Вирус денге
| Вирус денге | |
|---|---|
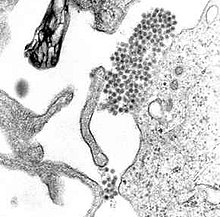 | |
| Микрофотография ПЭМ , показывающая вируса денге вирионы (скопление темных точек рядом с центром). | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Китриновирикота |
| Сорт: | Фласувирицеты |
| Заказ: | Амарилловирусы |
| Семья: | Флавивирусиды |
| Род: | Флавивирус |
| Разновидность: | Вирус денге |
Вирус денге ( DENV ) является причиной лихорадки денге . Это переносимый комарами одноцепочечный РНК- вирус семейства Flaviviridae ; Род флавивирусов . [1] [2] Было обнаружено четыре серотипа вируса, а пятый еще не подтвержден. [3] [4] [5] все это может вызвать полный спектр заболеваний. [1] Тем не менее, понимание вируса денге основным научным сообществом может быть упрощенным, поскольку, по- не отдельные антигенные группы, а континуум . видимому, существует [6] В этом же исследовании было выявлено 47 штаммов вируса денге . [7] Кроме того, коинфекция и отсутствие быстрых тестов на вирус Зика и чикунгунья усложняют ситуацию с реальными инфекциями. [8]
За последние 20 лет резко возросло распространение вируса денге , став одним из самых опасных человеческих патогенов, передающихся комарами, с которыми приходится иметь дело тропическим странам. По оценкам 2013 года, ежегодно происходит до 390 миллионов случаев инфицирования, и многие инфекции денге все чаще считаются бессимптомными или субклиническими. [9]
Эволюция
[ редактировать ]На основании анализа оболочечного белка как минимум три генотипа известны четвертый серотип . (от 1 до 3). В 2013 году был зарегистрирован [3] Единственное сообщение о пятом серотипе DEN-5 в 2015 г. [10] не воспроизводился и не сообщалось в дальнейшем. [5] По оценкам, скорость замены нуклеотидов для этого вируса составляет 6,5 × 10. −4 на нуклеотид в год, этот показатель аналогичен другим РНК-вирусам. По оценкам, американский африканский генотип развился между 1907 и 1949 годами. Этот период включает Первую и Вторую мировые войны , которые были связаны со значительным перемещением населения и нарушением окружающей среды, факторами, которые, как известно, способствуют эволюции новых трансмиссивных вирусов. разновидность . [ нужна ссылка ]
Байесовский анализ всех четырех серотипов показал, что их последний общий предок существовал около 340 г. н.э. (95% доверительный интервал: 280 г. до н.э. – 850 г. н.э.). [11]
Жизненный цикл
[ редактировать ]
Еще несколько сотен лет назад вирус денге передавался лесными циклами в Африке , Юго-Восточной Азии и Южной Азии между комарами рода Aedes и приматами , не являющимися людьми , с редкими случаями появления в человеческих популяциях. [12] [13] Однако глобальное распространение вируса денге последовало за его выходом из лесных циклов, и основной жизненный цикл теперь включает исключительно передачу между людьми и Aedes . комарами [14] Вертикальная передача инфекции от комара к комару также наблюдалась у некоторых видов переносчиков. [15] Было обнаружено, что собаки инфицированы вирусом, но необходимы дополнительные исследования, чтобы определить, могут ли собаки или другие животные служить резервуарами или просто случайными хозяевами. [16]
Недавние результаты показывают, что по мере заражения вирусом клеток человека гомеостатические процессы хозяина, такие как аутофагия и стрессовая реакция ЭР , не говоря уже об апоптозе. в зависимости от типа инфицированных клеток запускаются [17] Активация аутофагии и стресса ЭР во время инфекции усиливает размножение вируса. [18] [19] Попытки предоставить подробные описания жизненного цикла лихорадки денге на клеточном уровне публикуются в обзорных статьях различных исследовательских групп. [20] [21]
Геном
[ редактировать ]DENV Геном состоит примерно из 11 000 оснований одноцепочечной РНК с положительным смыслом (оцРНК), которая кодирует три структурных белка ( капсидный белок C, мембранный белок M, оболочечный белок E) и семь неструктурных белков (NS1, NS2a, NS2b, NS3). , NS4a, NS4b, NS5). [4] Он также включает короткие некодирующие области как на 5'-, так и на 3'- концах. [1] [22]
Структурные и ферментативные белки
[ редактировать ]
белок Е
[ редактировать ]Белок DENV E (оболочка), обнаруженный в виде димера на поверхности зрелой вирусной частицы, важен для первоначального прикрепления этой частицы к клетке-хозяину. Каждый мономер белка E состоит из трех эктодоменов, от ED1 до ED3, и трансмембранного сегмента. ED2 включает интерфейс димеризации, два сайта гликозилирования и пептид слияния с клеточной мембраной. ED3 представляет собой непрерывный полипептидный сегмент; его складка компактна и подобна иммуноглобулину. [23] [24] Вирус денге передается видами комаров рода Aedes . Несколько молекул, которые взаимодействуют с вирусным белком Е (ноинтегрин, захватывающий ICAM3, [25] CD209, [26] Раб 5, [27] ГРП 78, [28] и рецептор маннозы [29] ) было показано, что они являются важными факторами, опосредующими прикрепление и проникновение вируса. [24] мембранная форма рибосомального белка SA . В прикреплении может участвовать и [30] Известно, что белок Е содержит физико-химически консервативные эпитопы В-клеток и Т-клеток, которые можно использовать для разработки вакцины. [31] Рекомбинантные домены белка Е используются в качестве четко определенных антигенов при серологическом обнаружении антител, направленных против вируса денге , а также в качестве иммуногенов в вакцинах-кандидатах. [32] [33] [34]
белок prM/M
[ редактировать ]Белок prM (мембранный) DENV, который играет важную роль в формировании и созревании вирусной частицы, состоит из семи антипараллельных β-цепей, стабилизированных тремя дисульфидными связями. [24]
Гликопротеиновая оболочка зрелого вириона DENV состоит из 180 копий белков Е и М. Незрелый вирион начинается с белков E и prM, образующих 90 гетеродимеров, которые придают вирусной частице остроконечный внешний вид. Эта незрелая вирусная частица отпочковывается в эндоплазматическом ретикулуме и в конечном итоге по секреторному пути попадает в аппарат Гольджи. Когда вирион проходит через сеть транс-Гольджи, он подвергается воздействию низкого pH. Эта кислая среда вызывает конформационные изменения белка E, которые отделяют его от белка prM и заставляют образовывать гомодимеры E, которые плотно прилегают к поверхности вируса, придавая созревающему вириону гладкий вид. Во время этого созревания пептид pr отщепляется от пептида М протеазой хозяина, фурином . Белок М затем действует как трансмембранный белок под оболочкой E-белка зрелого вириона. Пептид pr остается связанным с белком Е до тех пор, пока вирусная частица не выйдет во внеклеточную среду. Этот pr-пептид действует как колпачок, закрывая гидрофобную петлю слияния белка Е до тех пор, пока вирусная частица не выйдет из клетки. [24]
белок NS3
[ редактировать ]DENV NS3 представляет собой сериновую протеазу, а также РНК-хеликазу и РТФазу/НТФазу. Протеазный домен состоит из шести β-нитей, организованных в два β-цилиндра, образованных остатками 1–180 белка. Каталитическая триада (His-51, Asp-75 и Ser-135) находится между этими двумя β-цилиндрами, и активность зависит от присутствия сегмента из 43 аминокислот кофактора NS2B. Этот кофактор окружает протеазный домен NS3 и становится частью активного сайта. Остальные остатки NS2B до и после области кофактора содержат спиральные домены, участвующие в мембраносвязывании. Остальные остатки NS3 (180–618) образуют три субдомена хеликазы DENV. Шестицепочечный параллельный β-лист, окруженный четырьмя α-спиралями, образует субдомены I и II, а субдомен III состоит из четырех α-спиралей, окруженных тремя более короткими α-спиралями и двумя антипараллельными β-нитями. [24]
белок NS4A
[ редактировать ]DENV NS4A — неструктурный белок, участвующий в изменении кривизны клеточной мембраны. [35] и индукция аутофагии. [19] Помимо способности изменять мембрану, NS4A является каркасом для комплекса репликации вируса и подвергается олигомеризации. [36] Мутации NS4A, влияющие на взаимодействие с NS4B, отменяют или значительно снижают репликацию вируса, что указывает на важность NS4A и его взаимодействия с NS4B в размножении денге. [37]
белок NS5
[ редактировать ]Белок DENV NS5 представляет собой пептид из 900 остатков с метилтрансферазным доменом на N-конце (остатки 1–296) и РНК-зависимой РНК-полимеразой (RdRp) на С-конце (остатки 320–900). Домен метилтрансферазы состоит из сэндвича α/β/β, фланкированного N- и C-концевыми субдоменами. DENV RdRp похож на другие RdRps, содержащие субдомены ладони, пальца и большого пальца, а также мотив GDD для включения нуклеотидов. [24]
Комплексы между белком Е и нейтрализующими антителами
[ редактировать ]Кристаллические структуры комплексов антител либо с эктодоменом (sE) вирусного белка Е, либо с его доменом 3 (ED3) помогли понять молекулярные основы распознавания и нейтрализации вируса. Некоторые из эпитопов частично или полностью недоступны в известной структуре зрелого вириона. Таким образом, предполагается, что соответствующие антитела связываются с альтернативными или переходными конформациями вируса при 37 °C. [ нужна ссылка ]
- Мышиное антитело E111 нейтрализует DENV1. Его Fab- и scFv-фрагменты кристаллизовались в комплексе с доменом ED3 DENV1. Его эпитоп расположен вокруг β-цепей C и C' ED3 и промежуточной петли. [38]
- Мышиное антитело 1A1D-2 сильно нейтрализует DENV1, DENV2 и DENV3. Его Fab-фрагмент кристаллизовался в комплексе с доменом ED3 DENV2. Его эпитоп охватывает β-цепи A и G ED3. [39]
- Мышиное антитело 2H12 перекрестно реагирует со всеми четырьмя серотипами DENV. Нейтрализует соответствующие вирусы, кроме DENV2. Его Fab-фрагмент кристаллизовался в комплексе с доменами ED3 DENV1, DENV3 и DENV4. Его эпитоп расположен вокруг консервативной петли AB ED3. [40]
- Мышиное антитело 4E11 нейтрализует все четыре серотипа DENV с различной эффективностью. Его фрагмент scFv был кристаллизован в комплексе с доменами ED3 четырех серотипов DENV. Его эпитоп охватывает β-цепи A и G ED3, как и эпитоп 1A1D-2. [41] [42] Структуры с разрешением 2,0 Å позволили проанализировать роль молекул воды внутри белковых интерфейсов и роль соматических гипермутаций за пределами этих интерфейсов во взаимодействиях и перекрестном узнавании. [43]
- Антитело шимпанзе 5H2 эффективно нейтрализует DENV4. Его Fab-фрагмент кристаллизовался в комплексе с белком sE DENV4. Его эпитоп включен в домен 1 (ED1) белка Е. [44]
- Человеческие антитела Ede1-C10, Ede2-A11 и Ede2-B7 эффективно нейтрализуют все четыре серотипа DENV. Их Fab- или scFv-фрагменты кристаллизовались в комплексе с белком sE DENV2. Детерминанты узнавания этих антител находятся в серотип-инвариантном сайте на границе раздела димера E и включают открытые боковые цепи петли слияния E и две консервативные боковые цепи гликанов. [45]
Болезнь
[ редактировать ]Общие названия лихорадки денге включают лихорадку перелома костей , рвоту и денди лихорадку ; К тяжелым формам относятся геморрагическая лихорадка денге и шоковый синдром денге. [46] Денге встречается в тропическом и субтропическом климате по всему миру, в основном в городских и пригородных районах. [47] Люди всех возрастов, контактирующие с инфицированными комарами, подвергаются риску развития лихорадки денге. Заболевание чаще всего возникает в сезон дождей в тропических странах Юго-Восточной Азии , Южной Азии и Южной Америки , где наблюдается большое количество инфицированных комаров. [48] Вирус передается человеку через укусы инфицированных самок комаров, хотя люди не способны передавать болезнь и не заразны. [47] [49] [48] Инкубационный период составляет от 3 до 14 дней, а период болезни — 3–7 дней. [49] [50] Признаки и симптомы могут включать сильную головную боль; ретроорбитальная боль; боль в мышцах, суставах и костях; макулярная или макулопапулезная сыпь; и незначительные геморрагические проявления, включая петехии, экхимозы, пурпуру, носовое кровотечение, кровоточивость десен, гематурию или положительный результат теста на жгут. [51] Недавний систематический обзор и метаанализ показали, что аллергические симптомы являются одними из основных симптомов, которые тесно связаны с тяжестью лихорадки денге. [52]
Механизм заражения
[ редактировать ]- Белок оболочки E вируса денге (DENV) связывается с клеточным рецептором. Точная природа клеточного рецептора полностью не выяснена.
- DENV подвергается эндоцитозу. Подкисление эндосомы приводит к конформационному изменению Е, обнажая последовательность слитого пептида, которая облегчает слияние оболочки с мембраной эндосомы, высвобождая капсид вириона в цитоплазму.
- Раскрытие оболочки происходит в цитоплазме.
- Механизм трансляции хозяина (рибосомы) переводит (+) оссРНК в один полипептид.
- Клеточные и вирусные протеиназы расщепляют полипептид на 10 белков (E, M, C и 7 неструктурных/ферментативных белков) при внедрении на мембрану ЭР.
- Как только синтезируется функциональная РНК-зависимая РНК-полимераза, может начаться репликация РНК. Синтез асимметричен: положительной цепи в 10 раз больше, чем отрицательной.
- Сборка происходит на внутриклеточных мембранах, которые отпочковываются в ЭР (образуя оболочку из мембраны ЭР). Последующее отпочкование из ЭР через Гольджи в везикулы обеспечивает созревание посредством посттрансляционных модификаций, например гликозилирования и трансформационных перестроек pH.
- Выход происходит путем экзоцитоза. [53]
Тяжелое заболевание
[ редактировать ]У некоторых людей развиваются более тяжелые формы денге, такие как геморрагическая лихорадка денге. Различные штаммы вирусов, взаимодействующие с людьми с разным иммунным фоном, приводят к сложному взаимодействию. Среди возможных причин — перекрестный серотипический иммунный ответ, реализуемый по механизму, известному как антителозависимое усиление , которое происходит, когда человек, ранее инфицированный лихорадкой денге, заражается во второй, третий или четвертый раз. Предыдущие антитела к старому штамму вируса денге теперь мешают иммунному ответу на нынешний штамм, что парадоксальным образом приводит к большему проникновению и поглощению вируса. [54]
Взаимодействие с иммунной системой
[ редактировать ]В последние годы многие исследования показали, что флавивирусы, особенно вирус денге , обладают способностью подавлять врожденный иммунный ответ во время инфекции. [55] [56] Действительно, вирус денге имеет множество неструктурных белков, которые позволяют ингибировать различные медиаторы реакции врожденной иммунной системы. [ нужна ссылка ] Эти белки действуют на двух уровнях:
Подавление передачи сигналов интерферона путем блокирования преобразователя сигнала
[ редактировать ]NS4B представляет собой небольшой гидрофобный белок, расположенный в ассоциации с эндоплазматической сетью. Он может блокировать фосфорилирование STAT 1 после индукции интерферонами типа I альфа и бета. Фактически, поскольку активность киназы Tyk2 снижается в связи с вирусом денге , снижается и фосфорилирование STAT 1. [57] Более того, реакция врожденной иммунной системы на вирус дополнительно ослабляется, поскольку экспрессия гена(ов), стимулирующего интерферон (ISG), ограничивается вышеупомянутым белком NS4B. Кофакторы NS2A и NS4A также могут принимать участие в ингибировании STAT 1. [58]
NS5 - присутствие этого белка массой 105 кДа приводит к инактивации STAT2 (посредством передачи сигнала ответа на интерферон), когда он экспрессируется отдельно. [59] Когда NS5 расщепляется протеазой (NS2B3) с NS4B, он может разрушать STAT2. Фактически, после расщепления NS5 протеазой происходит ассоциация лигазы E3 со STAT2, и лигаза E3 нацеливается на STAT2 для деградации. [60] [61]
Подавление реакции интерферона I типа
[ редактировать ]Протеазный комплекс NS2B3-b представляет собой протеолитическое ядро, состоящее из последних 40 аминокислот NS2B и первых 180 аминокислот NS3.Расщепление предшественника NS2B3 активирует протеазный комплекс. [62]
Этот протеазный комплекс позволяет ингибировать выработку интерферона I типа за счет снижения активности промотора IFN-бета; Протеазный комплекс NS2B3 участвует в ингибировании фосфорилирования IRF3. [63] Протеазный комплекс NS2B3 ингибирует (расщепляя) белок MITA, который обеспечивает активацию IRF3. [64]
Белок слюны комара D7
[ редактировать ]Вирус денге передается видом комаров Aedes aegypti , который производит слюну, содержащую более 100 уникальных белков, включая семейство белков D7. [65] Раньше ученые считали, что слюна A. aegypti при передаче фактически усиливает вирус денге в организме. Считалось, что слюна комара ускоряет распространение вируса из-за ослабления иммунного ответа хозяина. Однако недавнее исследование показало, что белок D7 препятствует передаче вируса в клетки-хозяева. [65]
Иммунные реакции антител, которые пытаются бороться с чужеродным вирусом, на самом деле увеличивают передачу и усугубляют инфекцию. Уровни белка D7 более распространены в слюнных железах комаров, инфицированных денге, по сравнению с неинфицированными. [65] D7 содержится в слюне комаров и считается, что он способствует процессу кровоснабжения. Несмотря на предыдущие предположения, D7 может модулировать клетку-хозяина и действовать против вируса, предотвращая вирусную инфекцию. [65] К сожалению, белки D7 провоцируют иммунные реакции, которые повышают уровень антител против D7. Эти антитела ингибируют функцию белков D7, которые усиливают передачу вируса денге. [ нужна ссылка ] . Хотя иммунные ответы против белков D7 могут ухудшить их противовирусную активность, исследование показало, что у субъектов, не страдающих DENV, уровни анти-D7 IgG немного выше, чем у инфицированных, хотя это не было статистически значимым. [66] Таким образом, необходимы дополнительные исследования семейства белков D7, чтобы выяснить его роль в инфекции DENV и его применимость в медицине.
Исследования вакцин
[ редактировать ]Два типа вакцины против денге были одобрены и коммерчески доступны. [67] [68] 5 декабря 2022 года Европейское агентство по лекарственным средствам одобрило Qdenga, живую четырехвалентную аттенуированную вакцину для взрослых, подростков и детей от четырех лет. [68] Вакцина Денгваксия 2016 года рекомендуется только лицам, которые ранее были инфицированы, или группам населения с высоким уровнем предшествующего инфицирования к девяти годам. [69] [47] Денгваксия одобрена в 11 странах (Мексика, Филиппины, Индонезия, Бразилия, Сальвадор, Коста-Рика, Парагвай, Гватемала, Перу, Таиланд и Сингапур). [70] [67] [71]
Несколько вакцин разрабатываются частными и государственными исследователями. [72] Разработка вакцины против этой болезни является сложной задачей. Поскольку заболевание могут вызывать четыре различных серотипа вируса, вакцина должна быть иммунизирована против всех четырех типов, чтобы быть эффективной. [3] Вакцинация только против одного серотипа может привести к тяжелому геморрагическому шоку при денге при заражении другим серотипом из-за антителозависимого усиления. При заражении вирусом денге иммунная система вырабатывает перекрестно-реактивные антитела, которые обеспечивают иммунитет к этому конкретному серотипу. Однако эти антитела не способны нейтрализовать другие серотипы при повторном заражении и фактически усиливают репликацию вируса . Когда макрофаги поглощают «нейтрализованный» вирус, вирус способен реплицироваться внутри макрофага, вызывая заболевание. Эти перекрестно реагирующие неэффективные антитела облегчают доступ вируса к макрофагам, что вызывает более тяжелое заболевание (геморрагическую лихорадку денге, шоковый синдром денге). Распространенной проблемой, с которой сталкиваются в регионах, эндемичных по денге, является заражение матерей денге; после рождения потомство несет иммунитет от своей матери и восприимчиво к геморрагической лихорадке при заражении любым из трех других серотипов. [73] В 2012 году одна вакцина находилась на стадии III испытаний, и началось планирование использования вакцины и надзор за ее эффективностью. [74]
В 2009 году Санофи-Пастер приступила к строительству нового предприятия в Невиль-сюр-Сон ( фр . ) , пригороде Лиона (Франция). Это подразделение производит вакцину из четырех серотипов для испытаний III фазы. В сентябре 2014 года генеральный директор компании Sanofi-Pasteur представил предварительные результаты исследования эффективности III фазы исследования в Латинской Америке. Эффективность каждого серотипа (ST) широко варьировала: 42,3% для ST2, 50,3% для ST1, 74,0% для ST3 и 77,7% для ST4. Полный анализ данных III фазы латиноамериканско-карибского исследования будет рассмотрен внешними экспертами перед публикацией в рецензируемом научном журнале. Первичные результаты должны быть представлены на ежегодном собрании Американского общества тропической медицины и гигиены , которое состоится 2–6 ноября 2014 г. в Новом Орлеане. [75]
В сентябре 2012 года сообщалось, что одна из вакцин не показала хороших результатов в клинических испытаниях. [3]
В конце 2015 — начале 2016 года первая вакцина против лихорадки денге, Dengvaxia (CYD-TDV) производства Sanofi-Pasteur, была зарегистрирована в нескольких странах для применения у лиц в возрасте 9–45 лет, проживающих в эндемичных районах. [ нужна ссылка ]
1 мая 2019 года Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США объявило об одобрении Dengvaxia, первой вакцины для профилактики болезни денге, вызываемой всеми серотипами вируса денге, у людей в возрасте от 9 до 16 лет, у которых ранее была лабораторно подтверждена инфекция денге и которые живут в эндемичных районах. Денге является эндемичным заболеванием на территориях Американского Самоа, Гуама, Пуэрто-Рико и Виргинских островов США. [76]
Исследования лекарств
[ редактировать ]Утвержденных прямых противовирусных методов лечения лихорадки Денге не существует. Большинство исследований противовирусных препаратов для лечения инфекций денге были сосредоточены на ингибировании протеазы NS2B/NS3 или белков NS5. Сообщаемые подходы к ингибиторам протеазы сосредоточены в основном на таргетных ковалентных ингибиторах . [77] [78] Один препарат, Балапиравир , перепрофилированный ингибитор полимеразы NS5 гепатита С, прошел фазу II клинических испытаний, но был остановлен из-за отсутствия эффективности. [79] [80]
Хотя не существует утвержденных прямых противовирусных методов лечения лихорадки денге, большинство противовирусных исследований сосредоточено на ингибировании протеазы NS2B/NS3 или белков NS5. Сообщаемые подходы к ингибиторам протеаз в основном нацелены на ковалентные ингибиторы. Один препарат, Балапиравир, первоначально перепрофилированный как ингибитор полимеразы NS5 гепатита С, достиг фазы II клинических испытаний, но был остановлен из-за отсутствия эффективности. [81]
Журнал исследует новый подход, изучая потенциал комплексов металл-ТПИ (с железом, кобальтом и цинком) в качестве ингибиторов репликации вируса денге. Zn-TPI и Fe-TPI продемонстрировали низкую токсичность и значительное ингибирование DENV-3, тогда как Co-TPI продемонстрировал противовирусную активность в низких концентрациях. Эти результаты открывают новые возможности для разработки более эффективных противовирусных методов лечения лихорадки денге. [82]
Ссылки
[ редактировать ]- ^ Jump up to: а б с Роденхейс-Зиберт И.А., Вильшут Дж., Смит Дж.М. (август 2010 г.). «Жизненный цикл вируса денге: вирусные факторы и факторы хозяина, модулирующие инфекционность» . Клеточные и молекулярные науки о жизни . 67 (16): 2773–86. дои : 10.1007/s00018-010-0357-z . ПМЦ 11115823 . ПМИД 20372965 . S2CID 4232236 .
- ^ ВОЗ (2009). Рекомендации по диагностике, лечению, профилактике и контролю лихорадки Денге (PDF) . Всемирная организация здравоохранения. ISBN 978-92-4-154787-1 .
- ^ Jump up to: а б с д Нормил Д. (октябрь 2013 г.). «Тропическая медицина. Удивительный новый вирус денге подрывает усилия по борьбе с болезнями». Наука . 342 (6157): 415. Бибкод : 2013Sci...342..415N . дои : 10.1126/science.342.6157.415 . ПМИД 24159024 .
- ^ Jump up to: а б Двиведи В.Д., Трипати И.П., Трипати Р.К., Бхарадвадж С. и Мишра С.К. (2017). Геномика, протеомика и эволюция вируса Денге . Брифинги по функциональной геномике.16(4): 217–227, https://doi.org/10.1093/bfgp/elw040.
- ^ Jump up to: а б Тейлор-Робинсон, Эндрю В. (3 марта 2016 г.). «Предполагаемый пятый серотип лихорадки денге – потенциальные последствия для диагностики, терапии и разработки вакцин» . Международный журнал клинической и медицинской микробиологии . 2016 . дои : 10.15344/2456-4028/2016/101 . ISSN 2456-4028 .
- ^ «Новое понимание вируса Денге » . ScienceDaily . 18 сентября 2015 года . Проверено 7 мая 2016 г.
- ^ «Вторичное заражение денге может быть опасным для жизни, говорят исследования» . НДТВ . Великобритания. 18 сентября 2015 года . Проверено 7 мая 2016 г.
- ^ Фогельс CB, Рюкерт С., Кавани С.М., Перкинс Т.А., Эбель Г.Д., Грубау Н.Д. (январь 2019 г.). «Коинфекция и ко-трансмиссия арбовируса: игнорируемая проблема общественного здравоохранения?» . ПЛОС Биология . 17 (1): e3000130. дои : 10.1371/journal.pbio.3000130 . ПМК 6358106 . PMID 30668574 .
- ^ Бхатт С., Гетинг П.В., Брэйди О.Дж., Мессина Дж.П., Фарлоу А.В., Мойес К.Л., Дрейк Дж.М., Браунштейн Дж.С., Хоэн А.Г., Санко О., Майерс М.Ф., Джордж Д.Б., Джениш Т., Винт Г.Р., Симмонс С.П., Скотт Т.В., Фаррар Дж.Дж. , Хэй С.И. (апрель 2013 г.). «Глобальное распространение и бремя денге» . Природа . 496 (7446): 504–7. Бибкод : 2013Natur.496..504B . дои : 10.1038/nature12060 . ПМК 3651993 . ПМИД 23563266 .
- ^ Мустафа, М.С.; Расотги, В.; Джайн, С.; Гупта, В. (2015). «Открытие пятого серотипа вируса денге (DENV-5): новая дилемма общественного здравоохранения в борьбе с денге» . Медицинский журнал Вооруженных Сил Индии . 71 (1): 67–70. дои : 10.1016/j.mjafi.2014.09.011 . ISSN 0377-1237 . ПМК 4297835 . ПМИД 25609867 .
- ^ Коста Р.Л., Волоч С.М., Шраго К.Г. (2012)Сравнительная эволюционная эпидемиология серотипов вируса денге. Заразить Генет Эвол 12(2):309-314
- ^ «Вирус Денге» . Информация о патогене (PathInfo) . Институт биоинформатики Вирджинии, Технологический институт Вирджинии. Архивировано из оригинала 30 августа 2010 г.
- ^ Холмс ЕС, Твидди СС (май 2003 г.). «Происхождение, появление и эволюционная генетика вируса денге». Инфекция, генетика и эволюция . 3 (1): 19–28. дои : 10.1016/s1567-1348(03)00004-2 . ПМИД 12797969 .
- ^ Холстед С.Б. (январь 1988 г.). «Патогенез лихорадки денге: проблемы молекулярной биологии». Наука . 239 (4839): 476–81. Бибкод : 1988Sci...239..476H . дои : 10.1126/science.3277268 . ПМИД 3277268 .
- ^ Хаддоу А.Д., Гузман Х., Попов В.Л., Вуд Т.Г., Виден С.Г., Хаддоу А.Д., Теш Р.Б., Уивер С.С. (июнь 2013 г.). «Первое выделение флавивируса Aedes в Западном полушарии и свидетельства вертикальной передачи комара Aedes (Stegomyia) albopictus (Diptera: Culicidae)» . Вирусология . 440 (2): 134–9. дои : 10.1016/j.virol.2012.12.008 . ПМИД 23582303 .
- ^ Тонгюань С., Киттаяпонг П. (30 августа 2017 г.). «Первые доказательства заражения денге у домашних собак, живущих в различных экологических условиях Таиланда» . ПЛОС ОДИН . 12 (8): e0180013. Бибкод : 2017PLoSO..1280013T . дои : 10.1371/journal.pone.0180013 . ПМЦ 5576688 . ПМИД 28854207 .
- ^ Гош Рой С., Садиг Б., Датан Э., Локшин Р.А., Закери З. (май 2014 г.). «Регуляция выживания и гибели клеток при флавивирусных инфекциях» . Всемирный журнал биологической химии . 5 (2): 93–105. doi : 10.4331/wjbc.v5.i2.93 (неактивен 31 января 2024 г.). ПМК 4050121 . ПМИД 24921001 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Датан Э., Рой С.Г., Жермен Г., Зали Н., Маклин Дж.Э., Гольшан Г., Харбаджан С., Локшин Р.А., Закери З. (март 2016 г.). «Аутофагия, вызванная денге, репликация вируса и защита от гибели клеток требуют активации пути стресса ER (PERK)» . Смерть клеток и болезни . 7 (е2127): е2127. дои : 10.1038/cddis.2015.409 . ПМЦ 4823927 . ПМИД 26938301 .
- ^ Jump up to: а б Маклин Дж. Э., Вудзинска А., Датан Э., Кваглино Д., Закери З. (июнь 2011 г.). «Аутофагия, индуцированная флавивирусом NS4A, защищает клетки от гибели и усиливает репликацию вируса» . Журнал биологической химии . 286 (25): 22147–59. дои : 10.1074/jbc.M110.192500 . ПМЦ 3121359 . ПМИД 21511946 .
- ^ Закери З., Гош-Рой С., Датан Э., Маклин Дж.Э., Занди К., Жермен Дж. (2015). «Клеточная смерть и вирусная инфекция – краткий обзор». 20 лет клеточной смерти . 1 (1): 292–325. дои : 10.13140/RG.2.2.35359.97447 .
- ^ Нойфельдт К.Дж., Кортезе М., Акоста Э.Г., Бартеншлагер Р. (февраль 2018 г.). «Перемонтирование сотовых сетей членами семейства Flaviviridae» . Обзоры природы. Микробиология . 16 (3): 125–142. дои : 10.1038/nrmicro.2017.170 . ПМК 7097628 . ПМИД 29430005 .
- ^ Хэнли К.А., Weaver SC, ред. (2010). Границы в исследованиях вируса денге . Кайстер Академик. п. 13. ISBN 978-1-904455-50-9 . Проверено 22 сентября 2020 г.
- ^ Модис Ю., Огата С., Клементс Д., Харрисон С.С. (июнь 2003 г.). «Лиганд-связывающий карман в гликопротеине оболочки вируса денге» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (12): 6986–91. Бибкод : 2003PNAS..100.6986M . дои : 10.1073/pnas.0832193100 . ПМК 165817 . ПМИД 12759475 .
- ^ Jump up to: а б с д и ж Перера Р., Кун Р.Дж. (август 2008 г.). «Структурная протеомика вируса денге» . Современное мнение в микробиологии . 11 (4): 369–77. дои : 10.1016/j.mib.2008.06.004 . ПМК 2581888 . ПМИД 18644250 .
- ^ Тассанитритхеп Б., Берджесс Т.Х., Гранелли-Пиперно А., Трампфеллер С., Финке Дж., Сан В., Эллер М.А., Паттанапаньясат К., Сарасомбат С., Биркс Д.Л., Штейнман Р.М., Шлезингер С., Марович М.А. (апрель 2003 г.). «DC-SIGN (CD209) опосредует заражение дендритных клеток человека вирусом денге» . Журнал экспериментальной медицины . 197 (7): 823–9. дои : 10.1084/jem.20021840 . ПМК 2193896 . ПМИД 12682107 .
- ^ Кришнан М.Н., Сукумаран Б., Пал У., Агайсс Х., Мюррей Дж.Л., Ходж Т.В., Фикриг Э. (май 2007 г.). «Rab 5 необходим для проникновения в клетки вирусов денге и Западного Нила» . Журнал вирусологии . 81 (9): 4881–5. дои : 10.1128/JVI.02210-06 . ПМК 1900185 . ПМИД 17301152 .
- ^ Джиндадамронгвех С., Теппарит С., Смит Д.Р. (май 2004 г.). «Идентификация GRP 78 (BiP) как рецепторного элемента, экспрессируемого клетками печени для серотипа 2 вируса денге» . Архив вирусологии . 149 (5): 915–27. дои : 10.1007/s00705-003-0263-x . ПМИД 15098107 . S2CID 12062084 .
- ^ Миллер Дж.Л., де Вет Б.Дж., Делет Б.Дж., Мартинес-Помарес Л., Рэдклифф К.М., Двек Р.А., Радд П.М., Гордон С. (февраль 2008 г.). «Манозный рецептор опосредует заражение макрофагов вирусом денге» . ПЛОС Патогены . 4 (2): е17. doi : 10.1371/journal.ppat.0040017 . ПМК 2233670 . ПМИД 18266465 .
- ^ Зидан Н., Ульд-Абейх М.Б., Пети-Топен И., Бедуэль Х. (декабрь 2012 г.). «Свернутые и неупорядоченные домены рибосомального белка SA человека выполняют как своеобразные, так и общие функции в качестве мембранных рецепторов» . Отчеты по биологическим наукам . 33 (1): 113–24. дои : 10.1042/BSR20120103 . ПМК 4098866 . ПМИД 23137297 .
- ^ Хусейн, Мажар; Идрис, Мухаммед; Афзал, Самия (22 июля 2015 г.). «Разработка глобального консенсуса в отношении гликопротеина оболочки вируса денге для разработки вакцин на основе эпитопов» . Современный компьютерный дизайн лекарств . 11 (1): 84–97. дои : 10.2174/1573409911666150529130134 . ISSN 1573-4099 . ПМИД 26022067 .
- ^ Зидан Н., Дюссар П., Бреманд Л., Бедуэль Х. (июль 2013 г.). «Перекрестная реактивность между человеческими IgM и четырьмя серотипами вируса денге, исследованная с помощью искусственных гомодимеров домена-III белков оболочки» . БМК Инфекционные болезни . 13 :302. дои : 10.1186/1471-2334-13-302 . ПМЦ 3701519 . ПМИД 23815496 .
- ^ Зидан Н., Дюссар П., Бреманд Л., Виллани М.Э., Бедуэль Х. (июнь 2013 г.). «Термодинамическая стабильность домена III белка оболочки флавивирусов и ее улучшение с помощью молекулярного дизайна» . Белковая инженерия, проектирование и отбор . 26 (6): 389–99. дои : 10.1093/протеин/gzt010 . ПМИД 23479674 .
- ^ Брандлер С., Раффи С., Найбург В., Френкиль М.П., Бедуэль Х., Депре П., Танжи Ф. (сентябрь 2010 г.). «Детская вакцина против кори, экспрессирующая четырехвалентный антиген денге, вызывает нейтрализующие антитела против всех четырех вирусов денге». Вакцина . 28 (41): 6730–9. doi : 10.1016/j.vaccine.2010.07.073 . ПМИД 20688034 .
- ^ Миллер С., Кастнер С., Крейнсе-Локкер Дж., Бюлер С., Бартеншлагер Р. (март 2007 г.). «Неструктурный белок 4А вируса денге представляет собой интегральный мембранный белок, вызывающий изменения мембраны 2К-регулируемым образом» . Журнал биологической химии . 282 (12): 8873–82. дои : 10.1074/jbc.M609919200 . ПМИД 17276984 . S2CID 19927382 .
- ^ Ли CM, Се X, Цзоу Дж, Ли Ш, Ли МЮ, Донг Х, Цинь КФ, Кан С, Ши ПЮ (июнь 2015 г.). «Детерминанты олигомеризации белка NS4A вируса денге» . Журнал вирусологии . 89 (12): 6171–83. дои : 10.1128/JVI.00546-15 . ПМЦ 4474302 . ПМИД 25833044 .
- ^ Цзоу Дж, Се X, Ван QY, Донг Х, Ли МЮ, Кан С, Юань Z, Ши ПЮ (апрель 2015 г.). «Характеристика взаимодействия белков NS4A и NS4B вируса денге» . Журнал вирусологии . 89 (7): 3455–70. дои : 10.1128/JVI.03453-14 . ПМК 4403404 . ПМИД 25568208 .
- ^ Остин С.К., Дауд К.А., Шреста Б., Нельсон К.А., Эделинг М.А., Джонсон С., Пирсон Т.К., Даймонд М.С., Фремонт Д.Х. (2012). «Структурные основы дифференциальной нейтрализации генотипов DENV-1 антителом, распознающим загадочный эпитоп» . ПЛОС Патогены . 8 (10): е1002930. дои : 10.1371/journal.ppat.1002930 . ПМЦ 3464233 . ПМИД 23055922 .
- ^ Лок С.М., Костюченко В., Нибаккен Г.Е., Холдэуэй Х.А., Баттисти А.Дж., Сукуполви-Петти С., Седлак Д., Фремонт Д.Х., Чипман П.Р., Рериг Дж.Т., Даймонд М.С., Кун Р.Дж., Россманн М.Г. (март 2008 г.). «Связывание нейтрализующего антитела с вирусом денге изменяет расположение поверхностных гликопротеинов» . Структурная и молекулярная биология природы . 15 (3): 312–7. дои : 10.1038/nsmb.1382 . ПМИД 18264114 . S2CID 64886 .
- ^ Мидгли CM, Фланаган А, Тран Х.Б., Деджнираттисай В., Чавансунтати К., Джумнаинсонг А, Вонгвиват В., Дуангчинда Т., Монгколсапая Дж., Граймс Дж.М., Скритон Г.Р. (май 2012 г.). «Структурный анализ перекрестно-реактивного антитела против лихорадки денге в комплексе с оболочочным доменом III раскрывает молекулярную основу перекрестной реактивности» . Журнал иммунологии . 188 (10): 4971–9. doi : 10.4049/jimmunol.1200227 . ПМЦ 3364712 . ПМИД 22491255 .
- ^ Лисова О., Харди Ф., Пети В., Бедуэль Х. (сентябрь 2007 г.). «Картирование для полноты и трансплантации группоспецифического, прерывистого, нейтрализующего эпитопа в белке оболочки вируса денге» . Журнал общей вирусологии . 88 (Часть 9): 2387–97. дои : 10.1099/vir.0.83028-0 . ПМИД 17698647 .
- ^ Кокберн Дж.Дж., Наварро Санчес М.Э., Фретес Н., Урвоас А., Старополи И., Кикути К.М., Коффи Л.Л., Аренсана Сейседос Ф., Бедуэль Х., Рей Ф.А. (февраль 2012 г.). «Механизм широкой перекрестной нейтрализации вируса денге моноклональными антителами» . Структура . 20 (2): 303–14. doi : 10.1016/j.str.2012.01.001 . ПМИД 22285214 .
- ^ Лисова О., Белкади Л., Бедуэль Х. (апрель 2014 г.). «Прямое и косвенное взаимодействие в распознавании перекрестно-нейтрализующего антитела и четырех серотипов вируса денге». Журнал молекулярного распознавания . 27 (4): 205–14. дои : 10.1002/jmr.2352 . ПМИД 24591178 . S2CID 5416842 .
- ^ Кокберн Дж.Дж., Наварро Санчес М.Е., Гонкальвес А.П., Зайцева Е., Стура Е.А., Кикути С.М., Дюкеррой С., Дюссар П., Черномордик Л.В., Лай С.Дж., Рей Ф.А. (февраль 2012 г.). «Структурное понимание механизма нейтрализации антител высших приматов против вируса денге» . Журнал ЭМБО . 31 (3): 767–79. дои : 10.1038/emboj.2011.439 . ПМЦ 3273384 . ПМИД 22139356 .
- ^ Рувински А, Гуардадо-Кальво П, Барба-Спат Г, Дюкеррой С, Вэни МК, Кикути СМ, Наварро Санчес МЭ, Дежнираттисай Ж, Вонгвиват Ж, Хауз А, Жирар-Блан Ц, Петрес С, Шепард ВЕ, Деспре П, Арензана -Сейседос Ф., Дюссар П., Монгколсапая Дж., Скритон Г.Р., Рей Ф.А. (апрель 2015 г.). «Детерминанты распознавания широко нейтрализующих человеческих антител против вирусов денге» (PDF) . Природа . 520 (7545): 109–13. Бибкод : 2015Natur.520..109R . дои : 10.1038/nature14130 . hdl : 10044/1/34173 . ПМИД 25581790 . S2CID 4394235 .
- ^ Виатрек, Мелисса. Лихорадка Денге. http://www.austincc.edu/microbio/2993p/df.htm. Архивировано 26 июля 2018 г. в Wayback Machine.
- ^ Jump up to: а б с «Денге и тяжелая денге» . Всемирная организация здравоохранения (ВОЗ) . 17 марта 2023 г. Проверено 8 октября 2023 г.
- ^ Jump up to: а б ДЕПАРТАМЕНТ ЗДРАВООХРАНЕНИЯ ОКРУГА КОЛУМБИИ. Информационный бюллетень о лихорадке Денге. https://doh.dc.gov/sites/default/files/dc/sites/doh/publication/attachments/Dengue_Fever.pdf
- ^ Jump up to: а б Лучшее здоровье. Болезнь, вызванная вирусом Денге. https://www.betterhealth.vic.gov.au/health/conditionsandtreatments/Dengue-virus-disease?viewAsPdf=true
- ^ Центры по контролю и профилактике заболеваний. Эпидемиология. https://www.cdc.gov/dengue/epidemiology/
- ^ Центры по контролю и профилактике заболеваний. Денге. https://wwwnc.cdc.gov/travel/yellowbook/2016/infectious-diseases-related-to-travel/dengue
- ^ Кин Н.Д., Эль-Кушайри А.Е., Ахмед А.М., Сафи А., Магид С.А., Мехьяр С.М. и др. (февраль 2020 г.). «Связь аллергических симптомов с инфекцией денге и ее тяжестью: систематический обзор и метаанализ» . Вирусология Синика . 35 (1): 83–92. дои : 10.1007/s12250-019-00165-6 . ПМК 7035405 . ПМИД 31637633 .
- ^ Ачесон, Николас Х. (2011). Основы молекулярной вирусологии, 2-е изд . Уайли.
- ^ Деджнираттисай В., Джумнаинсонг А., Онсирисакул Н., Фиттон П., Васанаватана С., Лимпитикул В., Путтикхант С., Эдвардс С., Дуангчинда Т., Супаса С., Чавансунтати К., Маласит П., Монгколсапая Дж., Скритон Дж. (май 2010 г.). «Перекрестно реагирующие антитела усиливают инфекцию вируса денге у людей» . Наука . 328 (5979): 745–8. Бибкод : 2010Sci...328..745D . дои : 10.1126/science.1185181 . ПМЦ 3837288 . ПМИД 20448183 .
- ^ Даймонд MS (сентябрь 2009 г.). «Механизмы уклонения флавивирусов от противовирусного ответа на интерферон I типа» . Журнал исследований интерферона и цитокинов . 29 (9): 521–30. дои : 10.1089/jir.2009.0069 . ПМИД 19694536 .
- ^ Джонс М., Дэвидсон А., Хибберт Л., Грюнвальд П., Шлаак Дж., Болл С., Фостер Г.Р., Джейкобс М. (май 2005 г.). «Вирус денге ингибирует передачу сигналов альфа-интерферона за счет снижения экспрессии STAT2» . Журнал вирусологии . 79 (9): 5414–20. doi : 10.1128/JVI.79.9.5414-5420.2005 . ПМЦ 1082737 . ПМИД 15827155 .
- ^ Хо Л.Дж., Хунг Л.Ф., Венг С.И., Ву В.Л., Чоу П., Линь Ю.Л., Чанг Д.М., Тай Т.Я., Лай Дж.Х. (июнь 2005 г.). «Вирус денге 2-го типа противодействует противовирусному эффекту ИФН-альфа, но не противовирусному эффекту ИФН-гамма посредством подавления передачи сигналов Tyk2-STAT в дендритных клетках человека» . Журнал иммунологии . 174 (12): 8163–72. дои : 10.4049/jimmunol.174.12.8163 . ПМИД 15944325 . S2CID 83577905 .
- ^ Муньос-Жордан Х.Л., Санчес-Бургос Г.Г., Лоран-Ролле М., Гарсиа-Састре А. (ноябрь 2003 г.). «Ингибирование передачи сигналов интерферона вирусом денге» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (24): 14333–8. Бибкод : 2003PNAS..10014333M . дои : 10.1073/pnas.2335168100 . ПМЦ 283592 . ПМИД 14612562 .
- ^ Ашур Дж., Лоран-Ролле М., Ши П.Ю., Гарсиа-Састре А. (июнь 2009 г.). «NS5 вируса денге опосредует связывание и деградацию STAT2» . Журнал вирусологии . 83 (11): 5408–18. дои : 10.1128/JVI.02188-08 . ПМК 2681973 . ПМИД 19279106 .
- ^ Маццон М., Джонс М., Дэвидсон А., Чейн Б., Джейкобс М. (октябрь 2009 г.). «Вирус денге NS5 ингибирует передачу сигналов интерферона-альфа, блокируя преобразователь сигнала и активатор фосфорилирования транскрипции 2» . Журнал инфекционных болезней . 200 (8): 1261–70. дои : 10.1086/605847 . ПМИД 19754307 .
- ^ Моррисон Дж., Агирре С., Фернандес-Сесма А. (март 2012 г.). «Уклонение от врожденного иммунитета вирусом Денге» . Вирусы . 4 (3): 397–413. дои : 10.3390/v4030397 . ПМК 3347034 . ПМИД 22590678 .
- ^ Юсоф Р., Клам С., Ветцель М., Мурти Х.М., Падманабхан Р. (апрель 2000 г.). «Очищенная сериновая протеаза NS2B/NS3 вируса денге типа 2 проявляет зависимость от кофактора NS2B при расщеплении субстратов с двухосновными аминокислотами in vitro» . Журнал биологической химии . 275 (14): 9963–9. дои : 10.1074/jbc.275.14.9963 . ПМИД 10744671 . S2CID 30345627 .
- ^ Родригес-Мадос-младший, Белича-Вильянуэва А, Берналь-Рубио Д, Ашур Дж, Эйллон Дж, Фернандес-Сесма А (октябрь 2010 г.). «Ингибирование ответа интерферона типа I в дендритных клетках человека инфекцией вируса денге требует каталитически активного комплекса NS2B3» . Журнал вирусологии . 84 (19): 9760–74. дои : 10.1128/JVI.01051-10 . ПМЦ 2937777 . ПМИД 20660196 .
- ^ Ю С.И., Чанг Т.Х., Лян Дж.Дж., Чан Р.Л., Ли Ю.Л., Ляо С.Л., Линь Ю.Л. (июнь 2012 г.). Даймонд М.С. (ред.). «Вирус денге нацелен на адаптерный белок MITA, чтобы подорвать врожденный иммунитет хозяина» . ПЛОС Патогены . 8 (6): e1002780. дои : 10.1371/journal.ppat.1002780 . ПМК 3386177 . ПМИД 22761576 .
- ^ Jump up to: а б с д Конвей М.Дж., Лондоно-Рентерия Б., Трупен А., Уотсон А.М., Климстра В.Б., Фикриг Э., Колпиттс Т.М. (сентябрь 2016 г.). «Белок слюны Aedes aegypti D7 подавляет инфекцию вируса денге» . PLOS Забытые тропические болезни . 10 (9): e0004941. дои : 10.1371/journal.pntd.0004941 . ПМК 5025043 . ПМИД 27632170 .
- ^ Лондоно-Рентерия Б.Л., Шакери Х., Розо-Лопес П., Конвей М.Дж., Дагган Н., Джабери-Дураки М., Колпиттс Т.М. (2018). «Белки слюны Aedes aegypti D7 в Колумбии» . Границы общественного здравоохранения . 6 : 111. дои : 10.3389/fpubh.2018.00111 . ПМЦ 5968123 . ПМИД 29868532 .
- ^ Jump up to: а б Ист, Сьюзи (6 апреля 2016 г.). «Первая в мире вакцина от лихорадки денге запущена на Филиппинах» . CNN . Проверено 17 октября 2016 г.
- ^ Jump up to: а б «Заявление STIKO о недавно одобренной живой вакцине против денге (Qdenga)» (на немецком языке). Институт Роберта Коха . 15 февраля 2023 г. Проверено 13 апреля 2023 г.
- ^ «Вакцина против денге: документ с изложением позиции ВОЗ – сентябрь 2018 г.» (PDF) . Еженедельный эпидемиологический журнал . 36 (93): 457–76. 7 сентября 2018 года . Проверено 12 апреля 2019 г.
- ^ «Вакцина Санофи от денге одобрена в 11 странах» . Рейтер . 2016 . Проверено 13 августа 2017 г.
- ^ «Вакцина от лихорадки денге доступна в Индонезии» . 17 октября 2016 г.
- ↑ Разработка вакцины . Архивировано 21 августа 2019 г. в Wayback Machine , Инициатива по вакцине против денге, ноябрь 2012 г., по состоянию на 5 ноября 2013 г.
- ^ Шмальджон А.Л., Макклейн Д. (1996). «Гл. 54: Альфавирусы (Togaviridae) и флавивирусы (Flaviviridae)» . У барона С. (ред.). Медицинская микробиология (4-е изд.). Медицинский филиал Техасского университета в Галвестоне. ISBN 978-0-9631172-1-2 . НБК7633.
- ^ Торреси Дж., Тапиа-Коньер Р., Марголис Х. (2013). «Подготовка к внедрению вакцины против денге: рекомендации 1-го Международного совещания против денге v2V» . PLOS Забытые тропические болезни . 7 (9): е2261. дои : 10.1371/journal.pntd.0002261 . ПМЦ 3784468 . ПМИД 24086776 .
- ^ «Вторая фаза III исследования вакцины против денге Aces компании Sanofi Pasteur» . ОБЩ Основные новости . Новости генной инженерии и биотехнологии. 3 сентября 2014 г.
- ^ Первая одобренная FDA вакцина для профилактики болезни денге в эндемичных регионах https://www.fda.gov/news-events/press-announcements/first-fda-approved-vaccine-prevention-dengue-disease-endemic-regions
- ^ Люнг, Д.; Шредер, К.; Уайт, Х.; Фанг, Северная Каролина; Стермер, MJ; Аббенанте, Г.; Мартин, Дж.Л.; Янг, пиарщик; Фэрли, ДП (07 декабря 2001 г.). «Активность рекомбинантной протеазы NS3 вируса денге 2 в присутствии укороченного кофактора NS2B, небольших пептидных субстратов и ингибиторов» . Журнал биологической химии . 276 (49): 45762–45771. дои : 10.1074/jbc.M107360200 . hdl : 10072/347872 . ISSN 0021-9258 . ПМИД 11581268 . S2CID 26124089 .
- ^ Дражич, Тонко; Копф, Сара; Корридан, Джеймс; Лейтольд, Мила М.; Бертоша, Бранимир; Кляйн, Кристиан Д. (9 января 2020 г.). «Пептид-β-лактамные ингибиторы протеазы NS2B-NS3 вируса Денге и Западного Нила демонстрируют два различных режима связывания» . Журнал медицинской химии . 63 (1): 140–156. doi : 10.1021/acs.jmedchem.9b00759 . ISSN 0022-2623 . ПМИД 31804823 . S2CID 208768169 .
- ^ Нгуен Н.М., Тран К.Н., Фунг Л.К., Дуонг К.Т., Хуинь Х.А., Фаррар Дж. и др. (май 2013 г.). «Рандомизированное двойное слепое плацебо-контролируемое исследование балапиравира, ингибитора полимеразы, у взрослых пациентов с денге» . Журнал инфекционных болезней . 207 (9): 1442–50. дои : 10.1093/infdis/jis470 . ПМЦ 3610419 . ПМИД 22807519 .
- ^ Чен Ю.Л., Абдул Гафар Н., Каруна Р., Фу Ю., Лим С.П., Шул В. и др. (февраль 2014 г.). «Активация мононуклеаров периферической крови инфекцией вируса денге депотенциирует балапиравир» . Журнал вирусологии . 88 (3): 1740–7. дои : 10.1128/JVI.02841-13 . ПМЦ 3911617 . ПМИД 24257621 .
- ^ https://repository.unair.ac.id/127532/
- ^ https://repository.unair.ac.id/127532/
Внешние ссылки
[ редактировать ]- 3D-структуры электронной микроскопии вируса денге из Банка данных EM (EMDB)
- «В Бразилии выпускают «хороших» комаров для борьбы с лихорадкой денге» . BBC News Латинская Америка и Карибский бассейн . 24 сентября 2014 г.
| Базы данных органов управления : Национальные |
|---|