Парамиксовирусиды
| Парамиксовирусиды | |
|---|---|
 | |
| Вирион и организация генома вируса чумы собак (CDV) | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Негарнавирикота |
| Сорт: | Монхивирицетес |
| Заказ: | Мононегавирусы |
| Семья: | Парамиксовирусиды |
| Подсемейства | |
Paramyxoviridae (от греческого para- «рядом» и myxa « слизь ») — семейство РНК-вирусов с отрицательной цепью отряда Mononegavirales . [1] [2] Позвоночные животные служат естественными хозяевами. [3] Заболевания, связанные с этим семейством, включают корь , эпидемический паротит и инфекции дыхательных путей . [4] В семействе четыре подсемейства, 17 родов, три из которых не отнесены к подсемейству, и 78 видов. [5]
Структура
[ редактировать ]Вирионы имеют оболочку, могут быть сферическими или плеоморфными и способны образовывать нитевидные вирионы. Диаметр около 150 нм. Геномы линейные, длиной около 15 КБ. [6] [1] Белки слияния и белки прикрепления выглядят как шипы на поверхности вириона. Белки матрикса внутри оболочки стабилизируют структуру вируса. Ядро нуклеокапсида состоит из геномной РНК, белков нуклеокапсида, фосфопротеинов и белков-полимераз.
Геном
[ редактировать ]
Геном представляет собой несегментированную РНК с отрицательным смыслом, длиной 15–19 тысяч оснований и содержит от шести до 10 генов. К экстрацистронным (некодирующим) регионам относятся:
- 3'-лидерная последовательность 50 нуклеотидов длиной , которая действует как промотор транскрипции .
- 5'-концевая последовательность длиной 50–161 нуклеотид.
- Межгеномные области между каждым геном имеют длину три нуклеотида для морбилливирусов, респировирусов и генипавирусов и переменную длину (один-56 нуклеотидов) для рубулавирусов.
Каждый ген содержит сигналы начала/остановки транскрипции в начале и конце, которые транскрибируются как часть гена.
Последовательность генов в геноме консервативна во всем семействе благодаря явлению, известному как полярность транскрипции (см. Mononegavirales ), при котором гены, ближайшие к 3'-концу генома, транскрибируются в большем количестве, чем гены, расположенные к 5'-концу. Это результат структуры генома. После транскрипции каждого гена РНК-зависимая РНК-полимераза останавливается, чтобы высвободить новую мРНК, когда она встречает межгенную последовательность. Когда работа РНК-полимеразы приостановлена, существует вероятность того, что она диссоциирует от генома РНК. Если он диссоциирует, он должен повторно войти в геном в лидерной последовательности, а не продолжать транскрибировать длину генома. В результате, чем дальше от лидерной последовательности расположены гены, расположенные ниже по ходу транскрипции, тем меньше они будут транскрибироваться РНК-полимеразой.
Доказательства существования модели с одним промотором были проверены, когда вирусы подвергались воздействию УФ-света. УФ-излучение может вызвать димеризацию РНК, что предотвращает транскрипцию РНК-полимеразой. Если вирусный геном соответствует модели с несколькими промоторами, уровень ингибирования транскрипции должен коррелировать с длиной гена РНК. Однако геном лучше всего описывался моделью с одним промотором. Когда геном парамиксовируса подвергался воздействию УФ-света, уровень ингибирования транскрипции был пропорционален расстоянию от лидерной последовательности. То есть, чем дальше ген от лидерной последовательности, тем больше вероятность того, что димеризация РНК ингибирует РНК-полимеразу.
Вирус использует модель с одним промотором, поскольку его гены расположены в относительном порядке белков, необходимых для успешной инфекции. Например, нуклеокапсидный белок N необходим в больших количествах, чем РНК-полимераза L.
Вирусы семейства Paramyxoviridae также антигенно стабильны, а это означает, что гликопротеины вирусов одинаковы у разных штаммов одного и того же типа. Выдвигаются две причины этого явления: первая заключается в том, что геном несегментирован и поэтому не может подвергаться генетической реассортации . Чтобы этот процесс произошел, необходимы сегменты, поскольку реассортация происходит, когда сегменты разных штаммов смешиваются вместе, чтобы создать новый штамм. При отсутствии сегментов ничто не может смешиваться друг с другом, поэтому антигенного сдвига не происходит. Вторая причина связана с идеей антигенного дрейфа . Поскольку РНК-зависимая РНК-полимераза не имеет функции проверки ошибок, при процессинге РНК происходит множество мутаций. Эти мутации накапливаются и в конечном итоге создаются новые штаммы. В соответствии с этой концепцией можно было бы ожидать, что парамиксовирусы не должны быть антигенно стабильными; однако, как представляется, верно обратное. Основная гипотеза антигенной стабильности вирусов заключается в том, что каждый белок и аминокислота выполняют важную функцию. Таким образом, любая мутация приведет к снижению или полной потере функции, что, в свою очередь, приведет к тому, что новый вирус станет менее эффективным. Эти вирусы не смогут выжить так долго, как более вирулентные штаммы, и поэтому вымрут.
Геномы многих парамиксовирусов подчиняются «правилу шести» . Общая длина генома почти всегда кратна шести. Вероятно, это связано с преимуществом того, что вся РНК связана с белком N (поскольку N связывает гексамеры РНК). Если РНК остается открытой, вирус не реплицируется эффективно. Последовательность гена:
- Нуклеокапсид – фосфопротеин – матрица – слияние – прикрепление – большой (полимераза)
Белки
[ редактировать ]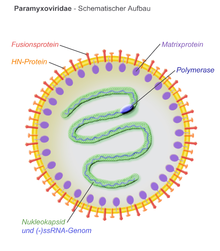
- N – белок нуклеокапсида связывается с геномной РНК (одна молекула на гексамер) и защищает РНК от расщепления нуклеазой.
- P – фосфопротеин связывается с белками N и L и входит в состав комплекса РНК-полимеразы. P — кофактор полимеразы.
- М – матричный белок собирается между оболочкой и ядром нуклеокапсида, организует и поддерживает структуру вириона.
- F – слитый белок выступает с поверхности оболочки в виде тримера и опосредует проникновение в клетку , вызывая слияние вирусной оболочки и клеточной мембраны путем слияния класса I. Одной из определяющих характеристик членов семейства Paramyxoviridae является потребность в нейтральном pH для фузогенной активности.
- H / HN / G – белки прикрепления клеток охватывают вирусную оболочку и выступают с поверхности в виде шипов. Они связываются с белками на поверхности клеток-мишеней, облегчая проникновение в клетку. Белки обозначены H ( гемагглютинин ) для морбилливирусов, поскольку они обладают гемагглютинационной активностью, наблюдаемой как способность вызывать слипание эритроцитов в лабораторных тестах. Белки прикрепления HN ( гемагглютинин-нейраминидаза ) встречаются у респировирусов, рубулавирусов и авулавирусов. Они обладают как гемагглютинационной, так и нейраминидазной активностью, которая расщепляет сиаловую кислоту на поверхности клетки, предотвращая повторное прикрепление вирусных частиц к ранее инфицированным клеткам. Прикрепляющие белки, не обладающие ни гемагглютинацией, ни нейраминидазной активностью, обозначаются G ( гликопротеинами ). Они встречаются у генипавирусов.
- L – большой белок является каталитической субъединицей РНК-зависимой РНК-полимеразы (RDRP).
- Акцессорные белки – механизм, известный как редактирование РНК (см. Mononegavirales ), позволяет производить несколько белков из гена P. Они не важны для репликации, но могут способствовать выживанию in vitro или могут участвовать в регуляции переключения с синтеза мРНК на синтез антигенома .
Жизненный цикл
[ редактировать ]
Репликация вируса цитоплазматическая . Проникновение в клетку-хозяина достигается путем прикрепления вируса к клетке-хозяину. Репликация и транскрипция соответствуют моделям вирусов с отрицательной цепью РНК . [7] Трансляция происходит путем «протекающего» сканирования, рибосомального шунтирования и терминации-повторной инициации РНК. Вирус покидает клетку-хозяина путем почкования. Естественными хозяевами служат позвоночные, включая человека и птиц. Путь передачи – частицы, передающиеся по воздуху. [1]
Paramyxoviridae ORF способны подвергаться редактированию мРНК, которое производит разные белки из одного и того же транскрипта мРНК путем смещения одного основания назад для считывания в другой открытой рамке считывания ( ) из-за присутствия вторичных структур, таких как псевдоузлы. Парамиксовирусы также подвергаются заиканию транскрипции, приводя к образованию поли (А) хвоста на конце транскриптов мРНК путем многократного перемещения назад по одному нуклеотиду за раз на конце матрицы РНК. [8] [9]
Таксономия
[ редактировать ]
Семейство: Paramyxoviridae. [5]
- Подсемейство: Avulavirinae , которое содержит три рода и 22 вида.
- Подсемейство: Metaparamyxovirinae , которое содержит один род и один вид.
- Подсемейство: Orthoparamyxovirinae , которое содержит восемь родов и 34 вида.
- Подсемейство: Rubulavirinae , которое содержит два рода и 18 видов.
- Неназначенные роды:
Патогенные парамиксовирусы
[ редактировать ]Ряд важных заболеваний человека вызывают парамиксовирусы. К ним относятся эпидемический паротит , а также корь , от которой в 2022 году умерло около 136 200 человек. [11]
Вирусы парагриппа человека (ВПЧ) являются второй по распространенности причиной заболеваний дыхательных путей у младенцев и детей. Существует четыре типа ВПЧ, известные как ВПЧ-1, ВПЧ-2, ВПЧ-3 и ВПЧ-4. HPIV-1 и HPIV-2 могут вызывать симптомы, похожие на простуду, а также круп у детей. HPIV-3 связан с бронхиолитом , бронхитом и пневмонией . ВПЧ-4 встречается реже, чем другие типы, и известно, что он вызывает легкие и тяжелые заболевания дыхательных путей. [12]
Парамиксовирусы также ответственны за ряд заболеваний у других видов животных, например , вирус чумы собак ( собаки ), вирус чумы чумы ( тюлени ), морбилливирус китообразных ( дельфины и морские свиньи ), вирус болезни Ньюкасла ( птицы ) и вирус чумы крупного рогатого скота ( крупный рогатый скот). ).
Некоторые парамиксовирусы, такие как генипавирусы , являются зоонозными патогенами, встречающимися в природе в организме животного-хозяина, но также способными инфицировать человека. Вирус Хендра и вирус Нипах рода Henipavirus появились у людей и домашнего скота в Австралии и Юго-Восточной Азии . Оба вируса заразны , высоковирулентны и способны заражать ряд видов млекопитающих и вызывать потенциально смертельные заболевания. Из-за отсутствия лицензированной человеческой вакцины (существует вакцина против вируса Хендра для лошадей) или противовирусной терапии вирус Хендра и вирус Нипах отнесены к уровня биобезопасности (BSL) 4 агентам . Геномная структура обоих вирусов соответствует типичному парамиксовирусу. [13]
Разнообразие и эволюция
[ редактировать ]Этот раздел необходимо обновить . ( март 2020 г. ) |
В последние несколько десятилетий, [ когда? ] парамиксовирусы были обнаружены у наземных, летучих и водных животных, что демонстрирует обширный круг хозяев и большое генетическое разнообразие вирусов. По мере развития молекулярных технологий и реализации программ вирусного надзора количество случаев открытия новых вирусов этой группы увеличивается. [4]
Эволюция парамиксовирусов до сих пор дискутируется. Используя пневмовирусы (мононегавирусное семейство Pneumoviridae ) в качестве внешней группы, парамиксовирусы можно разделить на две клады: одну, состоящую из авулавирусов и рубулавирусов , и другую, состоящую из респировирусов , генипавирусов и морбилливирусов . [14] Во второй кладе респировирусы представляют собой базальную группу. Клада респировирус-генипавирус-морбилливирус может быть базальной по отношению к кладе авулавирус-рубулавирус.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с «Вирусная зона» . ЭксПАСи . Проверено 15 июня 2015 г.
- ^ «Парамиксовирусы - Paramyxoviridae - РНК-вирусы с отрицательным смыслом - ICTV» . talk.ictvonline.org . Проверено 14 декабря 2020 г.
- ^ Филдс, Бернард Н .; Найп, Дэвид Махан; Хоули, Питер М., ред. (2013). Вирусология Филдса (6-е изд.). Филадельфия: Wolters Kluwer Health/Lippincott Williams & Wilkins. п. 883. ИСБН 9781451105636 . OCLC 825740706 .
- ^ Jump up to: а б Самал, С.К., изд. (2011). Биология парамиксовирусов . Кайстер Академик Пресс . ISBN 978-1-904455-85-1 .
- ^ Jump up to: а б «Таксономия вирусов: выпуск 2020 г.» . Международный комитет по таксономии вирусов (ICTV). Март 2021 года . Проверено 18 мая 2021 г.
- ^ Рима, Б; Балкема-Бушманн, А; Дандон, В.Г.; Дюпрекс, ВП ; Истон, А; Фушье, Р; Курат, Г; Лэмб, Р; Ли, Б; Рота, П; Ван, Л; Консорциум отчетов ICTV (декабрь 2019 г.). «Профиль таксономии вируса ICTV: Paramyxoviridae » . Журнал общей вирусологии . 100 (12): 1593–1594. дои : 10.1099/jgv.0.001328 . ПМЦ 7273325 . ПМИД 31609197 .
- ^ Фернс, Рэйчел; Племпер, Ричард К. (15 апреля 2017 г.). «Полимеразы парамиксовирусов и пневмовирусов» . Вирусные исследования . 234 : 87–102. doi : 10.1016/j.virusres.2017.01.008 . ISSN 0168-1702 . ПМЦ 5476513 . ПМИД 28104450 .
- ^ Хармон, Шон Б.; Мего, А. Джордж; Вертц, Гейл В. (январь 2001 г.). «Последовательности РНК, участвующие в терминации транскрипции респираторно-синцитиального вируса» . Журнал вирусологии . 75 (1): 36–44. doi : 10.1128/JVI.75.1.36-44.2001 . ISSN 0022-538X . ПМЦ 113895 . ПМИД 11119571 .
- ^ Жак, JP; Колакофски, Д. (1 мая 1991 г.). «Псевдошаблонная транскрипция в прокариотических и эукариотических организмах» . Гены и развитие . 5 (5): 707–713. дои : 10.1101/gad.5.5.707 . ISSN 0890-9369 . ПМИД 2026325 . S2CID 37461543 .
- ^ Марш Г.А., де Йонг С., Барр Дж.А., Тачеджян М., Смит С., Миддлтон Д., Ю М., Тодд С., Фурд Эй.Дж., Харинг В., Пейн Дж., Робинсон Р., Броз И., Крамери Дж., Филд Х.Э., Ван Л.Ф. (2012) ). «Вирус кедра: новый генипавирус, выделенный от австралийских летучих мышей» . ПЛОС Патогены . 8 (8): e1002836. дои : 10.1371/journal.ppat.1002836 . ПМК 3410871 . ПМИД 22879820 .
- ^ «Прогресс на пути к ликвидации кори – во всем мире, 2000–2022 гг.» . www.cdc.gov .
- ^ «CDC – HPIV – Обзор вирусов парагриппа человека» . www.cdc.gov . Проверено 19 сентября 2014 г.
- ^ Саватский (2008). «Вирус Хендра и Нипах» . Вирусы животных: молекулярная биология . Кайстер Академик Пресс. ISBN 978-1-904455-22-6 .
- ^ Маккарти А.Дж., Гудман С.Дж. (январь 2010 г.). «Переоценка противоречивых историй эволюции Paramyxoviridae и происхождения респировирусов с байесовской мультигенной филогенией». Заразить. Жене. Эвол . 10 (1): 97–107. Бибкод : 2010InfGE..10...97M . дои : 10.1016/j.meegid.2009.11.002 . ПМИД 19900582 .
Внешние ссылки
[ редактировать ]- ICTV Report: Paramyxoviridae
- Парамиксовирусы (1998) - морфология, геном, репликация, патогенез (требуется специальный доступ)
- «Вирус Хендра имеет растущее генеалогическое древо» . Пресс-релиз CSIRO по парамиксовирусу . 2001. Архивировано из оригинала 4 августа 2007 г.
- Вирусы животных
- геномы парамиксовирусов Ресурсный центр по вирусной биоинформатике,
- Вирусная зона : парамиксовирусы.
- База данных и ресурс анализа вирусных патогенов (ViPR): архив Paramyxoviridae 2022
| Базы данных органов управления : Национальные |
|---|