полиомавируса устанавливает детерминанты распознавания и патогенности лигандов» . ПЛОС Патогены . 11 (10): е1005104. дои : 10.1371/journal.ppat.1005104 . ПМЦ 4608799 . ПМИД 26474293 .
- ^ Шовальтер Р.М., Пастрана Д.В., Бак CB (июль 2011 г.). «Гликозаминогликаны и сиалилированные гликаны последовательно облегчают проникновение инфекционного полиомавируса в клетки Меркеля» . ПЛОС Патогены . 7 (7): e1002161. дои : 10.1371/journal.ppat.1002161 . ПМК 3145800 . ПМИД 21829355 .
- ^ Перейти обратно: а б Иноуэ Т., Цай Б. (январь 2013 г.). «Как вирусы используют эндоплазматический ретикулум для проникновения, репликации и сборки» . Перспективы Колд-Спринг-Харбор в биологии . 5 (1): а013250. doi : 10.1101/cshperspect.a013250 . ПМК 3579393 . ПМИД 23284050 .
- ^ Перейти обратно: а б с д Гёруп О, Чанг Й (2010). «Обновленная информация о полиомавирусах человека и раке». Достижения в области исследований рака . 106 : 1–51. дои : 10.1016/S0065-230X(10)06001-X . ISBN 9780123747716 . ПМИД 20399955 .
- ^ Андраби С., Хван Дж.Х., Чой Дж.К., Робертс Т.М., Шаффхаузен Б.С. (2011). «Сравнение мышиного полиомавируса и обезьяньего вируса 40 показывает значительные различия в функции малого Т-антигена» . Журнал вирусологии . 85 (20): 10649–10658. дои : 10.1128/JVI.05034-11 . ПМК 3187521 . ПМИД 21835797 .
- ^ Ротондо Х.К., Бонони И., Пуоццо А., Говони М., Фоски В., Ланца Г., Гафа Р., Габорио П., Тузе Ф.А., Селватичи Р., Мартини Ф., Тоньон М. (июль 2017 г.). «Карцинома из клеток Меркеля, возникающая у пациентов с аутоиммунными заболеваниями, получавших лечение биологическими препаратами, включая анти-ФНО» . Клинические исследования рака . 23 (14): 3929–3934. дои : 10.1158/1078-0432.CCR-16-2899 . hdl : 11392/2378829 . ПМИД 28174236 .
- ^ Уайт МК, Гордон Дж., Рейсс К., Дель Валле Л., Кроул С., Джордано А., Дарбинян А., Халили К. (декабрь 2005 г.). «Полиомавирусы человека и опухоли головного мозга». Исследования мозга. Обзоры исследований мозга . 50 (1): 69–85. дои : 10.1016/j.brainresrev.2005.04.007 . ПМИД 15982744 . S2CID 20990837 .
- ^ Казем С., ван дер Мейден Э., Ван Р.К., Розенберг А.С., Поуп Э., Бенуа Т., Флекман П., Фельткамп М.С. (2014). «Связанная с полиомавирусом Trichodysplasia spinulosa включает гиперпролиферацию, фосфорилирование pRB и активацию p16 и p21» . ПЛОС ОДИН . 9 (10): e108947. Бибкод : 2014PLoSO...9j8947K . дои : 10.1371/journal.pone.0108947 . ПМЦ 4188587 . ПМИД 25291363 .
- ^ Келли В.Л., Георгопулос С. (апрель 1997 г.). «Общий экзон T/t обезьяньего вируса 40, Т-антигены полиомавирусов JC и BK может функционально заменять J-домен молекулярного шаперона DnaJ Escherichia coli» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (8): 3679–84. Бибкод : 1997PNAS...94.3679K . дои : 10.1073/pnas.94.8.3679 . ПМК 20500 . ПМИД 9108037 .
- ^ Паллас Д.К., Шахрик Л.К., Мартин Б.Л., Джасперс С., Миллер Т.Б., Бротиган Д.Л., Робертс Т.М. (январь 1990 г.). «Малый и средний Т-антигены полиомы и малый Т-антиген SV40 образуют стабильные комплексы с протеинфосфатазой 2А». Клетка . 60 (1): 167–76. дои : 10.1016/0092-8674(90)90726-у . ПМИД 2153055 . S2CID 2007706 .
- ^ Зонтаг Э., Федоров С., Камибаяши С., Роббинс Д., Кобб М., Мамби М. (декабрь 1993 г.). «Взаимодействие антигена небольшой опухоли SV40 с протеинфосфатазой 2А стимулирует путь карт-киназы и индуцирует пролиферацию клеток» . Клетка . 75 (5): 887–97. дои : 10.1016/0092-8674(93)90533-В . ПМИД 8252625 .
- ^ Ватанабэ Г., Хоу А., Ли Р.Дж., Альбанезе С., Шу И.В., Карнезис А.Н., Зон Л., Кириакис Дж., Рунделл К., Пестель Р.Г. (ноябрь 1996 г.). «Индукция циклина D1 малым опухолевым антигеном обезьяньего вируса 40» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (23): 12861–6. Бибкод : 1996PNAS...9312861W . дои : 10.1073/pnas.93.23.12861 . ПМК 24011 . ПМИД 8917510 .
- ^ Квун Х.Дж., Шуда М., Фэн Х., Камачо С.Дж., Мур П.С., Чанг Ю (август 2013 г.). «Малый Т-антиген полиомавируса клеток Меркеля контролирует репликацию вируса и экспрессию онкопротеина, воздействуя на клеточную убиквитинлигазу SCFFbw7» . Клетка-хозяин и микроб . 14 (2): 125–35. дои : 10.1016/j.chom.2013.06.008 . ПМЦ 3764649 . ПМИД 23954152 .
- ^ Шуда М., Квун Х.Дж., Фэн Х., Чанг Ю., Мур П.С. (сентябрь 2011 г.). «Малый Т-антиген полиомавируса клеток Меркеля человека представляет собой онкопротеин, нацеленный на регулятор трансляции 4E-BP1» . Журнал клинических исследований . 121 (9): 3623–34. дои : 10.1172/JCI46323 . ПМК 3163959 . ПМИД 21841310 .
- ^ Чен XS, Стел Т, Харрисон С.С. (июнь 1998 г.). «Взаимодействие внутреннего белка VP2 полиомавируса с основным капсидным белком VP1 и значение участия VP2 во проникновении вируса» . Журнал ЭМБО . 17 (12): 3233–40. дои : 10.1093/emboj/17.12.3233 . ПМК 1170661 . ПМИД 9628860 .
- ^ Перейти обратно: а б с Шовальтер Р.М., Пастрана Д.В., Памфри К.А., Мойер А.Л., Бак CB (июнь 2010 г.). «Полиомавирус клеток Меркеля и два ранее неизвестных полиомавируса хронически выделяются с кожи человека» . Клетка-хозяин и микроб . 7 (6): 509–15. дои : 10.1016/j.chom.2010.05.006 . ПМЦ 2919322 . ПМИД 20542254 .
- ^ Сарыер И.К., Сарибас А.С., Уайт М.К., Сафак М. (май 2011 г.). «Заражение агнопротеин-негативными мутантами полиомавирусов JC и SV40 приводит к высвобождению вирионов, в большинстве случаев дефицитных по содержанию ДНК» . Вирусологический журнал . 8 : 255. дои : 10.1186/1743-422X-8-255 . ПМК 3127838 . ПМИД 21609431 .
- ^ Сарибас А.С., Чорич П., Хамазаспян А., Дэвис В., Аксман Р., Уайт М.К., Абу-Гарбиа М., Чайлдерс В., Кондра Дж.Х., Буазиз С., Сафак М. (октябрь 2016 г.). «Выход из неизведанного: структурные и функциональные особенности агнопротеина полиомавирусов» . Журнал клеточной физиологии . 231 (10): 2115–27. дои : 10.1002/jcp.25329 . ПМК 5217748 . ПМИД 26831433 .
- ^ «Веб-сайт таксономии ICTV» .
- ^ Международное агентство по исследованию рака (2013). «Рабочая группа МАИР по оценке канцерогенного риска для человека. Малярия и некоторые полиомавирусы (вирусы SV40, BK, JC и клетки Меркеля)» . Монографии МАИР по оценке канцерогенных рисков для человека . 104 .
- ^ Перес-Лосада М., Кристенсен Р.Г., Макклеллан Д.А., Адамс Б.Дж., Висиди Р.П., Демма Дж.К., Крэндалл К.А. (июнь 2006 г.). «Сравнение филогенетической кодивергенции между полиомавирусами и их хозяевами» . Журнал вирусологии . 80 (12): 5663–9. дои : 10.1128/JVI.00056-06 . ПМЦ 1472594 . ПМИД 16731904 .
- ^ Джон Р., Бак С.Б., Алландер Т., Этвуд В.Дж., Гарсиа Р.Л., Империале М.Дж., Майор Э.О., Рамквист Т., Норкин Л.К. (сентябрь 2011 г.). «Таксономические разработки семейства Polyomaviridae» . Архив вирусологии . 156 (9): 1627–34. дои : 10.1007/s00705-011-1008-x . ПМЦ 3815707 . ПМИД 21562881 .
- ^ Перейти обратно: а б «Таксономия вирусов: выпуск 2020 г.» . Международный комитет по таксономии вирусов (ICTV). Март 2021 года . Проверено 10 мая 2021 г.
- ^ Сикейра Дж.Д., Нг Т.Ф., Миллер М., Ли Л., Денг Икс, Додд Э., Батак Ф., Делварт Э. (июль 2017 г.). «Эндемическая инфекция выброшенных на берег южных каланов (Enhydra lutris nereis) новым парвовирусом, полиомавирусом и аденовирусом». Журнал болезней дикой природы . 53 (3): 532–542. дои : 10.7589/2016-04-082 . ПМИД 28192039 . S2CID 46740584 .
- ^ Дела Круз Ф.Н., Ли Л., Делварт Э., Песавент, Пенсильвания (2017). «Новый легочный полиомавирус у альпак» Ветеринарная микробиология . 201 : 49–55. дои : 10.1016/j.vetmic.2017.01.005 . ПМИД 28284622 .
- ^ Ци Д, Шан Т, Лю З, Дэн Х, Чжан З, Би В, Оуэнс-младший, Фэн Ф, Чжэн Л, Хуан Ф, Делварт Э, Хоу Р, Чжан В (2017). «Новый полиомавирус из носовой полости большой панды (Ailuropoda melanoleuca)» . Вирусологический журнал . 14 (1): 207. дои : 10.1186/s12985-017-0867-5 . ПМЦ 5658932 . ПМИД 29078783 .
- ^ Гонсалвес Мотта Майя Ф, Марсиэль де Соуза Ж, Сабино-Сантос Г, Хорхе Фумагалли М, Модха С, Рамиро Мурсия П, Тадеу Мораес Фигейреду Л (2018). «Новый полиомавирус у сигмодонтных грызунов из штата Сан-Паулу, Бразилия». Архив вирусологии . 163 (10): 2913–2915. дои : 10.1007/s00705-018-3913-8 . ПМИД 29931397 . S2CID 49351836 .
- ^ Лю П, Цю Ю, Син С, Чжоу Дж. Х., Ян В. Х., Ван Ц, Ли Ю., Хань Х, Чжан Ю. З., Гэ XY (2019). «Обнаружение и характеристика генома двух новых папилломавирусов и нового полиомавируса у землероек (Tupaia belangeri chinensis) в Китае» . Вирол Дж . 16 (1): 35. дои : 10.1186/s12985-019-1141-9 . ПМЦ 6423848 . ПМИД 30885224 .
- ^ Гейт Т., Дутта С., Оливер Дж., Робитайл А., Хампрас С., Комб Дж.Д., Маккей-Шопен С., Ле Кальвес-Кельм Ф., Фенске Н., Черпелис Б., Джулиано А.Р., Франчески С., Маккей Дж., Роллисон Д.Э., Томмасино М. ( 2017). «Выделение и характеристика нового предполагаемого полиомавируса человека» . Вирусология . 506 : 45–54. дои : 10.1016/j.virol.2017.03.007 . ПМЦ 9265179 . ПМИД 28342387 .
- ^ Альтман Л.К. (18 января 2008 г.). «Вирус связан с мощным раком кожи» . Нью-Йорк Таймс . Проверено 18 января 2008 г.
- ^ Шуда М, Арора Р, Квун Х.Дж., Фенг Х., Сарид Р., Фернандес-Фигерас М.Т., Толстов Ю., Джоеруп О., Мансухани М.М., Свердлов Ш., Чаудхари П.М., Кирквуд Дж.М., Налесник М.А., Кант Дж.А., Вайс Л.М., Мур П.С., Чанг И (сентябрь 2009 г.). «Инфекция полиомавирусом клеток Меркеля человека I. Экспрессия Т-антигена MCV в клеточной карциноме Меркеля, лимфоидных тканях и лимфоидных опухолях» . Международный журнал рака . 125 (6): 1243–9. дои : 10.1002/ijc.24510 . ПМК 6388400 . ПМИД 19499546 .
- ^ ван дер Мейден Э., Янссенс Р.В., Лаубер С., Баувес Бавинк Дж.Н., Горбаленя А.Е., Фельткамп М.К. (июль 2010 г.). «Обнаружение нового полиомавируса человека, связанного со спинулезной триходисплазией, у пациента с ослабленным иммунитетом» . ПЛОС Патогены . 6 (7): e1001024. дои : 10.1371/journal.ppat.1001024 . ПМЦ 2912394 . ПМИД 20686659 .
- ^ Казем С., ван дер Мейден Э., Фельткамп MC (август 2013 г.). «Полиомавирус, ассоциированный с триходисплазией,: вирусологическая основа и клиническое значение» . АПМИС . 121 (8): 770–82. дои : 10.1111/ап.12092 . ПМИД 23593936 . S2CID 13734654 .
- ^ Скуда Н., Хофманн Дж., Кальвиньяк-Спенсер С., Рупрехт К., Лиман П., Кюн Дж., Хенгель Х., Элерс Б. (май 2011 г.). «Новый полиомавирус человека, тесно связанный с лимфотропным полиомавирусом африканской зеленой обезьяны» . Журнал вирусологии . 85 (9): 4586–90. дои : 10.1128/jvi.02602-10 . ПМК 3126223 . ПМИД 21307194 .
- ^ Коруп С., Ритшер Дж., Кальвиньяк-Спенсер С., Труш Ф., Хофманн Дж., Моенс Ю., Зауэр И., Фойгт С., Шмук Р., Элерс Б. (2013). «Идентификация нового полиомавируса человека в органах желудочно-кишечного тракта» . ПЛОС ОДИН . 8 (3): e58021. Бибкод : 2013PLoSO...858021K . дои : 10.1371/journal.pone.0058021 . ПМЦ 3596337 . ПМИД 23516426 .
- ^ Мишра Н., Перейра М., Роудс Р.Х., Ан П., Пипас Дж.М. , Джайн К., Капур А., Бриз Т., Фауст П.Л., Липкин В.И. (ноябрь 2014 г.). «Идентификация нового полиомавируса у реципиента трансплантата поджелудочной железы со слепотой сетчатки и васкулитной миопатией» . Журнал инфекционных болезней . 210 (10): 1595–9. дои : 10.1093/infdis/jiu250 . ПМЦ 4334791 . ПМИД 24795478 .
- ^ Гарднер С.Д., Филд А.М., Коулман Д.В., Халм Б. (июнь 1971 г.). «Новый паповавирус человека (ВК), выделенный из мочи после трансплантации почки». Ланцет . 1 (7712): 1253–7. дои : 10.1016/s0140-6736(71)91776-4 . ПМИД 4104714 .
- ^ Пэджетт Б.Л., Уокер Д.Л., ЗуРейн Г.М., Эккроуд Р.Дж., Дессел Б.Х. (июнь 1971 г.). «Культивирование паповоподобного вируса из мозга человека с прогрессирующей мультифокальной лейкоэнцефалопатией». Ланцет . 1 (7712): 1257–60. дои : 10.1016/S0140-6736(71)91777-6 . ПМИД 4104715 .
- ^ Алландер Т., Андреассон К., Гупта С., Бьеркнер А., Богданович Г., Перссон М.А., Далианис Т., Рамквист Т., Андерссон Б. (апрель 2007 г.). «Идентификация третьего полиомавируса человека» . Журнал вирусологии . 81 (8): 4130–6. дои : 10.1128/JVI.00028-07 . ПМК 1866148 . ПМИД 17287263 .
- ^ Перейти обратно: а б Нгуен К.Д., Ли Э.Э., Юэ Ю, Сторк Дж., Пок Л., Норт Дж.П., Вандергрифф Т., Кокерелл С., Хослер Г.А., Пастрана Д.В., Бак С.Б., Ван Р.К. (май 2017 г.). «Полиомавирусы человека 6 и 7 связаны с зудящими и дискератозными дерматозами» . Журнал Американской академии дерматологии . 76 (5): 932–940.е3. дои : 10.1016/j.jaad.2016.11.035 . ПМЦ 5392424 . ПМИД 28040372 .
- ^ Хо Дж, Джедрик Дж. Дж., Фенг Х, Натали А. А., Грандинетти Л., Мирвиш Э., Креспо М. М., Ядав Д., Фасанелла К. Е., Прокселл С., Куан С. Ф., Пастрана Д. В., Бак CB, Шуда Ю., Мур П. С., Чанг Ю (май 2015 г.) ). «Зудящая сыпь, связанная с полиомавирусом человека 7, и виремия у реципиентов трансплантатов» . Журнал инфекционных болезней . 211 (10): 1560–5. дои : 10.1093/infdis/jiu524 . ПМЦ 4425822 . ПМИД 25231015 .
- ^ Топтан Т., Юсем С.А., Хо Дж., Мацусима Ю., Стабиле Л.П., Фернандес-Фигерас М.Т., Бхаргава Р., Ре А., Мур П.С., Чанг Ю. (февраль 2016 г.). «Обследование полиомавирусов человека при раке» . JCI-инсайт . 1 (2). doi : 10.1172/jci.insight.85562 . ПМЦ 4811373 . ПМИД 27034991 .
- ^ Сибрасс Э.А., Рейес А., Лим Э.С., Чжао Г., Мкакосия Р.С., Манари М.Дж., Гордон Дж.И., Ван Д. (октябрь 2012 г.). «Идентификация полиомавируса MW, нового полиомавируса в стуле человека» . Журнал вирусологии . 86 (19): 10321–6. дои : 10.1128/JVI.01210-12 . ПМЦ 3457274 . ПМИД 22740408 .
- ^ Бак CB, Фан GQ, Райджи М.Т., Мерфи П.М., Макдермотт Д.Х., Макбрайд А.А. (октябрь 2012 г.). «Полная последовательность генома десятого полиомавируса человека» . Журнал вирусологии . 86 (19): 10887. doi : 10.1128/JVI.01690-12 . ПМЦ 3457262 . ПМИД 22966183 .
- ^ Ю Г, Гренингер А.Л. , Иса П., Фан Т.Г., Мартинес М.А., де ла Лус Санчес М., Контрерас Х.Ф., Сантос-Пресиадо Дж.И., Парсоннет Дж. , Миллер С., ДеРизи Дж.Л., Делварт Э., Ариас К.Ф., Чиу С.И. (2012). «Обнаружение нового полиомавируса в образцах детей с острой диареей» . ПЛОС ОДИН . 7 (11): e49449. Бибкод : 2012PLoSO...749449Y . дои : 10.1371/journal.pone.0049449 . ПМК 3498111 . ПМИД 23166671 .
- ^ Лим Э.С., Рейес А., Антонио М., Саха Д., Икумапайи УН, Адейеми М., Стайн О.К., Скелтон Р., Бреннан Д.С., Мкакосия Р.С., Манари М.Дж., Гордон Дж.И., Ван Д. (февраль 2013 г.). «Открытие полиомавируса STL, полиомавируса наследственного рекомбинантного происхождения, который кодирует уникальный Т-антиген путем альтернативного сплайсинга» . Вирусология . 436 (2): 295–303. дои : 10.1016/j.virol.2012.12.005 . ПМЦ 3693558 . ПМИД 23276405 .
- ^ Гейт, Тарик; Дутта, Санкхадип; Оливер, Хавьер; Робитайл, Алексис; Хампрас, Шалака; Комб, Жан-Дамьен; Маккей-Шопен, Сандрин; Кальвес-Кельм, Флоренс Ле; Фенске, Нил (2017). «Выделение и характеристика нового предполагаемого полиомавируса человека» . Вирусология . 506 : 45–54. дои : 10.1016/j.virol.2017.03.007 . ПМЦ 9265179 . ПМИД 28342387 .
- ^ Прадо Х.К., Монези Т.А., Аморим А.Т., Лино В., Паладино А., Боккардо Э. (2018). «Полиомавирусы человека и рак: обзор» . Клиники (Сан-Паулу) . 73 (приложение 1): e558s. doi : 10.6061/clinics/2018/e558s . ПМК 6157077 . ПМИД 30328951 .
- ^ Далианис Т., Хирш Х.Х. (март 2013 г.). «Полиомавирусы человека в болезнях и раке». Вирусология . 437 (2): 63–72. дои : 10.1016/j.virol.2012.12.015 . ПМИД 23357733 .
- ^ Ван Гелу М., Хан М.Т., Элерс Б., Моенс У. (ноябрь 2012 г.). «Анализ генома новых полиомавирусов человека» . Обзоры по медицинской вирусологии . 22 (6): 354–77. дои : 10.1002/rmv.1711 . ПМИД 22461085 .
- ^ Эгли А., Инфанти Л., Дюмулен А., Бузер А., Самаридис Дж., Стеблер С., Госерт Р., Хирш Х.Х. (март 2009 г.). «Распространенность и репликация полиомавирусной инфекции BK и JC у 400 здоровых доноров крови» . Журнал инфекционных болезней . 199 (6): 837–46. дои : 10.1086/597126 . ПМИД 19434930 .
- ^ Перейти обратно: а б Пулен Д.Л., ДеКаприо Дж.А. (сентябрь 2006 г.). «Есть ли роль SV40 в развитии рака у человека?». Журнал клинической онкологии . 24 (26): 4356–65. дои : 10.1200/JCO.2005.03.7101 . ПМИД 16963733 .
- ^ Перейти обратно: а б цур Хаузен Х (декабрь 2003 г.). «SV40 при раке человека — бесконечная сказка?». Международный журнал рака . 107 (5): 687. doi : 10.1002/ijc.11517 . ПМИД 14566815 .
- ^ Газдар А.Ф., Бутель Дж.С., Карбоне М. (декабрь 2002 г.). «SV40 и опухоли человека: миф, ассоциация или причинно-следственная связь?». Обзоры природы. Рак . 2 (12): 957–64. дои : 10.1038/nrc947 . ПМИД 12459734 . S2CID 8878662 .
- ^ Кэрролл-Панхерст С., Энгельс Э.А., Стриклер Х.Д., Гедерт Дж.Дж., Вагнер Дж., Мортимер Э.А. (ноябрь 2001 г.). «Тридцатипятилетняя смертность после получения полиовакцины, зараженной SV40, в неонатальный период» . Британский журнал рака . 85 (9): 1295–7. дои : 10.1054/bjoc.2001.2065 . ПМЦ 2375249 . ПМИД 11720463 .
- ^ Шах КВ (январь 2007 г.). «SV40 и рак человека: обзор последних данных» . Международный журнал рака . 120 (2): 215–23. дои : 10.1002/ijc.22425 . ПМИД 17131333 . S2CID 20679358 .
- ^ Перейти обратно: а б Драхенберг CB, Хирш Х.Х., Рамос Э., Пападимитриу Х.К. (декабрь 2005 г.). «Полиомавирусная болезнь при трансплантации почки: обзор патологоанатомических данных и методов диагностики». Патология человека . 36 (12): 1245–55. дои : 10.1016/j.humpath.2005.08.009 . ПМИД 16311117 .
- ^ Висциди Р.П., Клейман Б. (2006). «Серологическая перекрестная реактивность между капсидами полиомавируса» . В Ахсане Н. (ред.). Полиомавирусы и болезни человека . Достижения экспериментальной медицины и биологии. Том. 577. стр. 73–84. дои : 10.1007/0-387-32957-9_5 . ISBN 978-0-387-29233-5 . ПМИД 16626028 .
- ^ Дрюс К., Башир Т., Деррис К. (январь 2000 г.). «Количественное определение полиомавируса человека JC в ткани головного мозга и спинномозговой жидкости пациентов с прогрессирующей мультифокальной лейкоэнцефалопатией методом конкурентной ПЦР». Журнал вирусологических методов . 84 (1): 23–36. дои : 10.1016/S0166-0934(99)00128-7 . ПМИД 10644084 .
- ^ Никелеит В., Хирш Х.Х., Бине И.Ф., Гудат Ф., Принс О., Далкен П., Тиль Г., Михатч М.Дж. (май 1999 г.). «Полиомавирусная инфекция реципиентов почечного аллотрансплантата: от латентной инфекции к манифестному заболеванию» . Журнал Американского общества нефрологов . 10 (5): 1080–9. дои : 10.1681/ASN.V1051080 . ПМИД 10232695 .
- ^ Рандхава П.С., Ватс А., Зигмунт Д., Свальски П., Скантлбери В., Шапиро Р., Финкельштейн С. (август 2002 г.). «Количественное определение вирусной ДНК в ткани почечного аллотрансплантата пациентов с вирусной нефропатией BK» . Трансплантация . 74 (4): 485–8. дои : 10.1097/00007890-200208270-00009 . ПМИД 12352906 . S2CID 30574884 .
- ^ Бусам К.Дж., Юнгблут А.А., Ректман Н., Койт Д., Пулитцер М., Бини Дж., Арора Р., Хэнсон Н.К., Тасселло Дж.А., Фросина Д., Мур П., Чанг Ю. (сентябрь 2009 г.). «Экспрессия полиомавируса клеток Меркеля в карциномах клеток Меркеля и ее отсутствие в комбинированных опухолях и нейроэндокринных карциномах легких» . Американский журнал хирургической патологии . 33 (9): 1378–85. дои : 10.1097/PAS.0b013e3181aa30a5 . ПМЦ 2932664 . ПМИД 19609205 .
- ^ Толстов Ю.Л., Пастрана Д.В., Фенг Х., Беккер Дж.К., Дженкинс Ф.Дж., Мошос С., Чанг Ю., Бак С.Б., Мур П.С. (сентябрь 2009 г.). «Инфекция полиомавирусом клеток Меркеля человека II. MCV является распространенной человеческой инфекцией, которую можно обнаружить с помощью иммуноанализа конформационного капсидного эпитопа» . Международный журнал рака . 125 (6): 1250–6. дои : 10.1002/ijc.24509 . ПМЦ 2747737 . ПМИД 19499548 .
- ^ Пастрана Д.В., Толстов Ю.Л., Беккер Дж.К., Мур П.С., Чанг Ю., Бак CB (сентябрь 2009 г.). «Количественная оценка серологической реакции человека на полиомавирус клеток Меркеля» . ПЛОС Патогены . 5 (9): e1000578. дои : 10.1371/journal.ppat.1000578 . ПМК 2734180 . ПМИД 19750217 .
- ^ Картер Дж.Дж., Полсон К.Г., Випф Г.К., Миранда Д., Мадлен М.М., Джонсон Л.Г., Лемос Б.Д., Ли С., Варкола А.Х., Айер Дж.Г., Нгием П., Галлоуэй Д.А. (ноябрь 2009 г.). «Ассоциация антител, специфичных к полиомавирусу клеток Меркеля, с карциномой клеток Меркеля» . Журнал Национального института рака . 101 (21): 1510–22. дои : 10.1093/jnci/djp332 . ПМЦ 2773184 . ПМИД 19776382 .
- ^ Элизабет Матису-Смит; К. Энн Хорсбург (2012). ДНК для археологов . Рутледж. ISBN 978-1598746815 .
- ^ Гросс Л. (июнь 1953 г.). «Фильтрующийся агент, выделенный из экстрактов лейкемии Ak, вызывающий рак слюнных желез у мышей C3H». Труды Общества экспериментальной биологии и медицины . 83 (2): 414–21. дои : 10.3181/00379727-83-20376 . ПМИД 13064287 . S2CID 34223353 .
- ^ Стюарт С.Е., Эдди Б.Е., Боргезе Н. (июнь 1958 г.). «Новообразования у мышей, инокулированных опухолевым агентом, содержащимся в культуре тканей». Журнал Национального института рака . 20 (6): 1223–43. дои : 10.1093/jnci/20.6.1223 . ПМИД 13549981 .
- ^ Эдди Б.Е., Стюарт С.Е. (ноябрь 1959 г.). «Характеристика вируса полиомы SE» . Американский журнал общественного здравоохранения и здоровья нации . 49 (11): 1486–92. дои : 10.2105/AJPH.49.11.1486 . ПМК 1373056 . ПМИД 13819251 .
- ^ Шовальтер Р.М., Пастрана Д.В., Бак CB (июль 2011 г.). «Гликозаминогликаны и сиалилированные гликаны последовательно облегчают проникновение инфекционного полиомавируса в клетки Меркеля» . ПЛОС Патогены . 7 (7): e1002161. дои : 10.1371/journal.ppat.1002161 . ПМК 3145800 . ПМИД 21829355 .
- ^ Готлиб К.А., Вильярреал LP (июнь 2001 г.). «Естественная биология среднего Т-антигена полиомавируса» . Обзоры микробиологии и молекулярной биологии . 65 (2): 288–318, вторая и третья страницы, оглавление. дои : 10.1128/ммбр.65.2.288-318.2001 . ПМК 99028 . ПМИД 11381103 .
- ^ Перетти А., Фитцджеральд ПК, Блисковский В., Пастрана Д.В., Бак CB (январь 2015 г.). «Геномная последовательность полиомавируса, ассоциированного с рыбой, полиомавируса 1 черноморского окуня (Centropristis striata)» . Геномные объявления . 3 (1): e01476-14. doi : 10.1128/genomeA.01476-14 . ПМЦ 4319505 . ПМИД 25635011 .
- ^ Лопес-Буэно А., Мавиан С., Лабелла А.М., Кастро Д., Боррего Дж.Дж., Алками А., Алехо А. (октябрь 2016 г.). «Сочетание иридовируса, полиомавируса и уникального члена новой группы папилломавирусов рыб у дорадоголового морского леща, пораженного лимфоцистисной болезнью» . Журнал вирусологии . 90 (19): 8768–79. дои : 10.1128/JVI.01369-16 . ПМК 5021401 . ПМИД 27440877 .
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|