Грызун
| Грызун Временной диапазон: | |
|---|---|
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Мирордер: | Простая идентичность |
| Заказ: | Грызуны Боудич , 1821 г. |
| Подзаказы | |
 | |
| Совокупный ареал всех видов грызунов (не включая интродуцированные популяции) | |





Грызуны (от лат.rodere , , «грызть») — ( / r млекопитающие отряда Rodentia oʊ ˈ d ɛ n ʃ ə / roh - DEN - shə ) для которых характерна единственная пара непрерывно растущих резцов в каждой из верхняя и нижняя челюсти . Около 40% всех видов млекопитающих составляют грызуны. Они являются родными для всех основных суши, за исключением Новой Зеландии , Антарктиды и нескольких океанических островов, хотя впоследствии они были завезены на большую часть этих суши в результате деятельности человека.
Грызуны чрезвычайно разнообразны по своей экологии и образу жизни, их можно встретить практически во всех наземных средах обитания, включая антропогенную среду. Виды могут вести древесный , роющий образ жизни , скачкообразный /рихохетальный (прыгающий на задних лапах) или полуводный образ жизни. Однако все грызуны имеют несколько общих морфологических особенностей, в том числе наличие только одной верхней и нижней пары постоянно растущих резцов. К хорошо известным грызунам относятся мыши , крысы , белки , луговые собачки , дикобразы , бобры , морские свинки и хомяки . Кролики , зайцы и пищухи , у которых также непрерывно растут резцы (но имеют две пары верхних резцов вместо одной), [1] когда-то были включены в состав грызунов, но теперь считаются отдельным отрядом Lagomorpha . , Rodentia и Lagomorpha — сестринские группы , имеющие одного общего предка и образующие кладу Glires Тем не менее .
Большинство грызунов — мелкие животные с крепким телом, короткими конечностями и длинными хвостами. Они используют свои острые резцы, чтобы перегрызать пищу, рыть норы и защищаться. Большинство из них едят семена или другой растительный материал, но у некоторых рацион более разнообразен. Они, как правило, социальные животные, и многие виды живут в обществах со сложными способами общения друг с другом. Спаривание среди грызунов может варьироваться от моногамии до полигинии и беспорядочных половых связей . У многих из них рождаются недоразвитые , альтрициальные детеныши, тогда как другие при рождении являются преждевременными (относительно хорошо развитыми).
грызунов Летопись окаменелостей восходит к палеоцену на суперконтиненте Лавразия . Грызуны сильно разнообразились в эоцене , расселившись по континентам, иногда даже пересекая океаны . Грызуны проникли в Южную Америку и на Мадагаскар из Африки и до появления Homo sapiens были единственными наземными плацентарными млекопитающими , достигшими и колонизировавшими Австралию.
Грызунов использовали в качестве еды, одежды, домашних животных и лабораторных животных в исследованиях. Некоторые виды, в частности бурая крыса , черная крыса и домовая мышь , являются серьезными вредителями , поедая и портя хранящиеся у человека продукты питания и распространяя болезни. Случайно завезенные виды грызунов часто считаются инвазивными и стали причиной исчезновения многих видов, таких как островные птицы, например дронт , ранее изолированный от наземных хищников.
Характеристики [ править ]
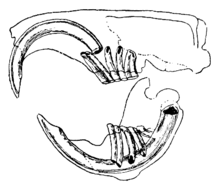

Отличительной особенностью грызунов являются пары непрерывно растущих, острых как бритва резцов с открытыми корнями . [2] Эти резцы имеют толстый слой эмали спереди и мало эмали сзади. [3] Поскольку они не перестают расти, животное должно продолжать их изнашивать, чтобы они не дотянулись до черепа и не проткнули его. Когда резцы стираются друг о друга, более мягкий дентин на задней части зубов стирается, оставляя острый край эмали в форме лезвия долота . [4] У большинства видов имеется до 22 зубов без клыков и передних премоляров . щель, или диастема Между резцами и щечными зубами у большинства видов возникает . Это позволяет грызунам засасывать щеки или губы, чтобы защитить рот и горло от древесной стружки и других несъедобных материалов, выбрасывая эти отходы по бокам рта. [ нужна ссылка ] Шиншиллы и морские свинки имеют рацион с высоким содержанием клетчатки; их коренные зубы не имеют корней и растут непрерывно, как и резцы. [5]
У многих видов коренные зубы относительно большие, сложной структуры, с сильно заостренными или ребристыми краями. Коренные зубы грызунов хорошо приспособлены для измельчения пищи на мелкие частицы. [2] Мускулатура челюстей сильная. Нижняя челюсть при грызении выдвигается вперед, а при жевании оттягивается назад. [3] Для грызения используются резцы, а для жевания – коренные зубы, однако из-за черепной анатомии грызунов эти способы кормления не могут использоваться одновременно и считаются взаимоисключающими. [6] У грызунов жевательная мышца играет ключевую роль в жевании, составляя 60–80% общей мышечной массы жевательных мышц и отражает растительноядный рацион грызунов. [7] Группы грызунов отличаются расположением челюстных мышц и связанных с ними структур черепа как от других млекопитающих, так и между собой.
Sciuromorpha , что позволяет им эффективно , такие как восточная серая белка , имеют большую глубокую жевательную мышцу кусать резцами. , У миоморф таких как коричневая крыса, увеличены височные и жевательные мышцы, что позволяет им мощно жевать коренными зубами. [8] У грызунов жевательные мышцы прикрепляются за глазами и способствуют ошеломлению глаз, которое возникает во время грызения, когда быстрое сокращение и расслабление мышцы заставляет глазные яблоки двигаться вверх и вниз. [8] Hystricomorpha , такие как морская свинка, имеют более крупные поверхностные жевательные мышцы и меньшие глубокие жевательные мышцы, чем крысы или белки, что, возможно , делает их менее эффективными при кусании резцами, но их увеличенные внутренние крыловидные мышцы могут позволить им двигать челюсть дальше в сторону. при жевании. [9] представляет Защечный мешок собой особую морфологическую особенность, используемую для хранения пищи, и он очевиден у определенных подгрупп грызунов, таких как кенгуровые крысы, хомяки, бурундуки и суслики, у которых есть два мешка, которые могут располагаться ото рта до передней части плеч. [10] Настоящие мыши и крысы не содержат этой структуры, но их щеки эластичны из-за высокой степени мускулатуры и иннервации в этой области. [11]

Хотя самый крупный вид, капибара , может весить до 66 кг (146 фунтов), большинство грызунов весят менее 100 г (3,5 унции). Грызуны имеют разнообразную морфологию, но обычно имеют приземистое тело и короткие конечности. [2] Передние конечности обычно имеют пять пальцев, включая противопоставленный большой палец, а задние конечности имеют от трех до пяти пальцев. Локоть придает предплечьям большую гибкость. [4] Большинство видов стопоходят , ходят как на ладонях, так и на подошвах ног и имеют когти. Когти роющих видов обычно длинные и крепкие, а у древесных грызунов когти короче и острые. Виды грызунов используют широкий спектр методов передвижения, включая ходьбу на четвероногих ногах , бег, рытье норы, лазание, прыжки на двух ногах ( крысы-кенгуру и прыгающие мыши ), плавание и даже скольжение. [4] Чешуйчатохвостые белки и летяги , хотя и не являются близкими родственниками, могут планировать с дерева на дерево, используя парашютоподобные перепонки , которые простираются от передних к задним конечностям. [12] Агути и быстроногие и похожи на антилоп , имеют пальцы ногти, похожие на копыта. У большинства грызунов есть хвосты, которые могут быть самых разных форм и размеров. Некоторые хвосты цепкие , как у евразийской промысловой мыши , а шерсть на хвостах может варьироваться от густой до полностью лысой. Хвост иногда используется для общения, например, когда бобры хлопают хвостом по поверхности воды, а домашние мыши трясут хвостом, сигнализируя о тревоге. У некоторых видов есть рудиментарные хвосты или они вообще отсутствуют. [2] У некоторых видов хвост способен к регенерации, если отломать его часть. [4]

У грызунов обычно хорошо развиты обоняние , слух и зрение. Ночные виды часто имеют увеличенные глаза, а некоторые чувствительны к ультрафиолетовому свету. У многих видов есть длинные чувствительные усы или вибриссы, позволяющие прикасаться или «взбивать» . [14] Действием усов в основном управляет ствол мозга, который сам провоцируется корой. [14] Однако Легг и др. 1989 обнаружили альтернативный контур между корой и усами через контуры мозжечка, а Hemelt & Keller 2008 — верхние бугорки. [14] У некоторых грызунов есть защечные мешки , которые могут быть покрыты мехом. Для очистки их можно вывернуть наизнанку. У многих видов язык не может дотянуться до резцов. У грызунов эффективная пищеварительная система, поглощающая почти 80% потребляемой энергии. При употреблении целлюлозы пища размягчается в желудке и попадает в слепую кишку , где бактерии восстанавливают ее до углеводных элементов. Затем грызун практикует копрофагию , поедая собственные фекальные гранулы, чтобы питательные вещества могли усваиваться кишечником. Поэтому грызуны часто производят твердые и сухие фекальные шарики. [2] Хорн и др. 2013 год [15] делает вывод, что у грызунов полностью отсутствует способность к рвоте. [16] [17] [18] [19] У многих видов пенис содержит кость — бакулюм ; яички могут располагаться либо на животе, либо в паху. [4]
Половой диморфизм встречается у многих видов грызунов. У некоторых грызунов самцы крупнее самок, а у других все наоборот. Половой диморфизм по мужскому типу характерен для сусликов , кенгуровых крыс, одиночных землекопов и карманных сусликов ; вероятно, это развилось из-за полового отбора и более активных боев между мужчинами. Половой диморфизм с женским уклоном существует среди бурундуков и прыгающих мышей . Непонятно, почему происходит такая закономерность, но в случае бурундуков желто-сосновых самцы могли выбирать более крупных самок из-за их большего репродуктивного успеха. У некоторых видов, например полевок , половой диморфизм может варьироваться от популяции к популяции. У рыжих полевок самки обычно крупнее самцов, но в альпийских популяциях встречается половой диморфизм по признаку самцов, возможно, из-за отсутствия хищников и большей конкуренции между самцами. [20]
Распространение и среда обитания [ править ]

Грызуны, одна из самых распространенных групп млекопитающих, встречаются на всех континентах, кроме Антарктиды. Это единственные наземные плацентарные млекопитающие, колонизировавшие Австралию и Новую Гвинею без вмешательства человека. Люди также позволили животным распространиться на многие отдаленные океанические острова (например, полинезийская крыса ). [4] Грызуны приспособились практически к любой наземной среде обитания: от холодной тундры (где они могут жить под снегом) до жарких пустынь.
Некоторые виды, такие как древесные белки и дикобразы Нового Света, ведут древесный образ жизни , в то время как некоторые, такие как суслики , туко-туко и слепыши, живут почти полностью под землей, где они строят сложные системы нор. Другие обитают на поверхности земли, но могут иметь нору, в которую могут укрыться. Бобры и ондатры известны тем, что ведут полуводный образ жизни. [2] но грызуном, лучше всего приспособленным к водной жизни, является, вероятно, безухая водяная крыса из Новой Гвинеи. [21] Грызуны также процветают в созданных человеком средах, таких как сельскохозяйственные и городские районы . [22]

Хотя некоторые виды являются обычными вредителями для человека, грызуны также играют важную экологическую роль. [2] Некоторые грызуны считаются ключевыми видами и инженерами экосистем в своих средах обитания. На Великих равнинах Северной Америки роющая деятельность луговых собачек играет важную роль в аэрации почвы и перераспределении питательных веществ, повышая содержание органических веществ в почве и увеличивая поглощение воды. Они поддерживают эти луговые среды обитания, [23] а некоторые крупные травоядные животные, такие как бизоны и вилороги, предпочитают пастись вблизи колоний луговых собачек из-за повышенной питательной ценности корма. [24]
Истребление луговых собачек также может способствовать потере регионального и местного биоразнообразия , усилению хищничества семян, а также возникновению и распространению инвазивных кустарников. [23] Роющие грызуны могут поедать плодовые тела грибов и распространять споры через их фекалии, тем самым позволяя грибам рассеяться и образовывать симбиотические отношения с корнями растений (которые обычно не могут процветать без них). Таким образом, эти грызуны могут играть роль в поддержании здоровья лесов. [25]
Во многих регионах с умеренным климатом бобры играют важную гидрологическую роль. Строя свои плотины и хатки, бобры меняют русла ручьев и рек. [26] и позволить создать обширные места обитания водно-болотных угодий. Одно исследование показало, что инженерная деятельность бобров приводит к 33-процентному увеличению количества видов травянистых растений в прибрежных районах . [27] Другое исследование показало, что бобры увеличивают популяцию дикого лосося. [28] Между тем, некоторые грызуны считаются вредителями из-за их широкого ареала. [29]
Поведение и история жизни [ править ]
Кормление [ править ]

Большинство грызунов травоядны и питаются исключительно растительным материалом, таким как семена, стебли, листья, цветы и корни. Некоторые из них всеядны , а некоторые — хищники. [3] Полевка — типичный травоядный грызун, питается травами, разнотравьем, корневыми клубнями, мхом и другой растительностью, а зимой подгрызает кору. Иногда он поедает беспозвоночных, таких как личинки насекомых. [30] Равнинный карманный суслик поедает растительный материал, найденный под землей во время прокладки туннелей, а также собирает траву, корни и клубни в свои защечные мешки и прячет их в подземных кладовых. [31]
Техасский карманный суслик избегает выхода на поверхность для кормления, захватывая челюстями корни растений и затягивая их вниз, в свою нору. Также практикуется копрофагия. [32] Африканская сумчатая крыса добывает пищу на поверхности, собирая все, что может быть съедобным, в свои вместительные защечные мешки, пока ее морда не выпирает вбок. Затем он возвращается в свою нору, чтобы перебрать собранный материал и поедать питательные вещества. [33]
Виды агути — одна из немногих групп животных, которые могут вскрывать большие коробочки плодов бразильского ореха . Внутри слишком много семян, чтобы их можно было съесть за один прием, поэтому агути уносит некоторые из них и прячет. Это способствует распространению семян, поскольку те семена, которые агути не удалось найти, при прорастании находятся далеко от родительского дерева. Другие орехоплодные деревья, как правило, осенью приносят избыток фруктов. Их слишком много, чтобы съесть их за один прием, а белки собирают и хранят излишки в расщелинах и дуплах деревьев. В пустынных регионах семена часто доступны только на короткий период времени. Кенгуровая крыса собирает все, что может найти, и хранит их в кладовых своей норы. [33]

Стратегия борьбы с сезонным изобилием — есть как можно больше и хранить излишки питательных веществ в виде жира. Сурки делают это, и осенью они могут быть на 50% тяжелее, чем весной. Во время долгой зимней спячки они полагаются на свои жировые запасы . [33] Бобры питаются листьями, почками и внутренней корой растущих деревьев, а также водными растениями. Они запасают еду на зиму, срубая осенью небольшие деревья и лиственные ветки и погружая их в пруд, втыкая концы в ил, чтобы закрепить их. Здесь они могут получить доступ к своим запасам пищи под водой, даже когда их пруд замерз. [34]
Хотя грызуны традиционно считались травоядными, большинство мелких грызунов случайно включают в свой рацион насекомых, червей, грибов, рыбу или мясо, а некоторые стали специализированными и полагаются на пищу животного происхождения. Функционально-морфологическое исследование зубной системы грызунов подтверждает идею о том, что примитивные грызуны были всеядными, а не травоядными. Исследования литературы показывают, что многие представители Sciuromorpha и Myomorpha, а также некоторые представители Hystricomorpha либо включали в свой рацион продукты животного происхождения, либо были готовы есть такую пищу, когда ей предлагали ее в неволе. Исследование содержимого желудка североамериканской белоногой мыши , обычно считающейся травоядной, показало 34% животного вещества. [35]
К более специализированным хищникам относятся землероековые крысы филиппинские , которые питаются насекомыми и мягкотелыми беспозвоночными, и австралийская водяная крыса , которая пожирает водных насекомых, рыбу, ракообразных, мидий, улиток, лягушек, птичьи яйца и водоплавающих птиц. [35] [36] Мышь -кузнечик из засушливых районов Северной Америки питается насекомыми, скорпионами и другими мелкими мышами, и лишь небольшая часть ее рациона составляет растительный материал. У него коренастое тело с короткими ногами и хвостом, но он проворный и может легко одолеть добычу такого же размера, как он сам. [37]
Социальное поведение [ править ]

Грызуны демонстрируют широкий спектр типов социального поведения: от кастовой системы млекопитающих до голого землекопа и [38] обширный «город» колониальной луговой собачки , [39] через семейные группы к самостоятельной, одиночной жизни съедобной сони . У взрослых сонь могут перекрываться диапазоны кормления, но они живут в отдельных гнездах и питаются отдельно, ненадолго собираясь вместе в период размножения для спаривания. Карманный суслик также является одиночным животным вне сезона размножения: каждая особь роет сложную систему туннелей и поддерживает свою территорию. [40]
Более крупные грызуны, как правило, живут семейными группами, где родители и их потомство живут вместе, пока молодые не разойдутся. Бобры живут большими семьями, обычно с парой взрослых особей, детенышами этого года, потомством прошлого года, а иногда и молодняком старшего возраста. [41] Коричневые крысы обычно живут небольшими колониями, состоящими из шести самок, делящих нору, и одного самца, защищающего территорию вокруг норы. При высокой плотности населения эта система нарушается, и самцы демонстрируют иерархическую систему доминирования с перекрывающимися ареалами. Потомство женского пола остается в колонии, а молодые самцы расходятся. [42] Луговая полевка моногамна . и образует пару на всю жизнь Вне сезона размножения степные полевки живут вместе с другими небольшими колониями. Самец не агрессивен по отношению к другим самцам до тех пор, пока не спарится, после чего он защищает территорию, самку и гнездо от других самцов. Пара ютится вместе, ухаживает друг за другом и разделяет обязанности по гнездованию и воспитанию детенышей. [43]

Среди наиболее социальных грызунов — суслики, которые обычно образуют колонии на основе родства самок, а самцы расселяются после отъема и становятся кочевниками, когда становятся взрослыми. Сотрудничество сусликов варьируется в зависимости от вида и обычно включает в себя подачу сигналов тревоги, защиту территорий, обмен пищей, защиту мест гнездования и предотвращение детоубийства. [44] Чернохвостая луговая собачка образует большие города, занимающие многие гектары. Норы не соединяются между собой, а выкапываются и занимают территориальные семейные группы, известные как кружки. Группа часто состоит из взрослого самца, трех-четырех взрослых самок, нескольких неразмножающихся годовалых особей и потомства текущего года. Особи внутри котерий дружелюбны друг к другу, но враждебны по отношению к посторонним. [39]
Возможно, наиболее крайними примерами колониального поведения грызунов являются эусоциальный голый землекоп и дамаралендский землекоп . Голый землекоп обитает полностью под землей и может образовывать колонии до 80 особей. В колонии размножается только одна самка и до трех самцов, остальные же члены более мелкие, бесплодные и выполняют функцию рабочих. Некоторые особи имеют средний размер. Они помогают в выращивании детенышей и могут заменить потомство в случае его смерти. [38] Для дамаралендского слепыша характерно наличие одного репродуктивно активного самца и самки в колонии, где остальные животные не являются по-настоящему стерильными, но становятся плодовитыми только в том случае, если они создают собственную колонию. [45]
Общение [ править ]
Обонятельный [ править ]

Грызуны используют запаховую маркировку во многих социальных контекстах, включая меж- и внутривидовое общение, маркировку троп и установление территорий. Их моча предоставляет генетическую информацию об особях, включая вид, пол и индивидуальную идентичность, а также метаболическую информацию о доминировании, репродуктивном статусе и здоровье. Соединения, полученные из главного комплекса гистосовместимости (MHC), связываются с несколькими белками мочи. Запах хищника подавляет его поведение, связанное с запахом. [46]
Грызуны способны узнавать близких родственников по запаху, что позволяет им проявлять кумовство (предпочтительное поведение по отношению к своим сородичам), а также избегать инбридинга. Это распознавание родственников осуществляется по обонятельным признакам мочи, кала и железистых выделений. Основная оценка может включать MHC, где степень родства двух людей коррелирует с общими для них генами MHC. нелетучие основные белки мочи (MUP), которые действуют как переносчики феромонов При межродственном общении, где требуются более постоянные маркеры запаха, например, на территориальных границах, также можно использовать . MUP также могут сигнализировать об индивидуальной идентичности: каждый самец домашней мыши ( Mus musculus ) выделяет мочу, содержащую около дюжины генетически закодированных MUP. [47]
Домовые мыши откладывают мочу, содержащую феромоны, для территориальной маркировки, индивидуального и группового распознавания, а также социальной организации. [48] Территориальные бобры и рыжие белки исследуют и знакомятся с запахами своих соседей и менее агрессивно реагируют на их вторжение, чем на нетерриториальные «плавающие» или чужаки. Это известно как « эффект дорогого врага ». [49] [50]
Слуховой [ править ]

Многие виды грызунов, особенно те, которые ведут дневной и социальный образ жизни, имеют широкий спектр сигналов тревоги, которые издаются, когда они чувствуют угрозу. От этого есть как прямые, так и косвенные выгоды. Потенциальный хищник может остановиться, когда узнает, что его обнаружили, или сигнал тревоги может позволить представителям своего вида или родственным особям предпринять действия по уклонению. [51] Некоторые виды, например луговые собачки, имеют сложные системы сигнализации против хищников. У этих видов могут быть разные призывы к разным хищникам (например, воздушным хищникам или наземным хищникам), и каждый призыв содержит информацию о характере конкретной угрозы. [52] Актуальность угрозы также передается акустическими свойствами звонка. [53]
Социальные грызуны имеют более широкий диапазон вокализации, чем одиночные виды. было обнаружено пятнадцать различных типов криков У взрослых слепышей Катаба , а у молодых - четыре. [54] Точно так же обыкновенный дегу , еще один социальный роющий грызун, демонстрирует широкий спектр методов общения и имеет сложный голосовой репертуар, включающий пятнадцать различных категорий звуков. [55] Ультразвуковые звонки играют роль в социальном общении между сонями и используются, когда особи находятся вне поля зрения друг друга. [56]
Домовые мыши используют как звуковые, так и ультразвуковые сигналы в различных контекстах. Звуковые вокализации часто можно услышать во время агонистических или агрессивных столкновений, тогда как ультразвук используется при половом общении, а также детенышами, когда они выпадают из гнезда. [48]
Лабораторные крысы (коричневые крысы, Rattus norvegicus ) издают короткие, высокочастотные ультразвуковые звуки во время предположительно приятных переживаний, таких как грубая игра, при ожидании обычных доз морфия , во время спаривания и при щекотке. Вокализация, описываемая как отчетливое «чириканье», сравнивается со смехом и интерпретируется как ожидание чего-то полезного. В клинических исследованиях щебетание связано с положительными эмоциональными ощущениями, а благодаря щекотке возникает социальная связь, в результате чего крысы приучаются искать щекотки. Однако с возрастом крыс склонность к чириканию снижается. Как и большинство крысиных вокализаций, частота щебетания слишком высока, чтобы люди могли его услышать без специального оборудования, поэтому детекторы летучих мышей . для этой цели использовались [57]
Визуальный [ править ]
Грызуны, как и все плацентарные млекопитающие, за исключением приматов, имеют в сетчатке всего два типа светочувствительных колбочек: [58] коротковолновый тип «синий-УФ» и средневолновый «зеленый» тип. Поэтому их классифицируют как дихроматы ; однако они визуально чувствительны к ультрафиолетовому (УФ) спектру и поэтому могут видеть свет, который не могут видеть люди. Функции этой чувствительности к УФ-излучению не всегда ясны. У дегу , например, живот отражает больше ультрафиолета, чем спина. Поэтому, когда дегу встает на задние лапы, что он делает, когда встревожен, он подставляет свой живот другим дегу, и ультрафиолетовое зрение может помочь передать тревогу. Когда он стоит на четвереньках, его спина с низким уровнем отражения ультрафиолета может сделать дегу менее заметным для хищников. [59] Ультрафиолетового света много днем, но не ночью. В утренние и вечерние сумеречные часы наблюдается значительное увеличение соотношения ультрафиолетового и видимого света. Многие грызуны активны в сумеречные часы (сумеречная активность), и чувствительность к ультрафиолету в это время будет полезной. Отражательная способность ультрафиолета имеет сомнительное значение для ночных грызунов. [60]
Моча многих грызунов (например, полевок, дегу, мышей, крыс) сильно отражает ультрафиолетовый свет, и это можно использовать в общении, оставляя как видимые, так и обонятельные метки. [61] Однако количество отражаемого УФ-излучения со временем уменьшается, что в некоторых случаях может быть невыгодным; обыкновенная пустельга может различать старые и свежие следы грызунов и с большим успехом охотится на недавно отмеченных маршрутах. [62]
Тактильный [ править ]

Вибрации могут служить подсказками представителям сородичей о конкретном поведении, предупреждении и избегании хищников, содержании стада или группы и ухаживании. Ближневосточный слепой землекоп был первым млекопитающим, для которого сейсмическая связь была зарегистрирована . Эти ископаемые грызуны бьются головой о стены своих туннелей. Первоначально такое поведение интерпретировалось как часть их поведения при строительстве туннелей, но в конечном итоге выяснилось, что они генерируют сейсмические сигналы с временной структурой для связи на большие расстояния с соседними землекопами. [63]
Барабанная дробь широко используется как предупреждение о хищниках или защитное действие. Его используют в основном ископаемые или полуфоссорные грызуны. [64] Знамяхвостая кенгуровая крыса издает несколько сложных моделей барабанного боя в различных контекстах, один из которых — когда она сталкивается со змеей. Барабанье ногами может предупредить находящееся поблизости потомство, но, скорее всего, означает, что крыса слишком насторожена для успешной атаки, что предотвращает хищническое преследование змеи. [63] [65] Несколько исследований показали намеренное использование колебаний грунта как средства внутривидового общения во время ухаживания капского землекопа . [66] Сообщается, что игра на барабанах используется в соревнованиях между мужчинами; доминирующий самец демонстрирует свой потенциал сохранения ресурсов, барабаня по барабанам, сводя таким образом к минимуму физический контакт с потенциальными соперниками. [63]
Стратегии спаривания [ править ]

Некоторые виды грызунов моногамны: взрослый самец и самка образуют прочную парную связь . Моногамия может проявляться в двух формах; Обязательные и факультативные. При облигатной моногамии оба родителя заботятся о потомстве и играют важную роль в его выживании. Это происходит у таких видов, как калифорнийские мыши , мыши Oldfield , малагасийские гигантские крысы и бобры. У этих видов самцы обычно спариваются только со своими партнерами. Помимо усиленной заботы о молодняке, облигатная моногамия также может быть полезна для взрослых самцов, поскольку снижает шансы никогда не найти себе пару или спариваться с бесплодной самкой. При факультативной моногамии самцы не обеспечивают прямой родительской заботы и остаются с одной самкой, поскольку не могут получить доступ к другим из-за пространственного рассредоточения. Прерийные полевки, по-видимому, являются примером такой формы моногамии, когда самцы охраняют и защищают самок в пределах их близости. [67]
У полигинных видов самцы будут пытаться монополизировать и спариваться с несколькими самками. Как и моногамия, полигиния у грызунов может проявляться в двух формах; Оборона и необорона. Защитная полигиния предполагает, что мужчины контролируют территории, содержащие ресурсы, привлекающие женщин. Это происходит у сусликов, таких как желтобрюхие сурки , калифорнийские суслики , колумбийские суслики и суслики Ричардсона . Самцы, имеющие территории, известны как «постоянные» самцы, а самки, живущие на территориях, известны как «постоянные» самки. В случае сурков местные самцы, похоже, никогда не теряют свою территорию и всегда выигрывают встречи с вторгающимися самцами. Также известно, что некоторые виды напрямую защищают своих самок, и последующие драки могут привести к серьезным ранениям. У видов с полигинией, не связанной с защитой, самцы не являются территориальными и широко кочуют в поисках самок, чтобы монополизировать их. Эти самцы устанавливают иерархию доминирования, при этом самцы высокого ранга имеют доступ к большинству самок. Это происходит у таких видов, как Суслики Белдинга и некоторые виды древесных белок. [67]
Беспорядочные половые связи , при которых и самцы, и самки спариваются с несколькими партнерами, также встречаются у грызунов. У таких видов, как белоногие мыши, самки рождают пометы от нескольких отцов. Распущенность приводит к усилению конкуренции сперматозоидов , а у мужчин, как правило, яички больше. У капского суслика семенники самца могут составлять 20 процентов длины головы и тела. [67] У некоторых видов грызунов есть гибкие системы спаривания, которые могут варьироваться от моногамии, полигинии и беспорядочных половых связей. [67]
Самки грызунов играют активную роль в выборе себе партнеров. Факторы, влияющие на предпочтение самок, могут включать размер, доминирование и пространственные способности самца. [68] У эусоциальных голых землекопов одна самка монополизирует спаривание как минимум с тремя самцами. [38]
У большинства видов грызунов, таких как коричневые крысы и домовые мыши, овуляция происходит в регулярном цикле, тогда как у других видов, таких как полевки, она индуцируется спариванием . Во время совокупления самцы некоторых видов грызунов вставляют брачную пробку в половое отверстие самки, как для предотвращения утечки спермы, так и для защиты от оплодотворения самки другими самцами. Самки могут вынуть пробку и сделать это либо сразу, либо через несколько часов. [68]
Метаболизм и тиреоидных гормонов йода в медиобазальном гипоталамусе изменяется в ответ на фотопериод . Гормоны щитовидной железы, в свою очередь, вызывают репродуктивные изменения. Это обнаружено Ватанабэ и др. 2004 и 2007 гг., Барретт и др. 2007, Фриман и др. 2007, и Хервиг и др. 2009 г. у сибирских хомяков , Revel et al. 2006 г. и Ясуо и др. 2007 г. на сирийских хомяках , Yasuo et al. 2007 и Росс и др. 2011 на крысах и Ono et al. 2008 год у мышей. [69]
Рождение и воспитание детей [ править ]

грызуны могут рождаться либо альтрициальными (слепыми, безволосыми и относительно недоразвитыми), либо ранними В зависимости от вида (в основном покрытыми шерстью, с открытыми и достаточно развитыми глазами). Альтрициальное состояние типично для белок и мышей, тогда как преждевременное состояние обычно встречается у таких видов, как морские свинки и дикобразы. Самки с альтрициальным потомством обычно строят тщательно продуманные гнезда перед родами и содержат их до тех пор, пока их потомство не будет отлучено от груди . Самка рожает сидя или лежа, и детеныши появляются в том направлении, в котором она смотрит. Новорожденные впервые выходят из гнезда через несколько дней после того, как открыли глаза, и поначалу продолжают регулярно возвращаться. По мере того, как они становятся старше и более развитыми, они реже посещают гнездо и покидают его навсегда после отъема от груди. [70]
У скороспелых видов матери мало вкладывают в строительство гнезда, а некоторые вообще не строят гнезда. Самка рожает стоя, и детеныши появляются позади нее. Матери этих видов поддерживают контакт со своими высокоподвижными детенышами с помощью материнских контактных криков. Несмотря на то, что не по годам развитые детеныши относительно независимы и отняты от груди в течение нескольких дней, они могут продолжать кормить грудью и ухаживать за ними со стороны своих матерей. Размеры помета грызунов также различаются, и самки с меньшими пометами проводят больше времени в гнезде, чем самки с более крупными пометами. [70]

Матери-грызуны обеспечивают как непосредственную родительскую заботу, такую как уход, уход, поиск и скопление в кучу, так и косвенную родительскую заботу, такую как хранение еды, строительство гнезда и защиту своего потомства. [70] У многих социальных видов о детенышах могут заботиться не только их родители, но и другие люди - практика, известная как совместное размножение или совместное размножение . Известно, что это происходит у чернохвостых луговых собачек и сусликов Белдинга, матери которых имеют общие гнезда и выкармливают неродственных детенышей вместе со своими собственными. Возникает вопрос, могут ли эти матери отличить своих детенышей. В патагонской маре детенышей также помещают в общие норы, но матери не позволяют кормить грудью другим детенышам, кроме своей собственной. [71]
Детоубийство существует у многих видов грызунов и может практиковаться взрослыми особями любого пола. Было предложено несколько причин такого поведения, в том числе пищевой стресс, конкуренция за ресурсы, избегание неправильного направления родительской заботы и, в случае самцов, попытки сделать мать сексуально восприимчивой. Последняя причина хорошо поддерживается у приматов и львов , но в меньшей степени у грызунов. [72] Детоубийство, по-видимому, широко распространено среди чернохвостых луговых собачек, включая детоубийство в результате вторжения самцов и самок-иммигрантов, а также случайный каннибализм собственного потомства. [73] Чтобы защититься от детоубийства со стороны других взрослых особей, самки грызунов могут использовать избегание или прямую агрессию против потенциальных преступников, многократное спаривание, территориальность или раннее прерывание беременности. [72] Фетцид может также произойти среди грызунов; у альпийских сурков доминирующие самки склонны подавлять размножение подчиненных, проявляя к ним антагонизм во время беременности. Возникающий в результате стресс приводит к аборту плода. [74]
Интеллект [ править ]

Грызуны обладают развитыми когнитивными способностями. Они могут быстро научиться избегать отравленных приманок, что делает борьбу с ними трудными вредителями. [2] Морские свинки могут учиться и запоминать сложные пути к еде. [75] Белки и кенгуровые крысы способны находить тайники с едой с помощью пространственной памяти , а не только по запаху. [76] [77]
Поскольку лабораторные мыши (домашние мыши) и крысы (коричневые крысы) широко используются в качестве научных моделей для дальнейшего понимания биологии, многое стало известно об их когнитивных способностях. Коричневые крысы демонстрируют когнитивную предвзятость , при которой обработка информации зависит от того, находятся ли они в положительном или отрицательном аффективном состоянии. [78] Например, лабораторные крысы, обученные реагировать на определенный тон, нажимая рычаг, чтобы получить вознаграждение, и нажимать другой рычаг в ответ на другой тон, чтобы избежать удара электрическим током, с большей вероятностью будут реагировать на промежуточный тон. выбирая рычаг вознаграждения, если их только что пощекотали (что им нравится), указывая на «связь между непосредственно измеряемым положительным аффективным состоянием и принятием решений в условиях неопределенности на модели животного». [79]
Лабораторные (коричневые) крысы могут обладать способностью к метапознанию — анализировать свое собственное обучение и затем принимать решения на основе того, что они знают или не знают, о чем свидетельствует выбор, который они делают, очевидно, балансируя между сложностью задач и ожидаемым вознаграждением, делая их первые животные, помимо приматов, обладающие такой способностью, [80] [81] но эти результаты оспариваются, поскольку крысы, возможно, следовали простым оперантного обусловливания . принципам [82] или поведенческая экономическая модель. [83] Коричневые крысы используют социальное обучение в самых разных ситуациях, но, возможно, особенно при формировании пищевых предпочтений. [84] [85]
и Классификация эволюция
Эволюционная история [ править ]

Зубной ряд является ключевым признаком, по которому можно распознать ископаемых грызунов, и самые ранние упоминания о таких млекопитающих относятся к палеоцену , вскоре после вымирания нептичьих динозавров около 66 миллионов лет назад. Эти окаменелости найдены в Лавразии , [86] суперконтинент, состоящий из современной Северной Америки, Европы и Азии. Дивергенция Glires — клады, состоящей из грызунов и зайцеобразных (кроликов, зайцев и пищух), от других плацентарных млекопитающих произошла в течение нескольких миллионов лет после границы мела и палеогена; грызуны и зайцеобразные затем распространились в кайнозое . [87] Некоторые данные молекулярных часов позволяют предположить, что современные грызуны (представители отряда Rodentia) появились в конце мелового периода . [88] хотя другие оценки молекулярной дивергенции согласуются с летописью окаменелостей. [89] [90]
Считается, что грызуны возникли в Азии, где местная многотуберкулезная фауна серьезно пострадала в результате мел-палеогенового вымирания и так и не восстановилась полностью, в отличие от своих североамериканских и европейских родственников. В образовавшемся экологическом вакууме грызуны и другие виды Glires смогли эволюционировать и диверсифицироваться, занимая ниши, оставленные вымершими мультитуберкулезными видами. Корреляция между распространением грызунов и гибелью мультитуберкулезных микроорганизмов является спорной темой, до конца не решенной. Разнообразие американских и европейских мультитуберкулезных сообществ действительно снижается по мере появления грызунов в этих областях, но оставшиеся азиатские мультитуберкулезные сообщества сосуществовали с грызунами без видимой замены, и в конечном итоге обе клады сосуществовали по крайней мере 15 миллионов лет. . [91]
История колонизации континентов мира грызунами сложна. Передвижения большого надсемейства Muroidea (включая хомяков , песчанок , настоящих мышей и крыс ) могли включать до семи колонизаций Африки, пяти Северной Америки, четырех Юго-Восточной Азии, двух Южной Америки и до десяти Евразии. [92]

В эоцене грызуны начали диверсифицироваться. Бобры появились в Евразии в позднем эоцене, а в позднем миоцене распространились в Северную Америку. [94] В конце эоцена гистрикогнаты вторглись в Африку, скорее всего, зародившись в Азии не менее 39,5 миллионов лет назад. [95] Ископаемые данные из Африки показывают, что некоторые гистрикогнаты ( кавиоморфы ) колонизировали Южную Америку , которая в то время была изолированным континентом, очевидно, используя океанские течения, чтобы пересечь Атлантику на плавучих обломках . [96] Кавиоморфы прибыли в Южную Америку 41 миллион лет назад (подразумевается, что появление гистрикогнатов в Африке произошло как минимум еще раньше). [95] и достигли Больших Антильских островов к раннему олигоцену , что позволяет предположить, что они, должно быть, быстро рассеялись по Южной Америке. [97]
несомидные грызуны переправились из Африки на Мадагаскар 20–24 миллиона лет назад. Считается, что [98] Все 27 видов местных малагасийских грызунов , по-видимому, являются потомками одной колонизации.
К 20 миллионам лет назад появились окаменелости, явно принадлежащие к нынешним семействам, таким как Muridae. [86] К миоцену , когда Африка столкнулась с Азией, африканские грызуны, такие как дикобраз, начали распространяться в Евразию . [99] Некоторые ископаемые виды были очень большими по сравнению с современными грызунами, включая гигантского бобра Castoroides ohioensis , который вырастал до 2,5 м в длину (8 футов 2 дюйма) и веса 100 кг (220 фунтов). [100] Самым крупным известным грызуном была Josephoartigasia monesi , пакарана с предполагаемой длиной тела 3 м (10 футов). [101]
Первые грызуны прибыли в Австралию через Индонезию около 5 миллионов лет назад. Хотя сумчатые являются наиболее известными млекопитающими в Австралии, многие грызуны , принадлежащие к подсемейству Murinae , входят в число видов млекопитающих континента . [102] Существует около пятидесяти видов «старых эндемиков», первая волна грызунов, колонизировавших страну в миоцене и раннем плиоцене , и восемь настоящих видов крыс ( Rattus ) «новых эндемиков», прибывших в последующей волне в позднем плиоцене. или ранний плейстоцен . Максимальный возраст самых ранних ископаемых грызунов в Австралии составляет 4,5 миллиона лет. [103] а молекулярные данные согласуются с колонизацией Новой Гвинеи с запада в конце миоцена или начале плиоцена с последующей быстрой диверсификацией. Следующая волна адаптивной радиации произошла после одной или нескольких колонизаций Австралии примерно 2–3 миллиона лет спустя. [104]
Грызуны участвовали в Великом американском обмене , который произошел в результате объединения Америк путем образования Панамского перешейка около 3 миллионов лет назад в эпоху Пьяченца . [105] В ходе этого обмена небольшое количество видов , таких как дикобразы Нового Света (Erethizontidae), направились на север. [86] Однако основное вторжение сигмодонтинов на юг предшествовало формированию сухопутного моста по крайней мере на несколько миллионов лет, вероятно, произошло в результате рафтинга. [106] [107] [108] Сигмодонтины резко диверсифицировались однажды в Южной Америке, хотя некоторая степень диверсификации, возможно, уже произошла в Центральной Америке до колонизации. [107] [108]
Стандартная классификация [ править ]
Использование названия отряда «Rodentia» приписывается английскому путешественнику и натуралисту Томасу Эдварду Боудичу (1821 г.). [109] Современное латинское слово Rodentia происходит от слова Rodens , причастия настоящего времени от Rodere – «грызть», «разъедать». [110] Зайцы . , кролики и пищухи (отряд зайцеобразных) имеют, как и грызуны, непрерывно растущие резцы и одно время были включены в отряд Однако у них есть дополнительная пара резцов на верхней челюсти, и эти два отряда имеют совершенно разные эволюционные истории. [111] Филогения грызунов относит их к кладам Glires, Euarchontoglires и Boreoeutheria . Кладограмма ниже показывает внутренние и внешние отношения Rodentia на основе попытки Wu et al., предпринятой в 2012 году. чтобы привести молекулярные часы в соответствие с палеонтологическими данными: [112]
| Бореоэтерия |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



Живые семейства грызунов на основе исследования, проведенного Fabre et al. 2012. [113]
| Классификация грызунов |
|---|



Отряд Rodentia можно разделить на подотряды , инфраотряды , надсемейства и семейства . Среди грызунов существует значительная степень параллелизма и конвергенции, вызванная тем фактом, что они имели тенденцию эволюционировать, чтобы заполнить во многом схожие ниши. Эта параллельная эволюция включает не только структуру зубов, но и подглазничную область черепа (ниже глазницы) и затрудняет классификацию, поскольку сходные черты могут не быть связаны с общим происхождением. [114] [115] Брандт (1855) первым предложил разделить Rodentia на три подотряда: Sciuromorpha, Hystricomorpha и Myomorpha, в зависимости от развития определенных мышц челюсти, и эта система получила широкое распространение. Шлоссер (1884) провел всесторонний обзор окаменелостей грызунов, в основном используя щечные зубы, и обнаружил, что они вписываются в классическую систему, но Туллборг (1899) предложил только два подотряда: Sciurognathi и Hystricognathi. Они основывались на степени изгиба нижней челюсти и должны были быть далее подразделены на Sciuromorpha, Myomorpha, Hystricomorpha и Bathyergomorpha. Мэтью (1910) создал филогенетическое древо грызунов Нового Света, но не включил в него наиболее проблемные виды Старого Света. Дальнейшие попытки классификации продолжались без согласия: некоторые авторы приняли классическую систему трех подотрядов, а другие - два подотряда Таллборга. [114]
Эти разногласия остаются нерешенными, а молекулярные исследования не разрешили ситуацию полностью, хотя и подтвердили монофилию группы и то, что клада произошла от общего палеоценового предка. Карлтон и Массер (2005) в книге «Виды млекопитающих мира» предварительно приняли систему из пяти подотрядов: Sciuromorpha, Castorimorpha, Myomorpha, Anomaluromorpha и Hystricomorpha. По состоянию на 2021 год Американское общество маммологов признает 34 современных семейства, содержащих более 481 рода и 2277 видов. [116] [117] [118]

Отряд Rodentia (от лат. Rodere — грызть)





- Подотряд Аномалуроморфы
- Семейство Anomaluridae : чешуйчатохвостые белки.
- Семейство Pedetidae : весенние зайцы.
- Семейство Zenkerellidae : камерунские чешуйчатохвосты.
- Подотряд Castorimorpha
- Надсемейство Castoroidea.
- Семейство Castoridae : бобры.
- Надсемейство Geomyoidea.
- Семейство Geomyidae : карманные суслики (настоящие суслики).
- Семейство Heteromyidae : кенгуровые крысы, кенгуровые мыши.
- Надсемейство Castoroidea.
- Подотряд Hystricomorpha
- Инфраотряд Ctenodactylomorphi
- Семейство Ctenodactylidae : Gundis.
- Семейство Diatomyidae : лаосская каменная крыса.
- Инфраотряд Гистрикогнати
- Парвордер Фиоморфа
- Семейство Bathyergidae : африканские землекопы.
- Семейство Heterocephalidae : голый землекоп.
- Семейство Hystricidae : дикобразы Старого Света.
- Семейство Petromuridae : барсучьи крысы.
- Семейство Thryonomyidae : тростниковые крысы.
- Пардер Кавиоморфа
- Надсемейство Erethizontoidea.
- Семейство Erethizontidae : дикобразы Нового Света.
- Надсемейство Chinchilloidea.
- Семейство Chinchillidae : шиншиллы , вискаши.
- Семейство Dinomyidae : караканы.
- Надсемейство Cavioidea.
- Семейство Caviidae : морские свинки , включая морских свинок и капибар.
- Семейство Dasyproctidae : агути.
- Семейство Cuniculidae : мирные птицы.
- Надсемейство Octodontoidea.
- Семейство Abrocomidae : шиншилловые крысы.
- Семейство Ctenomyidae : туко-тукос.
- Семейство Echimyidae : колючие крысы, хутии и нутрии.
- Семейство Octodontidae : октодонты.
- Надсемейство Erethizontoidea.
- Инфраотряд Ctenodactylomorphi
- Подотряд Миоморфа
- Надсемейство Dipodoidea.
- Семейство Dipodidae : тушканчики.
- Семейство Sminthidae : березовые мыши.
- Семейство Zapodidae : прыгающие мыши.
- Надсемейство Муроидеа
- Семейство Calomyscidae : мышевидные хомяки.
- Семейство Cricetidae : хомяки , крысы и мыши Нового Света , ондатры , полевки , лемминги .
- Семейство Muridae : настоящие мыши и крысы, песчанки , колючие мыши, хохлатая крыса.
- Семейство Nesomyidae : лазающие мыши, каменные мыши, белохвостые крысы, малагасийские крысы и мыши.
- Семейство Platacanthomyidae : колючая соня.
- Семейство Spalacidae : слепыши, бамбуковые крысы, цокоры.
- Надсемейство Dipodoidea.
- Подотряд Sciuromorpha
- Семейство Aplodontiidae : горный бобр.
- Семейство Gliridae (также Myoxydae, Muscardinidae): сони.
- Семейство Sciuridae : белки, в том числе бурундуки , луговые собачки , сурки.
Взаимодействие с людьми [ править ]
Сохранение [ править ]

Хотя грызуны не относятся к отряду млекопитающих, находящихся под наибольшей угрозой, существует 168 видов 126 родов, которые, как говорят, требуют природоохранного внимания. [119] в условиях ограниченного признания со стороны общественности. Поскольку 76 процентов родов грызунов содержат только один вид, значительная часть филогенетического разнообразия может быть утрачена при сравнительно небольшом количестве вымираний. В отсутствие более подробных знаний о видах, находящихся под угрозой, и точной таксономии, сохранение должно основываться главным образом на более высоких таксонах (таких как семейства, а не виды) и географических горячих точках. [119] Несколько видов рисовых крыс вымерли с 19 века, вероятно, из-за утраты среды обитания и внедрения чужеродных видов. [120] В Колумбии бурый волосатый карликовый дикобраз был зарегистрирован только в двух горных местонахождениях в 1920-е годы, тогда как краснохохлатая мягкошерстная колючая крыса известна только из своего типового местонахождения на побережье Карибского моря, поэтому эти виды считаются уязвимыми. [121] Комиссия МСОП по выживанию видов пишет: «Мы можем с уверенностью заключить, что многие южноамериканские грызуны находятся под серьезной угрозой, в основном из-за нарушения окружающей среды и интенсивной охоты». [122]
«Три ныне космополитических вида комменсальных грызунов-вредителей» [123] (коричневая крыса, черная крыса и домовая мышь) были расселены вместе с людьми, частично на парусных кораблях в эпоху географических открытий , а четвертый вид в Тихом океане, полинезийская крыса ( Rattus exulans ), серьезно пострадала от повреждена островная биота по всему миру. Например, когда черная крыса достигла острова Лорд-Хау в 1918 году, более 40 процентов видов наземных птиц острова, включая трубастого хвоста Лорд-Хау , [124] вымерли в течение десяти лет. Подобные разрушения наблюдались на острове Мидуэй (1943 г.) и острове Биг-Саут-Кейп (1962 г.). Природоохранные проекты могут при тщательном планировании полностью искоренить этих грызунов-вредителей на островах с помощью антикоагулянтного родентицида, такого как бродифакум . [123] Этот подход оказался успешным на острове Ланди в Соединенном Королевстве, где уничтожение примерно 40 000 коричневых крыс дает популяциям мэнских буревестников и атлантических тупиков шанс восстановиться после почти полного исчезновения. [125] [126]
Грызуны также оказались восприимчивы к изменению климата , особенно виды, живущие на низменных островах. Меломис Брэмбл -Кей , обитавший в самой северной точке суши Австралии , был первым видом млекопитающих, объявленным вымершим в результате антропогенного изменения климата . [127]
Эксплуатация [ править ]
Мех [ править ]

Человечество издавна использовало шкуры животных для одежды, так как кожа долговечна, а мех обеспечивает дополнительную изоляцию. [3] Коренные жители Северной Америки широко использовали бобровые шкуры, дублили и шили их для изготовления халатов. Европейцы оценили их качество, и торговля мехом в Северной Америке стала развиваться и стала иметь первостепенное значение для первых поселенцев. В Европе мягкий подшерсток, известный как «бобровая шерсть», оказался идеальным для валяния, из него делали бобровые шапки и отделку одежды. [128] [129] Позже нутрия стала более дешевым источником меха для валяния и широко выращивалась в Америке и Европе; однако мода изменилась, стали доступны новые материалы, и эта область меховой промышленности пришла в упадок. [130] Шиншилла имеет мягкую и шелковистую шерсть, а спрос на ее мех был настолько высок, что он был почти полностью истреблен в дикой природе , прежде чем сельское хозяйство стало основным источником шкур. [130] Иглы и остевые волосы дикобразов используются для изготовления традиционной декоративной одежды. Например, их остевые волосы используются при создании головного убора коренных американцев «свиная плотва» . Основные иглы можно покрасить, а затем использовать в сочетании с нитками для украшения кожаных аксессуаров, таких как ножны для ножей и кожаные сумки. Женщины лакота собирали иглы для работы с перьями , набрасывая одеяло на дикобраза и забирая иглы, застрявшие в одеяле. [131]
Потребление [ править ]
Человек употребляет в пищу по меньшей мере 89 видов грызунов, в основном Hystricomorpha, таких как морские свинки, агути и капибары; в 1985 году существовало как минимум 42 различных общества, в которых люди ели крыс. [132] Морские свинки впервые были выращены в пищу около 2500 г. до н.э., а к 1500 г. до н.э. они стали основным источником мяса для Империи инков . Римляне выращивали сонь в специальных горшках, называемых «глирария», или в больших открытых вольерах, где их откармливали грецкими орехами, каштанами и желудями. Сонь также ловили в дикой природе осенью, когда они были самыми жирными, и их либо жарили и обмакивали в мед, либо запекали с начинкой из смеси свинины, кедровых орехов и других ароматизаторов. Исследователи обнаружили, что в Амазонии, где крупных млекопитающих было мало, пака и обыкновенные агути составляли около 40 процентов ежегодной добычи, добываемой коренными жителями, но в лесных районах, где было много крупных млекопитающих, эти грызуны составляли лишь около 3 процентов от общего количества дичи. брать. [132]
Морские свинки используются в кухне Куско , Перу, в таких блюдах, как cuy al Horno , запеченная морская свинка. [3] [133] Традиционная андская печь, известная как конча или фогон , сделана из глины и глины, укрепленной соломой и волосами животных, таких как морские свинки. [134] В Перу в любое время насчитывается 20 миллионов домашних морских свинок, которые ежегодно производят 64 миллиона съедобных туш. Это животное является отличным источником пищи, поскольку его мясо на 19% состоит из белка. [132] В Соединенных Штатах в основном белок, а также ондатр, дикобразов и сурков люди едят . Народ навахо ел луговых собачек, запеченных в грязи, а пайуте ели сусликов, белок и крыс. [132]
Тестирование на животных [ править ]

Грызуны широко используются в качестве модельных организмов при испытаниях на животных. [3] [135] -мутанты-альбиносы Крысы впервые были использованы для исследований в 1828 году и позднее стали первыми животными, одомашненными в чисто научных целях. [136] В настоящее время домовая мышь является наиболее часто используемым лабораторным грызуном, и в 1979 году было подсчитано, что во всем мире ежегодно используется пятьдесят миллионов таких грызунов. Им отдают предпочтение из-за их небольшого размера, плодовитости, короткого периода беременности и простоты обращения, а также из-за того, что они восприимчивы ко многим заболеваниям и инфекциям, поражающим людей. Они используются в исследованиях в области генетики , биологии развития , клеточной биологии , онкологии и иммунологии . [137] Морские свинки были популярными лабораторными животными до конца 20 века; в 1960-х годах в США для исследований ежегодно использовалось около 2,5 миллионов морских свинок. [138] но к середине 1990-х годов это общее количество уменьшилось примерно до 375 000. [139] В 2007 году они составляли около 2% всех лабораторных животных. [138] Морские свинки сыграли важную роль в создании теории микробов в конце 19 века благодаря экспериментам Луи Пастера , Эмиля Ру и Роберта Коха . [140] Их несколько раз запускали в орбитальный космический полет - сначала СССР на «Спутник-9» биоспутнике 9 марта 1961 года с успешным возвращением. [141] Голый землекоп — единственное известное пойкилотермное млекопитающее ; его используют в исследованиях по терморегуляции . Необычно также отсутствие выработки нейромедиатора вещества P , и этот факт исследователи находят полезным при изучении боли . [142]
Грызуны обладают чувствительными обонятельными способностями, которые используются людьми для обнаружения запахов или интересующих химических веществ. [143] Гамбийская сумчатая крыса способна выявлять туберкулезные палочки с чувствительностью до 86,6% и специфичностью (обнаружение отсутствия бацилл) более 93%; тот же вид был обучен обнаруживать мины . [144] [145] Крыс изучали на предмет возможного использования в опасных ситуациях, например, в зонах стихийных бедствий. Их можно научить реагировать на команды, которые можно подавать удаленно, и даже убедить выйти в ярко освещенные места, которых крысы обычно избегают. [146] [147] [148]
В качестве домашних животных [ править ]

Грызуны, включая морских свинок, [149] мыши, крысы, хомяки, песчанки, шиншиллы, дегу и бурундуки — удобные домашние животные, способные жить в небольших помещениях, причем каждый вид обладает своими качествами. [150] Большинство из них обычно содержатся в клетках подходящего размера и имеют различные требования к пространству и социальному взаимодействию. Если обращаться с ними с раннего возраста, они обычно послушны и не кусаются. Морские свинки долго живут и нуждаются в большой клетке. [75] Крысам также нужно много места, и они могут стать очень ручными, могут научиться трюкам и, кажется, наслаждаются человеческим обществом. Мыши недолговечны, но занимают очень мало места. Хомяки ведут одиночный образ жизни, но, как правило, ведут ночной образ жизни. У них интересное поведение, но если с ними не обращаться регулярно, они могут занять оборонительную позицию. Песчанки обычно не агрессивны, редко кусаются и являются общительными животными, которым нравится компания людей и себе подобных. [151]
вредители и Как болезней переносчики

Некоторые виды грызунов являются серьезными сельскохозяйственными вредителями , поедая большое количество продуктов питания, хранящихся человеком. [152] Например, в 2003 году количество риса , потерянного из-за мышей и крыс в Азии, оценивалось в достаточном количестве, чтобы накормить 200 миллионов человек. Большая часть ущерба во всем мире наносится относительно небольшим количеством видов, в основном крысами и мышами. [153] В Индонезии и Танзании грызуны снижают урожайность примерно на пятнадцать процентов, а в некоторых случаях в Южной Америке потери достигают девяноста процентов. По всей Африке грызуны, включая Mastomys и Arvicanthis, повреждают зерновые, арахис, овощи и какао. В Азии крысы, мыши и такие виды, как Microtus Brandti , Meriones unguiculatus и Eospalax Baileyi, повреждают посевы риса, сорго , клубней, овощей и орехов. В Европе, наряду с крысами и мышами, виды Apodemus , Microtus и в отдельных вспышках Arvicola terrestris наносят ущерб садам, овощам и пастбищам, а также зерновым культурам. В Южной Америке более широкий спектр видов грызунов, таких как Holochilus , Akodon , Calomys , Oligoryzomys , Phyllotis , Sigmodon и Zygodontomys , повреждают многие сельскохозяйственные культуры, включая сахарный тростник, фрукты, овощи и клубнеплоды. [153]
Грызуны также являются важными переносчиками болезней. [154] Черная крыса с переносимыми ею блохами играет основную роль в распространении бактерии Yersinia pestis, вызывающей бубонную чуму . [155] и переносит организмы, ответственные за сыпной тиф , болезнь Вейля , токсоплазмоз и трихинеллез . [154] Ряд грызунов являются переносчиками хантавирусов , в том числе вирусов Пуумала , Добрава и Сааремаа , которые могут заразить человека. [156] Грызуны также способствуют переносу таких заболеваний, как бабезиоз , кожный лейшманиоз , гранулоцитарный анаплазмоз человека , болезнь Лайма , омская геморрагическая лихорадка , вирус Повассан , риккетсиоз , возвратный тиф , пятнистая лихорадка Скалистых гор и вирус Западного Нила . [157]

Поскольку грызуны доставляют неудобства и угрожают общественному здоровью , человеческое общество часто пытается их контролировать. Традиционно это включало отравление и отлов в ловушки — методы, которые не всегда были безопасными и эффективными. В последнее время комплексная борьба с вредителями пытается улучшить контроль за счет сочетания обследований для определения размера и распределения популяции вредителей, установления пределов толерантности (уровней активности вредителей, при которых необходимо вмешательство), вмешательств и оценки эффективности на основе повторные опросы. Вмешательства могут включать просвещение, принятие и применение законов и постановлений, изменение среды обитания, изменение методов ведения сельского хозяйства и биологический контроль с использованием патогенов или хищников , а также отравление и отлов. [158] Использование таких патогенов, как сальмонелла, имеет тот недостаток, что они могут заразить человека и домашних животных, а грызуны часто становятся устойчивыми. Использование хищников, включая хорьков , мангустов и варанов, было признано неудовлетворительным. Домашние и дикие кошки способны эффективно бороться с грызунами, если их популяция не слишком велика. [159] В Великобритании, в частности, два вида, домовая мышь и коричневая крыса, активно контролируются, чтобы ограничить ущерб выращиваемым сельскохозяйственным культурам, потери и загрязнение хранящихся культур и структурный ущерб объектам, а также для соблюдения закона. [160]
См. также [ править ]
- Фе, Фай, Фо, Фум и Фуи — мыши, которые вращались вокруг Луны на Аполлоне-17.
- Мышиные модели метастазов рака молочной железы
Ссылки [ править ]
- ^ Смит, Эндрю Т. «Зайцеморф» . Британская энциклопедия . Проверено 31 мая 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час Сингл, Г.; Дикман, ЧР; Макдональд, Д.В. (2001). «Грызуны». В Макдональде, Д.В. (ред.). Энциклопедия млекопитающих (2-е изд.). Издательство Оксфордского университета. стр. 578–587. ISBN 978-0-7607-1969-5 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Ваггонер, Бен (15 августа 2000 г.). «Знакомство с грызунами» . Музей палеонтологии Калифорнийского университета. Архивировано из оригинала 29 апреля 2009 года . Проверено 4 июля 2014 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Новак, Р.М. (1999). Млекопитающие мира Уокера . Издательство Университета Джонса Хопкинса . п. 1244. ИСБН 978-0-8018-5789-8 .
- ^ Ниемец, Брук А. (15 октября 2011 г.). Заболевания зубов, полости рта и челюстно-лицевой области мелких животных: цветной справочник . ЦРК Пресс. п. 13. ISBN 978-1-84076-630-1 .
- ^ Кокс, Филип Г.; Рэйфилд, Эмили Дж.; Фэган, Майкл Дж.; Херрел, Энтони; Патаки, Тодд С.; Джеффри, Натан (27 апреля 2012 г.). «Функциональная эволюция системы питания грызунов» . ПЛОС ОДИН . 7 (4): e36299. Бибкод : 2012PLoSO...736299C . дои : 10.1371/journal.pone.0036299 . ISSN 1932-6203 . ПМЦ 3338682 . ПМИД 22558427 .
- ^ Тернбулл, Уильям Д. (1970). Жевательный аппарат млекопитающих . Том. 18. [Чикаго]: Издательство Филд Музея. Архивировано из оригинала 29 апреля 2022 года . Проверено 11 мая 2022 г.
- ↑ Перейти обратно: Перейти обратно: а б Фроберг-Фейко, Карен (1 октября 2014 г.). «Дайте крысе кость: удовлетворение потребности грызунов грызть» . Лабораторное животное . 43 (10): 378–379. дои : 10.1038/laban.611 . ISSN 1548-4475 . S2CID 19686731 .
- ^ Кокс, Филип Г.; Джеффри, Натан (2011). «Обзор морфологии мускулатуры, закрывающей челюсти, у белок, крыс и морских свинок с помощью микроКТ с контрастным усилением» (PDF) . Анатомическая запись . 294 (6): 915–928. дои : 10.1002/ar.21381 . ПМИД 21538924 . S2CID 17249666 . Архивировано (PDF) из оригинала 28 октября 2020 г. Проверено 26 августа 2020 г.
- ^ Дакетт, В. (1853). «защечный мешочек». Разговор на английском языке и чтение . Эд Мишель Леви. п. 3. Архивировано из оригинала 5 марта 2023 года . Проверено 1 ноября 2020 г.
- ^ Мустафа, О. (2015). «Морфология ротовой полости африканской гигантской крысы» . Болгарский журнал ветеринарной медицины . 18 (1): 19–30. дои : 10.15547/bjvm.793 .
- ^ Торингтон, Р.В. младший; Дэрроу, К.; Андерсон, CG (1998). «Анатомия законцовки крыла и аэродинамика летяг» (PDF) . Журнал маммологии . 79 (1): 245–250. дои : 10.2307/1382860 . JSTOR 1382860 . Архивировано (PDF) из оригинала 9 апреля 2009 г. Проверено 26 августа 2014 г.
- ^ «Шиншилла» . Зоопарк Элмвуд Парк . Архивировано из оригинала 30 декабря 2021 года . Проверено 30 декабря 2021 г.
- ↑ Перейти обратно: Перейти обратно: а б с Петерсен, Карл CH (8 июля 2014 г.). «Корковый контроль движения усов» . Ежегодный обзор неврологии . 37 (1). Годовые обзоры : 183–203. doi : 10.1146/annurev-neuro-062012-170344 . ISSN 0147-006X . ПМИД 24821429 .
- ^ Хорн, Чарльз К.; Кимбалл, Брюс А.; Ван, Хун; Каус, Джеймс; Динель, Сэмюэл; Надь, Аллиза; Гатрайт, Гордон Р.; Йейтс, Билл Дж.; Эндрюс, Пол ЛР (10 апреля 2013 г.). Коваса, Михай (ред.). «Почему грызунов не может рвать? Сравнительное поведенческое, анатомическое и физиологическое исследование» . ПЛОС ОДИН . 8 (4): е60537. Бибкод : 2013PLoSO...860537H . дои : 10.1371/journal.pone.0060537 . ISSN 1932-6203 . ПМЦ 3622671 . ПМИД 23593236 .
- ^ Капур, Харит; Лохани, Куш Радж; Ли, Томми Х.; Агравал, Девендра К.; Миттал, Сумит К. (27 июля 2015 г.). «Животные модели пищевода Барретта и аденокарциномы пищевода — прошлое, настоящее и будущее» . Клиническая и трансляционная наука . 8 (6): 841–847. дои : 10.1111/cts.12304 . ПМЦ 4703452 . ПМИД 26211420 .
- ^ Балабан, Кэри Д.; Йейтс, Билл Дж. (2017). «Что такое тошнота? Исторический анализ меняющихся взглядов» . Автономная нейронаука . 202 : 5–17. дои : 10.1016/j.autneu.2016.07.003 . ISSN 1566-0702 . ПМК 5203950 . ПМИД 27450627 .
- ^ Шир, Линдси А.; Спектор, Алан К. (1 января 2019 г.). «Функциональные и нейробиологические свойства дурного вкуса» . Физиологические обзоры . 99 (1): 605–663. doi : 10.1152/physrev.00044.2017 . ISSN 0031-9333 . ПМК 6442928 . ПМИД 30475657 .
- ^ Хорн, Чарльз К.; Арделл, Джеффри Л.; Фишер, Ли Э. (1 марта 2019 г.). «Электротерапевтическое воздействие на вегетативную нервную систему» . Физиология . 34 (2): 150–162. дои : 10.1152/физиол.00030.2018 . ISSN 1548-9213 . ПМК 6586833 . ПМИД 30724129 .
- ^ Шульте-Хостедде, AI (2008). «Глава 10: Половой диморфизм размеров у грызунов». В Вольфе, Джерри О.; Шерман, Пол В. (ред.). Общества грызунов: экологическая и эволюционная перспектива . Издательство Чикагского университета. стр. 117–119. ISBN 978-0-226-90538-9 .
- ^ Хелген, Кристофер М. (2005). «Мыши-земноводные Новой Гвинеи (Rodentia, Muridae): родовой статус Baiyankamys и описание нового вида Hydromys ». Зоотакса . 913 : 1–20. дои : 10.11646/zootaxa.913.1.1 . ISSN 1175-5326 .
- ^ Паршад, В.Р. (1999). «Борьба с грызунами в Индии» (PDF) . Комплексные обзоры борьбы с вредителями . 4 (2): 97–126. дои : 10.1023/А:1009622109901 . S2CID 36804001 . Архивировано (PDF) из оригинала 4 сентября 2014 года . Проверено 30 августа 2014 г.
- ↑ Перейти обратно: Перейти обратно: а б Янке, Аксель; Мартинес-Эстевес, Лурдес; Бальванера, Патрисия; Пачеко, Хесус; Себальос, Херардо (2013). «Уменьшение численности луговых собачек сокращает предоставление экосистемных услуг и приводит к опустыниванию полузасушливых лугов» . ПЛОС ОДИН . 8 (10): е75229. Бибкод : 2013PLoSO...875229M . дои : 10.1371/journal.pone.0075229 . ISSN 1932-6203 . ПМК 3793983 . ПМИД 24130691 .
- ^ Крюгер, Кирстен (1986). «Пищевые отношения между бизонами, вилорогами и луговыми собачками: экспериментальный анализ». Экология . 67 (3): 760–770. Бибкод : 1986Экол...67..760К . дои : 10.2307/1937699 . ISSN 0012-9658 . JSTOR 1937699 .
- ^ Перес, Франциско; Кастильо-Гевара, Читлалли; Галиндо-Флорес, Хема; Куаутле, Мариана; Эстрада-Торрес, Артуро (2012). «Влияние прохождения кишечника двумя высокогорными грызунами на активность спор и образование микоризы двух видов эктомикоризных грибов ( Laccaria trichodermophora и Suillus tomentosus )». Ботаника . 90 (11): 1084–1092. дои : 10.1139/b2012-086 . ISSN 1916-2790 .
- ^ Берчстед, Д.; Дэниелс, М.; Торсон, Р.; Вокоун, Дж. (2010). «Дисконкум реки: применение бобровых модификаций к базовым условиям для восстановления лесных верховьев» . Бионаука . 60 (11): 908–922. дои : 10.1525/bio.2010.60.11.7 . S2CID 10070184 . Архивировано из оригинала 5 августа 2020 года . Проверено 7 декабря 2018 г.
- ^ Райт, JP; Джонс, CG; Флекер, А.С. (2002). «Инженер экосистемы, бобр, увеличивает видовое богатство в масштабе ландшафта» (PDF) . Экология . 132 (1): 96–101. Бибкод : 2002Oecol.132...96W . дои : 10.1007/s00442-002-0929-1 . ПМИД 28547281 . S2CID 5940275 . Архивировано (PDF) из оригинала 20 февраля 2014 года . Проверено 4 июля 2014 г.
- ^ Кемп, PS; Уортингтон, штат Техас; Лэнгфорд, TE l.; Дерево, ARJ; Гейвуд, MJ (2012). «Качественное и количественное воздействие реинтродуцированных бобров на речную рыбу». Рыба и рыболовство . 13 (2): 158–181. Бибкод : 2012AqFF...13..158K . дои : 10.1111/j.1467-2979.2011.00421.x .
- ^ «Грызуны… Вредители… Вредители… Какая разница?» . Получите решения Smart Rat . 1 апреля 2020 года. Архивировано из оригинала 30 декабря 2021 года . Проверено 30 декабря 2021 г.
- ^ Ханссон, Леннарт (1971). «Динамика среды обитания, питания и популяции полевки Microtus agrestis (L.) на юге Швеции» . Вильтреви . 8 : 268–278. ISSN 0505-611X . Архивировано из оригинала 27 сентября 2013 года.
- ^ Конниор, МБ (2011). « Geomys bursarius (Rodentia: Geomyidae)» . Виды млекопитающих . 43 (1): 104–117. дои : 10.1644/879.1 .
- ^ «Техасский карманный суслик» . Млекопитающие Техаса: Грызуны . NSRL: Музей Техасского технологического университета. Архивировано из оригинала 5 октября 2018 года . Проверено 4 июля 2014 г.
- ↑ Перейти обратно: Перейти обратно: а б с Аттенборо, Дэвид (2002). Жизнь млекопитающих . Книги Би-би-си. стр. 61–86. ISBN 978-0-563-53423-5 .
- ^ Мюллер-Шварце, Дитланд; Сунь, Лисинг (2003). Бобер: естественная история инженера по водно-болотным угодьям . Издательство Корнельского университета. стр. 67–75. ISBN 978-0-8014-4098-4 . Архивировано из оригинала 5 марта 2023 года . Проверено 27 января 2016 г.
- ↑ Перейти обратно: Перейти обратно: а б Лэндри, Стюарт О. младший (1970). «Грызуны как всеядные». Ежеквартальный обзор биологии . 45 (4): 351–372. дои : 10.1086/406647 . JSTOR 2821009 . ПМИД 5500524 . S2CID 30382320 .
- ^ « Hydromys chrysogaster : Водяная крыса» . Вода для здоровой страны . ЦСИРО. 30 июня 2004 г. Архивировано из оригинала 21 мая 2014 г. . Проверено 4 июля 2014 г.
- ^ «Северная мышь-кузнечик» . Млекопитающие Техаса: Грызуны . NSRL: Музей Техасского технологического университета. Архивировано из оригинала 21 октября 2020 года . Проверено 4 июля 2014 г.
- ↑ Перейти обратно: Перейти обратно: а б с Джарвис, Дженнифер (1981). «Эусоциальность млекопитающих: совместное размножение в колониях голых землекопов». Наука . 212 (4494): 571–573. Бибкод : 1981Sci...212..571J . дои : 10.1126/science.7209555 . JSTOR 1686202 . ПМИД 7209555 .
- ↑ Перейти обратно: Перейти обратно: а б Хугланд, Джон Л. (1995). Чернохвостая луговая собачка: общественная жизнь роющего млекопитающего . Издательство Чикагского университета. п. 1. ISBN 978-0-226-35118-6 .
- ^ Воган, Т.А. (1962). «Репродукция равнинного карманного суслика в Колорадо». Журнал маммологии . 43 (1): 1–13. дои : 10.2307/1376874 . JSTOR 1376874 .
- ^ Бейкер, Брюс В.; Хилл, Эдвард П. (2003). «Глава 15: Бобер» . В Фельдхамере, Джордж А.; Томпсон, Брюс С.; Чепмен, Джозеф А. (ред.). Дикие млекопитающие Северной Америки: биология, управление и охрана . Джу Пресс. стр. 288–310. ISBN 978-0-8018-7416-1 .
- ^ Хэнсон, Энн (25 октября 2006 г.). «Поведение дикой норвежской крысы» . Поведение и биология крыс . Архивировано из оригинала 1 октября 2014 года . Проверено 1 июля 2014 г.
- ^ Уинслоу, Джеймс Т.; Гастингс, Ник; Картер, К. Сью; Харбо, Кэрролл Р.; Инсел, Томас Р. (1993). «Роль центрального вазопрессина в образовании пар у моногамных степных полевок» (PDF) . Письма к природе . 365 (6446): 545–548. Бибкод : 1993Natur.365..545W . дои : 10.1038/365545a0 . ПМИД 8413608 . S2CID 4333114 . Архивировано из оригинала (PDF) 14 июля 2014 года.
- ^ Йенсен, Эрик; Шерман, Пол В. (2003). «Глава 10: Суслики» . В Фельдхамере, Джордж А.; Томпсон, Брюс С.; Чепмен, Джозеф А. (ред.). Дикие млекопитающие Северной Америки: биология, управление и охрана . Джу Пресс. стр. 211–225. ISBN 978-0-8018-7416-1 .
- ^ Беннетт, Северная Каролина; Джарвис, ДЖУМ (2004). « Криптомис дамаренсис » . Виды млекопитающих . 756 : Номер 756: стр. 1–5. дои : 10.1644/756 .
- ^ Аракава, Хироюки; Бланшар, Д. Кэролайн; Аракава, Кейко; Данлэп, Кристофер; Бланшар, Роберт Дж. (2008). «Поведение ароматической маркировки как запаховая коммуникация у мышей» . Неврологические и биоповеденческие обзоры . 32 (7): 1236–1248. doi : 10.1016/j.neubiorev.2008.05.012 . ПМЦ 2577770 . ПМИД 18565582 .
- ^ Холмс, Уоррен Г.; Матео, Джилл М. (2008). «Глава 19: Распознавание родства у грызунов: проблемы и доказательства» . В Вольфе, Джерри О.; Шерман, Пол В. (ред.). Общества грызунов: экологическая и эволюционная перспектива . Издательство Чикагского университета. стр. 216–230. ISBN 978-0-226-90538-9 .
- ↑ Перейти обратно: Перейти обратно: а б Шервин, CM (2002). «Комфортные помещения для мышей в научно-исследовательских учреждениях» . Виктор и Энни Рейнхардт (ред.). Комфортные помещения для лабораторных животных (9-е изд.). Институт защиты животных. Архивировано из оригинала 6 октября 2014 года.
- ^ Бьоркойли, Торе; Розелл, Фрэнк (2002). «Испытание феномена дорогого врага у евразийского бобра». Поведение животных . 63 (6): 1073–1078. дои : 10.1006/anbe.2002.3010 . HDL : 11250/2437993 . S2CID 53160345 .
- ^ Ваше, М.; Феррон, Дж.; Гуат, П. (2001). «Способность красных белок ( Tamiasciurus hudsonicus ) различать обонятельные признаки сородичей». Канадский журнал зоологии . 79 (7): 1296–1300. дои : 10.1139/z01-085 . S2CID 86280677 .
- ^ Шелли, Эрин Л.; Блюмштейн, Дэниел Т. (2005). «Эволюция голосовой тревожной коммуникации у грызунов». Поведенческая экология . 16 (1): 169–177. CiteSeerX 10.1.1.541.4408 . дои : 10.1093/beheco/arh148 .
- ^ Слободчиков, Китай; Пасека, Андреа; Вердолин, Дженнифер Л. (2009). «Тревожные сигналы луговых собачек кодируют метки о цветах хищников» (PDF) . Познание животных . 12 (3): 435–439. дои : 10.1007/s10071-008-0203-y . ПМИД 19116730 . S2CID 13178244 . Архивировано из оригинала (PDF) 7 октября 2018 года . Проверено 2 июля 2014 г.
- ^ Циммерманн, Эльке; Леливельд, Лизетт; Шешка, Лизетт (2013). «8: К эволюционным корням аффективной просодии в акустической коммуникации человека: сравнительный подход к голосам млекопитающих» . В Альтенмюллере, Эккарте; Шмидт, Сабина; Циммерманн, Эльке (ред.). Эволюция эмоционального общения: от звуков у млекопитающих к речи и музыке у человека . Издательство Оксфордского университета. стр. 123–124. ISBN 978-0-19-164489-4 .
- ^ Ванден Хоул, Шарлотта; Ван Даэле, Пол ААГ; Десмет, Нильс; Девос, Пол и Адрианс, Доминик (2014). «Подразумевает ли социальность сложную систему голосовой коммуникации? Пример Fukomys micklemi (Bathyergidae, Rodentia)». Биоакустика . 23 (2): 143–160. Бибкод : 2014Bioac..23..143В . дои : 10.1080/09524622.2013.841085 . S2CID 84503870 .
- ^ Лонг, резюме (2007). «Вокализации дегу ( Octodon degus ), социального грызуна-кавиоморфа». Биоакустика . 16 (3): 223–244. Бибкод : 2007Bioac..16..223L . дои : 10.1080/09524622.2007.9753579 . ISSN 0952-4622 . S2CID 84569309 .
- ^ Анчиллотто, Леонардо; Созио, Джулия; Мортеллити, Алессио; Руссо, Данило (2014). «Ультразвуковая коммуникация у Gliridae (Rodentia): орешниковая соня ( Muscardinus avellanarius ) на примере». Биоакустика . 23 (2): 129–141. Бибкод : 2014Bioac..23..129A . дои : 10.1080/09524622.2013.838146 . S2CID 84012458 .
- ^ Панксепп, Яак; Бургдорф, Джефф (2003). « Смеющиеся крысы и эволюционные предшественники человеческой радости?». Физиология и поведение . 79 (3): 533–547. CiteSeerX 10.1.1.326.9267 . дои : 10.1016/S0031-9384(03)00159-8 . ПМИД 12954448 . S2CID 14063615 .
- ^ Хаверкамп, Силке; Вассле, Хайнц; Дюбель, Йенс; Кунер, Томас; Августин, Джордж Дж.; Фэн, Гопин; Эйлер, Томас (2005). «Изначальная сине-конусная цветовая система сетчатки мыши» . Журнал неврологии . 25 (22): 5438–5445. doi : 10.1523/JNEUROSCI.1117-05.2005 . ПМК 6725002 . ПМИД 15930394 .
- ^ Хэнсон, Энн. «Что видят крысы?» . Поведение и биология крыс . Архивировано из оригинала 24 сентября 2014 года . Проверено 1 июля 2014 г.
- ^ Пикрелл, Джон (8 июля 2003 г.). «Мочевое зрение? Как грызуны общаются с ультрафиолетовым светом» . Национальные географические новости . Архивировано из оригинала 13 марта 2014 года . Проверено 8 июля 2014 г.
- ^ Дежарден, К.; Маруниак, Дж.А.; Бронсон, Ф.Х. (1973). «Социальный ранг домашних мышей: дифференциация, выявленная с помощью ультрафиолетовой визуализации моделей маркировки мочи». Наука . 182 (4115): 939–941. Бибкод : 1973Sci...182..939D . дои : 10.1126/science.182.4115.939 . ПМИД 4745598 . S2CID 44346136 .
- ^ Витала, Дж.; Корпимяки, Э.; Палокангас, П.; Койвула, М. (1995). «Привлечение пустельг к запахам полевок, видимым в ультрафиолетовом свете». Природа . 373 (6513): 425–427. Бибкод : 1995Natur.373..425V . дои : 10.1038/373425a0 . S2CID 4356193 .
- ↑ Перейти обратно: Перейти обратно: а б с «Вибрационная коммуникация у млекопитающих» . Карта жизни: конвергентная эволюция онлайн . Кембриджский университет. 4 августа 2010 года. Архивировано из оригинала 14 июля 2014 года . Проверено 5 июля 2014 г.
- ^ Рэндалл, Дж. А. (2001). «Эволюция и функция игры на барабанах как средство общения у млекопитающих» . Американский зоолог . 41 (5): 1143–1156. дои : 10.1093/icb/41.5.1143 .
- ^ Рэндалл, Ян А.; Маток, Марджори Д. (1997). «Почему крысы-кенгуру ( Dipodomys spectabilis ) барабанят по змеям?» . Поведенческая экология . 8 (4): 404–413. дои : 10.1093/beheco/8.4.404 .
- ^ Наринс, премьер-министр; Райхман, О.Дж.; Джарвис, JUM; Льюис, скорая помощь (1992). «Передача сейсмического сигнала между норами капского слепыша Georychus capensis ». Журнал сравнительной физиологии А. 170 (1): 13–22. дои : 10.1007/BF00190397 . ПМИД 1573567 . S2CID 22600955 .
- ↑ Перейти обратно: Перейти обратно: а б с д Уотерман, Джейн (2008). «Глава 3: Стратегии спаривания самцов грызунов». В Вольфе, Джерри О.; Шерман, Пол В. (ред.). Общества грызунов: экологическая и эволюционная перспектива . Издательство Чикагского университета. стр. 28–39. ISBN 978-0-226-90538-9 .
- ↑ Перейти обратно: Перейти обратно: а б Соломан, Нэнси Г.; Кин, Брэйн (2008). «Глава 4: Репродуктивные стратегии самок грызунов». В Вольфе, Джерри О.; Шерман, Пол В. (ред.). Общества грызунов: экологическая и эволюционная перспектива . Издательство Чикагского университета. стр. 42–52. ISBN 978-0-226-90538-9 .
- ^ Накане, Юсуке; Ёсимура, Такаши (15 февраля 2019 г.). «Фотопериодическая регуляция размножения позвоночных». Ежегодный обзор биологических наук о животных . 7 (1). Годовые обзоры : 173–194. doi : 10.1146/annurev-animal-020518-115216 . ISSN 2165-8102 . ПМИД 30332291 . S2CID 52984435 .
- ↑ Перейти обратно: Перейти обратно: а б с Макгуайр, Бетти; Бернис, Уильям Э. (2008). «Глава 20: Родительская забота». В Вольфе, Джерри О.; Шерман, Пол В. (ред.). Общества грызунов: экологическая и эволюционная перспектива . Издательство Чикагского университета. стр. 231–235. ISBN 978-0-226-90538-9 .
- ^ Холмс, Уоррен Г.; Матео, Джилл М. (2008). «Глава 19: Распознавание родства у грызунов: проблемы и доказательства». В Вольфе, Джерри О.; Шерман, Пол В. (ред.). Общества грызунов: экологическая и эволюционная перспектива . Издательство Чикагского университета. стр. 226–227. ISBN 978-0-226-90538-9 .
- ↑ Перейти обратно: Перейти обратно: а б Эбенспергер, Луис А.; Блюмспегер, Дэниел Т. (2008). «Глава 23: Детоубийство неродительского происхождения». В Вольфе, Джерри О.; Шерман, Пол В. (ред.). Общества грызунов: экологическая и эволюционная перспектива . Издательство Чикагского университета. стр. 274–278. ISBN 978-0-226-90538-9 .
- ^ Хугланд, Дж. Л. (1985). «Детоубийство луговых собачек: кормящие самки убивают потомство близких родственников». Наука . 230 (4729): 1037–1040. Бибкод : 1985Sci...230.1037H . дои : 10.1126/science.230.4729.1037 . ПМИД 17814930 . S2CID 23653101 .
- ^ Хакландера, Клаус; Мёстльб, Эрих; Арнольд, Уолтер (2003). «Подавление репродукции у самок альпийских сурков Marmota marmota ». Поведение животных . 65 (6): 1133–1140. дои : 10.1006/anbe.2003.2159 . S2CID 53218701 .
- ↑ Перейти обратно: Перейти обратно: а б Чартерс, Джесси Блаунт Аллен (1904). «Ассоциативные процессы морской свинки: исследование психического развития животного с хорошо медулированной при рождении нервной системой» . Журнал сравнительной неврологии и психологии . XIV (4): 300–337. Архивировано из оригинала 5 марта 2023 года . Проверено 1 ноября 2020 г.
- ^ Джейкобс, Люсия Ф.; Лиман, Эмили Р. (1991). «Серые белки помнят места закопанных орехов» (PDF) . Поведение животных . 41 : 103–110. дои : 10.1016/s0003-3472(05)80506-8 . S2CID 50448069 . Архивировано из оригинала (PDF) 28 июля 2020 года . Проверено 13 августа 2014 г.
- ^ Джейкобс, Люсия Ф. (1992). «Память местоположений тайников у крыс-кенгуру Мерриама» (PDF) . Поведение животных . 43 (4): 585–593. дои : 10.1016/S0003-3472(05)81018-8 . S2CID 14173113 . Архивировано из оригинала (PDF) 26 августа 2014 года.
- ^ Хардинг, Э.Дж.; Пол, ЕС; Мендл, М. (2004). «Поведение животных: когнитивная предвзятость и аффективное состояние» . Природа . 427 (6972): 312. Бибкод : 2004Natur.427..312H . дои : 10.1038/427312а . ПМИД 14737158 . S2CID 4411418 .
- ^ Рыгула, Рафаль; Плута, Елена; Попик, Петр (2012). «Смеющиеся крысы оптимистичны» . ПЛОС ОДИН . 7 (12): е51959. Бибкод : 2012PLoSO...751959R . дои : 10.1371/journal.pone.0051959 . ПМК 3530570 . ПМИД 23300582 .
- ^ Карлайл, Ким (8 марта 2007 г.). «Крысы, способные размышлять над психическими процессами» . Университет Джорджии. Архивировано из оригинала 6 октября 2014 года . Проверено 13 августа 2014 г.
- ^ Фут, Эллисон Л.; Кристалл, Джей Ди (2007). «Метапознание у крысы» . Современная биология . 17 (6): 551–555. Бибкод : 2007CBio...17..551F . дои : 10.1016/j.cub.2007.01.061 . ЧВК 1861845 . ПМИД 17346969 .
- ^ Смит, Дж. Дэвид; Беран, MJ; Коучман, Джей Джей; Коутиньо, MVC (2008). «Сравнительное исследование метапознания: более четкие парадигмы, более безопасные выводы» . Психономический бюллетень и обзор . 15 (4): 679–691. дои : 10.3758/PBR.15.4.679 . ПМК 4607312 . ПМИД 18792496 .
- ^ Йозефовец, Дж.; Стаддон, JER; Черутти, DT (2009). «Метапознание у животных: откуда мы знаем, что они знают?» . Сравнительные обзоры познания и поведения . 4 : 29–39. дои : 10.3819/ccbr.2009.40003 .
- ^ Хэнсон, Энн (2012). «Как крысы выбирают, что есть?» . Поведение и биология крыс . Архивировано из оригинала 7 декабря 2014 года . Проверено 24 августа 2014 г.
- ^ Галеф, Беннетт Г.; Лаланд, Кевин Н. (июнь 2005 г.). «Социальное обучение животных: эмпирические исследования и теоретические модели» . Бионаука . 55 (6): 489–499. doi : 10.1641/0006-3568(2005)055[0489:sliaes]2.0.co;2 .
- ↑ Перейти обратно: Перейти обратно: а б с Кей, Эмили Х.; Хоекстра, Хопи Э. (2008). «Грызуны» . Современная биология . 18 (10): Р406–Р410. Бибкод : 2008CBio...18.R406K . дои : 10.1016/j.cub.2008.03.019 . ПМИД 18492466 .
- ^ Ашер1, Роберт Дж.; Мэн, Джин; Уибл, Джон Р.; Маккенна, Малкольм К.; Ружье, Гильермо В.; Дашевег, Демберлин; Новачек, Майкл Дж. (2005). «Стебель зайцеобразных и древность Глиреса». Наука . 307 (5712): 1091–1094. Бибкод : 2005Sci...307.1091A . дои : 10.1126/science.1107808 . ПМИД 15718468 . S2CID 42090505 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Суарес, Родриго; Фернандес-Абурто, Педро; Мангер, Пол; Мподозис, Хорхе (2011). «Ухудшение вомероназального пути Гао у сексуально диморфных млекопитающих» . ПЛОС ОДИН . 6 (10): Рисунок 4. Бибкод : 2011PLoSO...626436S . дои : 10.1371/journal.pone.0026436 . ПМК 3198400 . ПМИД 22039487 . Архивировано из оригинала 6 апреля 2022 года . Проверено 6 апреля 2022 г.
- ^ Дузери, EJP; Дельсук, Ф.; Стэнхоуп, MJ; Хучон, Д. (2003). «Локальные молекулярные часы в трех ядерных генах: время расхождения для грызунов и других млекопитающих и несовместимость калибровок ископаемых» . Журнал молекулярной эволюции . 57 : С201–13. Бибкод : 2003JMolE..57S.201D . дои : 10.1007/s00239-003-0028-x . ПМИД 15008417 . S2CID 23887665 .
- ^ Хорнер, Д.С.; Лефкиммиатис, К.; Рейес, А.; Гисси, К.; Сакконе, К.; Песоле, Г. (2007). «Филогенетический анализ полных последовательностей митохондриального генома предполагает базальную дивергенцию загадочного грызуна Anomalurus » . Эволюционная биология BMC . 7 (1): 16. Бибкод : 2007BMCEE...7...16H . дои : 10.1186/1471-2148-7-16 . ПМК 1802082 . ПМИД 17288612 .
- ^ Вуд, Д. Джозеф (2010). Вымирание мультитуберкулезных заболеваний за пределами Северной Америки: глобальный подход к тестированию модели конкуренции (MS). Университет штата Огайо. Архивировано из оригинала 8 апреля 2015 года . Проверено 6 апреля 2015 г.
- ^ Шенк, Джон Дж.; Роу, Кевин С.; Степпан, Скотт Дж. (2013). «Экологические возможности и необходимость диверсификации повторных континентальных колонизаций муроидными грызунами». Систематическая биология . 62 (6): 837–864. дои : 10.1093/sysbio/syt050 . ПМИД 23925508 .
- ^ Хопкинс, Саманта С.Б. (2005). «Эволюция окаменелости и адаптивная роль рогов у Mylagaulidae (Mammalia: Rodentia)» . Труды Королевского общества Б. 272 (1573): 1705–1713. дои : 10.1098/rspb.2005.3171 . ПМЦ 1559849 . ПМИД 16087426 .
- ^ Сэмюэлс, Джошуа X.; Занканелла, Джон (2011). «Раннее гемфиллийское появление кастора (Castoridae) из формации Гремучая змея в Орегоне» (PDF) . Журнал палеонтологии . 85 (5): 930–935. дои : 10.1666/11-016.1 . S2CID 128866799 . Архивировано (PDF) из оригинала 24 декабря 2013 года . Проверено 29 июня 2014 г.
- ↑ Перейти обратно: Перейти обратно: а б Мариво, Лоран; Эссид, Эль-Мабрук; Марзуги, Виссем; Аммар, Хайет Хаяти; Аднет, Сильвен; Маранда, Бернар; Мерзеро, Жиль; Табусе, Родольф; Виани-Лио, Моник (2014). «Новый и примитивный вид Protophiomys (Rodentia, Hystricognathi) из позднего среднего эоцена Джебель-эль-Кебара, Центральный Тунис» . Палеовертебрата . 38 (1): 1–17. дои : 10.18563/pv.38.1.e2 . S2CID 55599571 . Архивировано из оригинала 12 августа 2014 года . Проверено 29 июня 2014 г.
- ^ Гербрант, Эммануэль; Ярость, Жан-Клод (2006). «Палеобиогеография Африки: насколько отличается от Гондваны и Лавразии?». Палеогеография, Палеоклиматология, Палеоэкология . 241 (2): 224–246. Бибкод : 2006PPP...241..224G . дои : 10.1016/j.palaeo.2006.03.016 .
- ^ Велес-Хуарбе, Хорхе; Мартин, Томас; Макфи, Росс Д.Э. (2014). «Самые ранние карибские грызуны: олигоценовые кавиоморфы из Пуэрто-Рико». Журнал палеонтологии позвоночных . 34 (1): 157–163. Бибкод : 2014JVPal..34..157V . дои : 10.1080/02724634.2013.789039 . S2CID 140178414 .
- ^ Али, младший; Хубер, М. (20 января 2010 г.). «Биоразнообразие млекопитающих на Мадагаскаре, контролируемое океанскими течениями». Природа . 463 (4 февраля 2010 г.): 653–656. Бибкод : 2010Natur.463..653A . дои : 10.1038/nature08706 . ПМИД 20090678 . S2CID 4333977 .
- ^ Векуа, А.; Бендукидзе О.; Бухсианидзе, М.; Ванишвили Н.; Аугусти, Дж.; Мартинес-Наварро, Б.; Рук, Л. (2010). «Дикобраз в позднем неогене и четвертичном периоде Грузии» (PDF) . Вестник Национальной академии наук Грузии . 4 (3): 140–149. Архивировано из оригинала (PDF) 16 июля 2014 года.
- ^ «Гигантский бобр» . Тетради по естествознанию . Канадский музей природы. 28 мая 2013 г. Архивировано из оригинала 26 января 2021 г. . Проверено 19 октября 2014 г.
- ^ Риндеркнехт, Андрес; Бланко, Р. Эрнесто (2008). «Самый крупный ископаемый грызун» . Труды Королевского общества Б. 275 (1637): 923–928. дои : 10.1098/rspb.2007.1645 . ПМК 2599941 . ПМИД 18198140 .
- ^ Брид, Билл; Форд, Фред (2007). Местные мыши и крысы (PDF) . Издательство CSIRO. стр. 3, 5 и пассим. ISBN 978-0-643-09166-5 . Архивировано (PDF) из оригинала 24 сентября 2015 года . Проверено 29 июня 2014 г.
- ^ «План действий для австралийских грызунов» . Окружающая среда Австралии. 1 апреля 1995 года. Архивировано из оригинала 7 октября 2014 года . Проверено 18 сентября 2014 г.
- ^ Роу, КЦ; Рено, ML; Ричмонд, DM; Адкинс, Р.М.; Степпан, SJ (2008). «Плиоценовая колонизация и адаптивная радиация в Австралии и Новой Гвинее (Сахул): многолокусная систематика старых эндемичных грызунов (Muroidea: Murinae)». Молекулярная филогенетика и эволюция . 47 (1): 84–101. Бибкод : 2008МОЛПЭ..47...84Р . дои : 10.1016/j.ympev.2008.01.001 . ПМИД 18313945 .
- ^ Баскин, Джон А.; Томас, Ронни Г. (2007). «Южный Техас и Великая американская развязка» (PDF) . Сделки Ассоциации геологических обществ побережья Мексиканского залива . 57 : 37–45. Архивировано из оригинала (PDF) 18 июля 2014 года.
- ^ Маршалл, LG; Батлер, РФ; Дрейк, RE; Кертис, Г.Х.; Тедфорд, Р.Х. (1979). «Калибровка Великого американского обмена». Наука . 204 (4390): 272–279. Бибкод : 1979Sci...204..272M . дои : 10.1126/science.204.4390.272 . ПМИД 17800342 . S2CID 8625188 .
- ↑ Перейти обратно: Перейти обратно: а б Смит, Маргарет Ф.; Паттон, Джеймс Л. (1999). «Филогенетические взаимоотношения и радиация сигмодонтных грызунов в Южной Америке: данные цитохрома b». Журнал эволюции млекопитающих . 6 (2): 89–128. дои : 10.1023/А:1020668004578 . S2CID 22355532 .
- ↑ Перейти обратно: Перейти обратно: а б Парада, А.; Пардиньяс, UFJ; Салазар-Браво, Дж.; Д'Элия, Г.; Пальма, RE (март 2013 г.). «Датирование впечатляющего неотропического излучения: молекулярные оценки времени Sigmodontinae (Rodentia) дают представление о его исторической биогеографии». Молекулярная филогенетика и эволюция . 66 (3): 960–968. Бибкод : 2013МОЛПЭ..66..960П . дои : 10.1016/j.ympev.2012.12.001 . hdl : 11336/5595 . ПМИД 23257216 .
- ^ Степпан, Скотт Дж. (18 апреля 2006 г.). «Родентия» . Веб-проект «Древо жизни» . Архивировано из оригинала 17 июля 2014 года . Проверено 14 июля 2014 г.
- ^ «грызун (сущ.)» . Интернет-словарь этимологии . Архивировано из оригинала 18 мая 2015 года . Проверено 7 мая 2015 г.
- ^ Смит, Эндрю Т. «Зайцеморф» . Британская энциклопедия . Архивировано из оригинала 12 августа 2014 года . Проверено 11 августа 2014 г.
- ^ У, Шаоюань; Ву, Вэньюй; Чжан, Фучунь; Да, Цзе; Ни, Сицзюнь; Сун, Чимин; Эдвардс, Скотт В.; Мэн, Джин; Орган, Крис Л. (2012). «Молекулярные и палеонтологические доказательства постмелового происхождения грызунов» . ПЛОС ОДИН . 7 (10): е46445. Бибкод : 2012PLoSO...746445W . дои : 10.1371/journal.pone.0046445 . ПМЦ 3465340 . ПМИД 23071573 .
- ^ Фабр; и др. (2012). «Взгляд на закономерности диверсификации грызунов: филогенетический подход» . Эволюционная биология BMC . 12 (1): 88. Бибкод : 2012BMCEE..12...88F . дои : 10.1186/1471-2148-12-88 . ПМЦ 3532383 . ПМИД 22697210 .
- ↑ Перейти обратно: Перейти обратно: а б Вуд, Альберт Э. (1955). «Пересмотренная классификация грызунов». Журнал маммологии . 36 (2): 165–187. дои : 10.2307/1375874 . JSTOR 1375874 .
- ^ Вуд, Альберт Э. (1958). «Существуют ли подотряды грызунов?». Систематическая биология . 7 (4): 169–173. дои : 10.2307/2411716 . JSTOR 2411716 .
- ^ Карлтон, доктор медицины; Мюссер, Г.Г. (2005). «Орден Грызунов» . В Уилсоне, Дон Э.; Ридер, ДиЭнн М. (ред.). Виды млекопитающих мира – таксономический и географический справочник . Том. 12. ЖУ Пресс. стр. 745–752. ISBN 978-0-8018-8221-0 .
- ^ Ханикатт, Родни Л. (2009). «Грызуны (Rodentia)» (PDF) . В Хеджесе, Южная Каролина; Кумар, С. (ред.). Древо времени жизни . Издательство Оксфордского университета. Архивировано (PDF) из оригинала 23 сентября 2013 года . Проверено 29 сентября 2014 г.
- ^ База данных, Разнообразие млекопитающих (10 августа 2021 г.), База данных разнообразия млекопитающих , doi : 10.5281/zenodo.5175993 , заархивировано из оригинала 22 августа 2021 г. , получено 9 октября 2021 г.
- ↑ Перейти обратно: Перейти обратно: а б Амори, Г.; Гипполити, С. (2003). «Подход к приоритетам сохранения грызунов в 21 веке, основанный на более высоких таксонах» . Биоразнообразие и охрана животных . 26 (2): 1–18. Архивировано из оригинала 14 июля 2014 года . Проверено 27 июня 2014 г.
- ^ Морган, GS (1993). «Четвертичные наземные позвоночные Ямайки». Биостратиграфия Ямайки . Мемуары Геологического общества Америки. Том. 182. стр. 417–442. дои : 10.1130/mem182-p417 . ISBN 978-0-8137-1182-9 .
- ^ «Оценка сохранения грызунов» . ВАЗА. Архивировано из оригинала 15 июля 2014 года . Проверено 27 июня 2014 г.
- ^ Гудинас, Эдуардо (1989). Лидикер, Уильям Зандер (ред.). Грызуны: Мировой обзор видов, требующих сохранения: на основе материалов семинара Группы специалистов по грызунам МСОП/SSC, состоявшегося на Четвертом Международном териологическом конгрессе, 17 августа 1985 г., Эдмонтон, Альберта, Канада . МСОП. п. 23.
- ↑ Перейти обратно: Перейти обратно: а б Пряжка, AP; Фенн, MGP (1992). «Борьба с грызунами в сохранении исчезающих видов» . Материалы 15-й конференции по позвоночным вредителям : документ 12. Архивировано из оригинала 22 июля 2014 г. Проверено 27 июня 2014 г. 3–5 марта 1992 г.
- ^ Хиндвуд, Калифорния (1940). «Птицы острова Лорд-Хау». Эму . 40 (1): 1–86. Бибкод : 1940EmuAO..40....1H . дои : 10.1071/mu940001 .
- ^ «Ланди тупики вернулись с пропасти» . Би-би-си Девон . 22 февраля 2008 г. Архивировано из оригинала 24 сентября 2015 г. Проверено 30 июня 2014 г.
- ^ Митчелл, Хизер (27 мая 2014 г.). «Много тупиков? Новая надежда для Ланди и других британских островов морских птиц» . РСПБ. Архивировано из оригинала 16 июля 2014 года . Проверено 30 июня 2014 г.
- ^ Слезак, Михаил (14 июня 2016 г.). «Выявлено: первые виды млекопитающих исчезли из-за изменения климата, вызванного деятельностью человека» . Хранитель . Лондон. Архивировано из оригинала 14 июня 2016 года . Проверено 8 декабря 2020 г.
- ^ Файнштейн, Келли (1 марта 2006 г.). «Валяние бобровой шапки» . Модный валяный мех . Калифорнийский университет в Санта-Крус. Архивировано из оригинала 6 октября 2014 года . Проверено 24 сентября 2014 г.
- ^ Иннис, Гарольд А. (1999). Торговля мехом в Канаде: введение в канадскую экономическую историю . Университет Торонто Пресс. стр. 9–12. ISBN 978-0-8020-8196-4 .
- ↑ Перейти обратно: Перейти обратно: а б «Излишняя торговля: Одежда и отделка» . Зоопарк Грэнби. Май 2010 г. Архивировано из оригинала 10 августа 2014 г. Проверено 9 августа 2014 г.
- ^ «Искусство и легенда квилтинга Лакота» . Музей и культурный центр Акта Лакота. Архивировано из оригинала 20 марта 2014 года . Проверено 29 июня 2013 г.
- ↑ Перейти обратно: Перейти обратно: а б с д Фидлер, Линвуд А. (1990). «Грызуны как источник пищи» . Материалы четырнадцатой конференции по позвоночным вредителям, 1990 : 149–155. Архивировано из оригинала 11 января 2020 года . Проверено 8 июля 2014 г.
- ^ Ноултон, Дэвид (13 июля 2011 г.). «Морская свинка, домашнее животное или праздничное угощение» . Куско ест . Архивировано из оригинала 14 июля 2014 года . Проверено 5 июля 2014 г.
- ^ Морвели, Уолтер Кораса; Ноултон, Дэвид (5 марта 2012 г.). «Традиционные грязевые печи и духовки делают лучшую еду» . Куско ест . Архивировано из оригинала 15 июля 2014 года . Проверено 6 июля 2014 г.
- ^ Вольф, Джерри О.; Шерман, Пол В. (2008). Общества грызунов: экологическая и эволюционная перспектива . Издательство Чикагского университета. стр. 3–8. ISBN 978-0-226-90538-9 .
- ^ Кринк, Джордж Дж.; Буллок, Джиллиан Р.; Бантон, Трейси (2000). «История, штаммы и модели». Лабораторная крыса (Справочник экспериментальных животных) . Академическая пресса. стр. 3–16. ISBN 978-0-12-426400-7 .
- ^ Морс, Герберт К. (1981). «Лабораторная мышь: историческая оценка» . В Фостере, Генри (ред.). Мышь в биомедицинских исследованиях: история, генетика и дикие мыши . Эльзевир. стр. xi , 1. ISBN 978-0-323-15606-6 .
- ↑ Перейти обратно: Перейти обратно: а б Гад, Шейн К. (2007). Модели животных в токсикологии (2-е изд.). Тейлор и Фрэнсис. стр. 334–402. ISBN 978-0-8247-5407-5 .
- ^ Харкнесс, Джон Э.; Вагнер, Джозеф Э. (1995). Биология и медицина кроликов и грызунов . Уильямс и Уилкинс. стр. 30–39. ISBN 978-0-683-03919-1 .
- ^ Геррини, Анита (2003). Эксперименты с людьми и животными . Джонс Хопкинс. стр. 98–104. ISBN 978-0-8018-7196-2 .
- ^ Грей, Тара (1998). «Краткая история животных в космосе» . Национальное управление по аэронавтике и исследованию космического пространства . Архивировано из оригинала 11 октября 2004 года . Проверено 5 марта 2007 г.
- ^ Шервин, CM (2010). «25: Разведение и благополучие нетрадиционных лабораторных грызунов». В Хубрехте, Р.; Кирквуд, Дж. (ред.). Справочник UFAW по уходу и содержанию лабораторных животных . Уайли-Блэквелл. стр. 359–369.
- ^ Уайнс, Майкл (19 мая 2004 г.). «Гамбийские грызуны рискуют умереть из-за бананов» . Возраст . Архивировано из оригинала 30 сентября 2017 года . Проверено 21 июня 2014 г.
- ^ Мхелела, Хасан (13 сентября 2012 г.). «Гигантские крысы обучены обнаруживать мины и туберкулез в Африке» . Би-би-си. Архивировано из оригинала 28 июня 2014 года . Проверено 27 июня 2014 г.
- ^ Бакалар, Николас (3 января 2011 г.). «Обнаружение туберкулеза: никаких микроскопов, только крысы» . Нью-Йорк Таймс . Архивировано из оригинала 9 мая 2015 года . Проверено 23 августа 2014 г.
- ^ Хардер, Бен (1 мая 2002 г.). «Ученые «гоняют» крыс с помощью пульта» . Нэшнл Географик . Архивировано из оригинала 6 сентября 2013 года . Проверено 9 ноября 2013 г.
- ^ Солон, О. (9 сентября 2013 г.). «Миссия человека — создать системы дистанционного управления собаками, тараканами и акулами» . Проводной . Архивировано из оригинала 4 ноября 2013 года . Проверено 9 декабря 2013 г.
- ^ Сюй, С.; Талвар, СК; Хоули, ES; Ли, Л.; Чапин, Дж. К. (2004). «Многоканальная телеметрическая система для микростимуляции мозга свободно перемещающихся животных». Журнал методов нейробиологии . 133 (1–2): 57–63. doi : 10.1016/j.jneumeth.2003.09.012 . ПМИД 14757345 . S2CID 10823 .
- ^ «Морские свинки» . РСПКА. 2014. Архивировано из оригинала 14 июня 2014 года . Проверено 21 июня 2014 г.
- ^ «Домашние грызуны» . РСПКА. 2014. Архивировано из оригинала 14 июня 2014 года . Проверено 21 июня 2014 г.
- ^ Брокель, Рэй (1983). Песчанки и другие мелкие грызуны . Детская пресса. стр. 5–20. ISBN 978-0-516-01679-5 .
- ^ Меербург, Б.Г.; Синглтон, Греция; Лейрс, Х. (2009). «Год Крысы заканчивается: пора бороться с голодом!». Наука борьбы с вредителями . 65 (4): 351–2. дои : 10.1002/ps.1718 . ПМИД 19206089 .
- ↑ Перейти обратно: Перейти обратно: а б Стенсет, Нильс Хр; Лейрс, Хервиг; Сконхофт, Андерс; Дэвис, Стивен А.; Печ, Роджер П.; Андреассен, Гарри П.; Синглтон, Грант Р.; Лима, Маурисио; Мачанъу, Роберт С.; Макунди, Родос Х.; Чжан, Жибин; Браун, Питер Р.; Ши, Дачжао; Ван, Синьжун (2003). «Мыши, крысы и люди: биоэкономика сельскохозяйственных грызунов-вредителей» . Границы в экологии и окружающей среде . 1 (77): 367–375. дои : 10.2307/3868189 . JSTOR 3868189 .
- ↑ Перейти обратно: Перейти обратно: а б Меербург, Б.Г.; Синглтон, Греция; Кийлстра, А. (2009). «Болезни, передающиеся грызунами, и их риски для здоровья населения». Критические обзоры по микробиологии . 35 (3): 221–70. дои : 10.1080/10408410902989837 . ПМИД 19548807 . S2CID 205694138 .
- ^ Маккормик, М. (2003). «Крысы, коммуникации и чума: к экологической истории» (PDF) . Журнал междисциплинарной истории . 34 (1): 1–25. дои : 10.1162/002219503322645439 . S2CID 128567627 . Архивировано из оригинала (PDF) 22 февраля 2015 года.
- ^ «Болезни, передающиеся грызунами» . Европейский центр профилактики и контроля заболеваний. Архивировано из оригинала 3 сентября 2014 года . Проверено 1 сентября 2014 г.
- ^ «Болезни, передающиеся грызунами косвенно» . Центры по контролю и профилактике заболеваний. 2012. Архивировано из оригинала 18 августа 2014 года . Проверено 1 сентября 2014 г.
- ^ Центры по контролю и профилактике заболеваний (2006 г.). Комплексная борьба с вредителями: проведение городских обследований на грызунах (PDF) . Атланта: Министерство здравоохранения и социальных служб США. Архивировано (PDF) из оригинала 20 июля 2017 года . Проверено 11 сентября 2017 г.
- ^ Водзицкий, К. (1973). «Перспективы биологической борьбы с популяциями грызунов» . Бюллетень Всемирной организации здравоохранения . 48 (4): 461–467. ПМК 2481104 . ПМИД 4587482 .
- ^ «Борьба с грызунами в сельском хозяйстве – руководство HGCA» . Совет по развитию сельского хозяйства и садоводства. 2012. Архивировано из оригинала 25 апреля 2019 года . Проверено 24 февраля 2018 г.
Дальнейшее чтение [ править ]
- Маккенна, Малкольм К.; Белл, Сьюзен К. (1997). Классификация млекопитающих выше видового уровня . Издательство Колумбийского университета. ISBN 978-0-231-11013-6 .
- Уилсон, Делавэр; Ридер, Д.М., ред. (2005). Виды млекопитающих мира: таксономический и географический справочник . Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-8221-0 .
- Карлтон, доктор медицины; Массер, Г.Г. «Орден Rodentia», страницы 745–752 в книге Wilson & Reeder (2005).
Внешние ссылки [ править ]
Зоология, остеология, сравнительная анатомия [ править ]
- ArchéoZooThèque: Остеология грызунов. Архивировано 29 января 2015 г. в Wayback Machine (фотографии).
- ArchéoZooLibrary: рисунки скелетов грызунов
Разное [ править ]
- Африканские грызуны
- Фотографии грызунов на Flickr
- Информационные бюллетени о видах грызунов Национальной ассоциации по борьбе с вредителями, касающиеся оленей, норвежских крыс и других видов грызунов.