Оризомис
| Оризомис Временной диапазон: Ранчолабриан (300 000 лет назад) – настоящее время.
| |
|---|---|

| |
| Болотная рисовая крыса ( Oryzomys palustris ) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Грызуны |
| Семья: | Крисетиды |
| Подсемейство: | сигмодонтинае |
| Племя: | Оризамиин |
| Род: | Оризомис Бэрд , 1857 г. |
| Типовой вид | |
| Mus palustris Харлан , 1837 г.
| |
| Разновидность [ 1 ] | |

| |
| Распространение Oryzomys : темно-синяя болотная рисовая крыса ( O. palustris ); светло-голубой, бывший ареал болотной рисовой крысы; красный, О. couesi ; розовый, O. albiventer ; темно-зеленый, O. peninsulae ; оранжевый, О. nelsoni ; светло-зеленый, O. antillarum ; желтый, O. dimidiatus и O. couesi ; коричневый, О. горгаси . | |
| Синонимы [ 1 ] | |
| |
Oryzomys — род полуводных обитающих грызунов трибы Oryzomyini, на юге Северной Америки и на крайнем севере Южной Америки. Включает восемь видов , два из которых — болотная рисовая крыса ( О. palustris ) США и О. couesi Мексики и Центральной Америки — широко распространены; шесть других имеют более ограниченное распространение. У этого вида богатая таксономическая история, и большинство видов когда-то относились к болотной рисовой крысе; в будущем могут быть признаны дополнительные виды. Название Oryzomys было введено в 1857 году Спенсером Фуллертоном Бэрдом для болотной рисовой крысы и вскоре было применено к более чем ста видам американских грызунов. Впоследствии род постепенно стал более узким, пока его нынешнее содержание не было установлено в 2006 году, когда было установлено десять новых родов для видов, ранее отнесенных к Oryzomys .
Виды Oryzomys — это крысы среднего размера с длинным грубым мехом. Верхняя часть тела от серого до красноватого цвета, нижняя часть тела от белого до желтовато-желтого цвета . У животных широкие ступни с редуцированными или отсутствующими когтевыми пучками волос вокруг когтей и, по крайней мере, у некоторых видов, с перепонками между пальцами ног . Рострум (передняя часть черепа) широкий, черепная коробка высокая. И болотная рисовая крыса, и O. couesi имеют 56 хромосом , лишены желчного пузыря и имеют сложный пенис (что характерно для Sigmodontinae) с некоторыми чертами, редкими среди oryzomyines; эти характеристики неизвестны у других видов этого рода.
Среда обитания включает различные виды водно-болотных угодий, таких как озера, болота и реки. Виды Oryzomys хорошо плавают, активны в ночное время и питаются как растительной, так и животной пищей. Они строят плетеные гнезда из растительности. После периода беременности от 21 до 28 дней рождается около четырех детенышей. Виды Oryzomys заражены многочисленными паразитами и переносят как минимум три хантавируса , один из которых ( вирус Байу ) также заражает человека. Два, может быть, три вида вымерли за последние два столетия, и по крайней мере еще один находится под угрозой исчезновения, но широко распространенная болотная рисовая крыса и O. couesi не находятся под угрозой.
Таксономия
[ редактировать ]Oryzomys — один из примерно тридцати родов трибы Oryzomyini , разнообразной группы, насчитывающей более ста видов, многие из которых ранее также входили в состав Oryzomys . [ 9 ] Oryzomyini — одно из нескольких триб подсемейства Sigmodontinae семейства Cricetidae , в которое входят сотни других видов преимущественно мелких грызунов, распространенных преимущественно в Америке и Евразии. [ 10 ]
В рамках Oryzomyini филогенетический анализ, проведенный Марсело Векслером в 2006 году, в котором использовались как морфологические данные, так и данные о последовательностях ДНК, обнаружил некоторые доказательства того, что Oryzomys наиболее тесно связан с группой, включающей Holochilus , Lundomys и Pseudoryzomys . Хотя анализ, основанный на морфологических и объединенных данных, подтвердил эту взаимосвязь, последовательности одного только гена Rbp3 вместо этого поместили Oryzomys в группу, в которую входили Nectomys , Sigmodontomys и несколько других родов. Во всех анализах Oryzomys фигурировал в кладе D Oryzomyini. [ 11 ] Отношения между Oryzomys и группой Holochilus были подтверждены пятью синапоморфиями (общими производными признаками) - отсутствием или редукцией как гипотенара, так и межпальцевых подушечек ; уменьшение ногтевых пучков волос вокруг когтей; наличие заднего края скуловой пластинки черепа на том же уровне, что и передний край первого верхнего моляра ; и передний конус (передний бугор) первого верхнего моляра, разделенный переднесрединной ямкой . Первые три представляют собой адаптации к полуводному образу жизни, который разделяют Oryzomys и члены группы Holochilus , и, таким образом, могут быть примерами конвергентной эволюции . [ 12 ]
Ограничение
[ редактировать ]Название Oryzomys было введено в 1857 году Спенсером Фуллертоном Бэрдом для болотной рисовой крысы (ныне Oryzomys palustris ) на востоке Соединенных Штатов. [ 13 ] который был впервые описан двадцатью годами ранее Ричардом Харланом . [ 8 ] Название сочетает в себе греческое слово oryza «рис» и mys «мышь» и относится к привычкам питания болотной рисовой крысы. [ 14 ] [ 15 ] Бэрд поместил Oryzomys в подрод ныне несуществующего рода Hesperomys и включил в него только болотную рисовую крысу - классификацию, которой следовал Эллиот Куэс в 1874 и 1877 годах. [ 16 ] В 1890 г. Oryzomys был возведен в ранг рода, а в последующие годы к нему были приписаны многочисленные дополнительные виды, многие из которых вскоре были выделены в отдельные роды. [ 17 ] В Каталоге млекопитающих 1898 года Луи Эдуард Труессар перечислил 67 видов Oryzomys . [ 18 ] включая некоторые, которые теперь отнесены к Calomys , Necromys , Thomasomys и другим родам, не связанным с Oryzomys . [ 19 ] Некоторые из предложенных новых родов вскоре снова были отнесены к Oryzomys . [ 20 ] а в книге «Семейства и роды живых грызунов» (1941) Джон Эллерман перечислил Microryzomys , Oligoryzomys , Melanomys , Nesoryzomys и Oecomys как синонимы Oryzomys . [ 21 ] и включил в него около 127 видов. [ 22 ] В 1948 году Филип Гершковитц предположил, что другие оризомиины, такие как Nectomys и Megalomys, также могут быть включены в Oryzomys . [ 23 ] и Клейтон Рэй последовал этому предложению в 1962 году. [ 24 ]
Классификация Гершковица и Рэя никогда не получила широкого распространения, и с 1976 года авторы начали восстанавливать некоторые другие группы, отнесенные к Oryzomys, как отдельные роды. [ 25 ] род был сокращен до 43 видов (из 110 в Oryzomyini) В третьем издании (2005 г.) журнала Mammal Species of the World . [ 26 ] но это еще не была естественная монофилетическая группа; [ 27 ] скорее, он в основном объединял те оризомиины, у которых не было ярко выраженной специализации других родов. [ 28 ] В 2006 году всесторонний филогенетический анализ Марсело Векслера предоставил дополнительные доказательства того, что этот род был полифилетическим , поскольку виды Oryzomys были рассредоточены по всему оризомииновому дереву. Он предложил создать одиннадцать новых родов для включения тех видов, которые не были тесно связаны с видом типовым Oryzomys , болотной рисовой крысы; [ 29 ] он рассматривал другие варианты, которые потребовали бы меньшего количества новых родов, но утверждал, что это приведет к появлению менее значимых групп на уровне рода в Oryzomyini. [ 30 ] Позже в том же году Векслер, Персекильо и Восс создали десять новых родов — Aegialomys , Cerradomys , Eremoryzomys , Euryoryzomys , Hylaeamys , Mindomys , Nephelomys , Oreoryzomys , Sooretamys и Transandinomys — для видов, ранее отнесенных к Oryzomys , и поместили еще шесть родственных видов. к "Oryzomys" alfaroi в Handleyomys в ожидании описания для них новых родов. [ 31 ] Они оставили только пять видов у Oryzomys , который теперь наконец стал естественной монофилетической группой. Благодаря последующей таксономической работе количество видов с тех пор увеличилось как минимум до восьми. [ 32 ]
Некоторые проблемы остаются: ? Oryzomys pliocaenicus , миоценовая окаменелость из Канзаса, идентичность неясна, но может принадлежать Bensonomys , [ 33 ] и окаменелости из миоцена Орегона и плиоцена Нью-Мексико также были отнесены к Oryzomys , но, вероятно, ошибочно. [ 33 ] Возможный вид Oryzomys был зарегистрирован из ирвингтонского периода (плейстоцена) Саскачевана . [ 34 ]
Разновидность
[ редактировать ]
Современная концепция Oryzomys происходит от группы palustris-mexicanus в составе гораздо более крупного рода Oryzomys , признанной Мерриамом (1901) , и группы palustris , предложенной Гольдманом (1918). [ 1 ] Мерриам выделил в своей группе 21 вид, но Голдман объединил их в восемь — болотную рисовую крысу в США, O. couesi на крайнем юге Техаса, в Мексике и Центральной Америке, а также шесть других видов с небольшим распространением. [ 37 ] В 1960 году Рэймонд Холл объединил O. couesi и болотную рисовую крысу в один вид, Oryzomys palustris , после чего в состав O. palustris были включены и другие локализованные формы . [ 38 ] Гершковитц описал еще один вид этой группы, O. gorgasi из Колумбии, в 1970 году. [ 39 ] а в следующем году он отметил, что O. dimidiatus , ранее отнесенный к нектомиям , был похож на O. palustris . [ 40 ] После 1979 года болотная рисовая крыса и O. couesi снова стали считаться отдельными в результате дальнейших исследований в Техасе, где их ареалы пересекаются. [ 38 ] При обзоре O. gorgasi в 2001 году Дж. Санчес Х. и его коллеги переопределили и охарактеризовали группу O. palustris и перечислили O. couesi , O. dimidiatus , O. gorgasi и болотную рисовую крысу в качестве ее членов; [ 41 ] Гай Массер и Майкл Карлтон в третьем издании журнала «Виды млекопитающих мира» за 2005 г. дополнительно перечислили O. nelsoni с острова Мария-Мадре на западе Мексики. [ 8 ]
В 2006 году Векслер и его коллеги последовали определению Санчеса и других, данному в 2001 году для ограниченного рода Oryzomys , но добавили O. antillarum с Ямайки . в качестве вида [ 42 ] Карлтон и Хоакин Арройо-Кабралес исследовали Oryzomys из западной Мексики в 2009 году и в этом контексте предоставили расширенный диагноз Oryzomys . Они признали восемь видов: шесть ранее упомянутых, а также O. albiventer и O. peninsulae . [ 1 ] Также в 2009 году Роберт Восс и Векслер идентифицировали субископаемое Oryzomys curasoae с Кюрасао как островную популяцию O. gogasi . [ 43 ] В следующем году Делтон Хэнсон и его коллеги опубликовали исследование, в котором использовались данные о последовательностях ДНК цитохрома b , межфоторецепторного ретиноидсвязывающего белка и алкогольдегидрогеназы 1 для оценки взаимоотношений внутри Oryzomys . Они рекомендовали разделить болотную рисовую крысу на два вида, а O. couesi — на четыре вида на основе наблюдаемого расхождения последовательностей и других данных. [ 44 ]
Мерриам разделил свою группу palustris-mexicanus на две «серии» в зависимости от цвета нижней части тела (белая или буроватая ). [ 45 ] Гольдман разделил свою группу palustris на две «секции»: couesi секцию с O. couesi и шестью родственными видами и секцию palustris только с O. palustris . Он отметил, что последние отличались от первых в целом более темным, буроватым, более длинным мехом и более крупными клиновидно-небными впадинами (отверстиями в мезоптеригоидной ямке , щелью позади конца неба ) . [ 46 ] Поскольку анализ Векслера 2006 года включал только O. couesi и болотную рисовую крысу среди видов Oryzomys в строгом смысле этого слова, он не смог проверить эти группы. [ 47 ] Карлтон и Арройо-Кабралес согласились с разделением Гольдмана, перечислив дополнительных признаков, и отметили, что группа palustris может быть более адаптирована к полуводному образу жизни, чем члены группы couesi . У последних мех обычно красновато-коричневый, в отличие от серовато-коричневого у группы palustris . Члены группы couesi имеют меньшие клиновидно-небные впадины и меньшее клиновидно-небное отверстие , отверстие (отверстие) сбоку черепа над коренными зубами и более развитую переднегубную поясную часть на третьем нижнем моляре (гребень в передней части зуб). Подушечка гипотенара задней части стопы, расположенная на подошве вдали от пальцев, имеется в группе couesi , но отсутствует в группе palustris . Межпальцевая перепонка может быть более развита у группы palustris . [ 36 ] Используя морфологические данные, Восс и Векслер обнаружили более тесную связь между O. couesi и O. gogasi , исключив O. palustris , но с низкой достоверностью. [ 48 ] Данные о последовательности ДНК, полученные Хэнсоном и его коллегами, подтвердили глубокое разделение между группами palustris и couesi , но образец из Коста-Рики (отнесенный к O. couesi ) был примерно так же далек от этих двух групп, как и друг от друга. [ 49 ]
В настоящее время род включает следующие виды: [ 1 ]
| Имя | Распределение | Характеристики | Таксономические комментарии |
|---|---|---|---|
| Оризомис альбивентер | Мичоакан , Гуанахуато , Халиско (внутренняя часть западной Мексики) [ 50 ] | Большой, длинный хвост, крепкий череп. [ 50 ] | Описан в 1901 г. как вид; подвид O. couesi в 1918 г.; восстановлен как вид в 2009 г. [ 51 ] |
| Оризомис антилларум | Ямайка (вымершие) [ 52 ] | Длинные носовые кости , короткие резцовые отверстия (перфорации в небе ), крепкие скуловые дуги (скулы). [ 53 ] | Описан в 1898 г. как вид; подвид O. couesi в 1966 г.; восстановлен как вид в 1993/2006 г. [ 54 ] |
| Оризомис куэзи | Южный Техас и прибрежная Сонора (северо-запад Мексики) до северо-запада Колумбии. [ 55 ] | Верхняя часть тела от желтовато-желтой до красноватой, нижняя часть тела от белого до желтовато-желтого цвета, клиновидно-небные впадины небольшие или отсутствуют. [ 56 ] | Описан как вид в 1877 г.; подвид болотной рисовой крысы в 1960 г.; восстановлен как вид в 1979 г.; различные другие виды время от времени отделялись от него и смешивались с ним. [ 57 ] Генетические данные предполагают, что могут присутствовать четыре вида: один вдоль побережья Тихого океана от Соноры до Сальвадора; один из Техаса в Никарагуа; один в Коста-Рике; и один в Панаме. [ 44 ] |
| Оризомис половина | Юго-восточная Никарагуа [ 58 ] | Нижняя часть тела серая, лапы коричневые, хвост сверху почти такого же цвета, как снизу. [ 59 ] | Описан в 1905 году как разновидность нектомий ; реклассифицирован в Оризомис в 1948 г. [ 60 ] |
| Оризомис горгаси | Северо-западная Колумбия , северо-западная Венесуэла ; вымершие на Кюрасао [ 61 ] | Крепкий рострум (передняя часть черепа); резцовое отверстие, суживающееся сзади; клиновидно-небные пустоты отсутствуют; подсквамозное окно (отверстие в задней части черепа) маленькое [ 62 ] | Описан в 1970 г.; Oryzomys curasoae описан в 2001 г.; [ 58 ] оба синонимичны в 2009 году [ 43 ] |
| Оризомис Нельсони | Остров Мария Мадре , Наярит, западная Мексика (вымершие) [ 63 ] | Большой, длинный хвост, головотрубо тяжелое и загнутое вниз, резцы большие и широкие. [ 64 ] | Описан в 1898 г. как вид; подвид O. palustris в 1971 г.; иначе рассматривается как отдельный вид [ 63 ] |
| Болотная рисовая крыса ( Oryzomys palustris ) | На востоке Соединенных Штатов от Нью-Джерси и Канзаса на юг до Флориды и Техаса , а также до Тамаулипаса , Мексика; ранее дальше на север, в Айову и юго-запад Пенсильвании. [ 65 ] | По сравнению с симпатрическим O. couesi , где они встречаются: короткий хвост, белая нижняя часть тела, большое клиновидно-небное отверстие. [ 66 ] | Описан в 1827 г.; [ 63 ] особый статус формы Флорида-Кис ( argentatus ; впервые описан в 1978 году) оспаривается; [ 67 ] генетические данные позволяют предположить, что популяции к западу от Алабамы могут представлять собой отдельный вид. [ 68 ] |
| Полуостров Оризомис | Южная оконечность Южной Нижней Калифорнии (возможно, вымершая) [ 69 ] | Умеренно большой, серый на голове и передних конечностях, широкие квадратные скуловые дуги, длинные и широкие резцовые отверстия, верхний резец почти ортодонтный. [ 70 ] | Описан в 1899 г. как вид; подвиды O. palustris в 1971 г. и O. couesi в 1994 г.; восстановлен как вид в 2009 г. [ 70 ] |
Описание
[ редактировать ]| Разновидность | н [ Примечание 4 ] | Общая длина | Хвост | Задняя лапа |
|---|---|---|---|---|
| Оризомис альбивентер [ 71 ] | 12 | 285.4 (245–314) | 155.4 (129–173) | 36.1 (33–40) |
| Оризомис антилларум [ 72 ] | 3 | 247 (228–260) | 119.7 (108–132) | 29.3 (28–30) |
| Oryzomys couesi из Наярита [ 71 ] | 62 | 244.8 (210–288) | 125.1 (105–150) | 30.5 (27–33) |
| Оризомис половина [ 73 ] | 3 | 249 (228–278) | 129 (110–150) | 28.3 (27–31) |
| Оризомис горгаси [ 74 ] | 6–10 [ Примечание 5 ] | 259 (220–290) | 130 (116–138) | 31 (30–32) |
| Оризомис Нельсони [ 71 ] | 4 | 322 (288–344) | 181.5 (160–191) | 37.3 (35–39) |
| Болотная рисовая крыса [ 75 ] | – | 226–305 | 108–156 | 28–37 |
| Полуостров Оризомис [ 71 ] | 14 | 265.6 (227–305) | 136.8 (114–156) | 32.0 (29–34) |
| Измерения производятся в миллиметрах и в виде «среднее (минимум–максимум)». | ||||
В состав Oryzomys входят полуводные специализированные грызуны-оризомиины среднего размера. У них длинный грубый мех от серовато-красноватого цвета на верхней части тела и от белого до желтоватого цвета на нижней части тела. [ 76 ] Болотная рисовая крыса внешне напоминает интродуцированные виды черную крысу и коричневую крысу , но имеет большие различия в цвете верхней и нижней частей тела. [ 75 ] Вибриссы (бакенбарды) короткие , уши маленькие и хорошо опушенные. Хвост обычно такой же длины или длиннее, чем голова и тело, и покрыт редкими волосами, но волосы на нижней стороне длиннее, чем наверху. У самок восемь молочных желез , как и у большинства оризомиин. Задние лапы широкие, первый и пятый пальцы заметно короче средних трех. Верхняя поверхность волосистая, а нижняя голая и покрыта мелкими неровностями ( чешуйками ). Подушечки, как и ногтевые пучки, обычно развиты слабо. [ 77 ] Межпальцевая перепонка может присутствовать, но ее развитие в пределах рода варьируется. [ 36 ]
Кариотип . зарегистрирован в различных популяциях болотной рисовой крысы и O. couesi и, по-видимому, стабилен внутри рода при 56 хромосомах , при этом фундаментальное число хромосомных плеч колеблется от 56 до 60 (2n = 56, FN = 56–60) ). [ 44 ] У обоих видов желудок имеет характерный рисунок сигмодонтов ( однокамерно -гемигландулярный): он не разделен на две камеры incisura angularis и передняя часть (антральный отдел) покрыта железистым эпителием . [ 78 ] Кроме того, желчный пузырь отсутствует - синапоморфия Oryzomyini. [ 79 ]
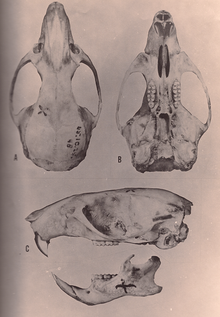
Виды Oryzomys имеют большой череп с коротким рострумом и высокой черепной коробкой . , Межглазничная область расположенная между глазами, наиболее узкая спереди и по краям окружена хорошо развитыми бусинами. Скуловая пластинка широкая, спереди имеет хорошо развитую скуловую вырезку. Скуловая дуга крепкая и содержит небольшую, но отчетливую скуловую кость . , Межтеменная кость часть крыши черепной коробки, узкая и короткая; [ 81 ] его узость является синапоморфией O. couesi плюс болотной рисовой крысы. согласно анализу Векслера, [ 12 ] Режущие отверстия длинные, их задний край находится впереди первых коренных зубов или дальше назад. Небо также длинное, выходит за задний край верхнечелюстной кости и перфорировано около третьих моляров хорошо развитыми заднелатеральными небными ямками . нет Алисфеноидной стойки , продолжения алисфеноидной кости , которая у некоторых других оризомиин разделяет два отверстия в черепе, . Слуховые буллы крупные. Состояние артерий головы крайне тяжелое . [ 81 ] В нижней челюсти (нижней челюсти) хорошо развит венечный отросток — отросток сзади. [ 82 ] заметен капсульный отросток , приподнятая нижнечелюстная кость, в которой находится корень нижнего резца. [ 46 ]
Как обычно у оризомиин, коренные зубы пенталофодонтные (имеют мезолофы и мезолофиды , хорошо развитые дополнительные гребни) и бунодонтные , с бугорками выше соединительных гребней. [ 1 ] Бугры на верхних коренных зубах расположены двумя продольными рядами, а не тремя, как у черных и коричневых крыс. [ 83 ] Передние бугры первого верхнего и нижнего моляра (переднеконус и переднеконид соответственно) широкие и не разделены полностью переднемедиальным изгибом или флексидом . Позади переднего конуса антеролоф (меньший гребень) завершен и отделен от переднего конуса. [ 1 ] И на втором, и на третьем нижних коренных зубах присутствует антеролофид (гребень на внутреннем переднем углу), предполагаемая синапоморфия этого рода. [ 12 ] Первые коренные зубы помимо основных имеют дополнительные маленькие корни, так что верхний первый моляр имеет четыре, а нижний — три или четыре корня. [ 1 ]
Как характерно для Sigmodontinae, болотная рисовая крыса и O. couesi имеют сложный половой член с бакулюмом (костью полового члена) с большими выступами по бокам. [ 84 ] Внешняя поверхность полового члена в основном покрыта небольшими шипами , но имеется широкая полоса неостистой ткани. [ 85 ] Сосочек (сосковый выступ) на дорсальной (верхней) стороне полового члена покрыт маленькими шипами - признак, который эти два вида разделяют только с Oligoryzomys среди исследованных oryzomyines. [ 86 ] На уретральном отростке , расположенном в кратере на конце полового члена, [ 87 ] мясистый отросток ( подверхушечная долька имеется ); он отсутствует у всех других оризомиинов с изученными пенами, кроме Holochilus brasiliensis . [ 88 ] Оба признака обнаружены как синапоморфии O. couesi и болотной рисовой крысы в анализе Векслера. [ 12 ]
Распространение, экология и поведение
[ редактировать ]
Ареал Oryzomys простирается от Нью-Джерси на востоке США через Мексику и Центральную Америку на юг до северо-запада Колумбии и с востока до северо-запада Венесуэлы и Кюрасао. [ 90 ] Виды Oryzomys обычно обитают во влажных местах обитания, таких как болота, ручьи и мангровые заросли . [ 1 ] но и болотная рисовая крыса, и O. couesi иногда встречаются и в более засушливых местах обитания. [ 91 ] Они встречаются или встречались на многих континентального шельфа островах и на одном океаническом острове , Ямайке; их способность колонизировать острова может быть вызвана их тесной связью с водой и частым появлением на прибрежных водно-болотных угодьях. [ 69 ] Самые старые окаменелости датируются ранчолабрианами в Соединенных Штатах, около 300 000 лет назад; хотя были и более ранние записи в Северной Америке, на самом деле они не относятся к Oryzomys или даже Oryzomyini. [ 92 ] Oryzomyines, вероятно, возникли в Южной Америке к востоку от Анд; Считается, что присутствие Oryzomys в Центральной Америке и других трансандских регионах является результатом одного из нескольких независимых вторжений в этот регион оризомиин. [ 93 ] Альтернативно, Oryzomys мог произойти от плиоценового североамериканского Jacobsomys . [ 94 ] O. antillarum , возможно, достиг Ямайки во время последнего ледникового периода , когда уровень моря был низким. [ 52 ]
Поведение известно в основном по болотной рисовой крысе и O. couesi , с некоторыми разрозненными данными по другим видам. Оризоми ведут полуводный образ жизни, проводят много времени в воде, а в остальном в основном живут на земле; [ 95 ] И болотная рисовая крыса, и O. couesi известны как отличные пловцы и убегают в воду, если их потревожить. [ 96 ] Оба также активны ночью и строят гнезда из переплетенной растительности, которые можно подвешивать над водой. [ 97 ] У обоих видов размножение может происходить в течение всего года, но известно, что у болотной рисовой крысы оно сезонно изменчиво. [ 98 ] В обоих случаях беременность длится от 21 до 28 дней, а размер помета обычно составляет от одного до семи, в среднем от трех до пяти. [ 99 ] Молодые болотные рисовые крысы и O. couesi становятся репродуктивно активными примерно в 50-дневном возрасте. [ 100 ]
Болотная рисовая крыса O. couesi и O. gogasi , как известно, всеядны и питаются как растительным, так и животным материалом. Они едят как семена, так и зеленые части растений, а также поедают различных животных, включая насекомых , ракообразных и многих других. [ 101 ] Сипуха . ( Tyto alba ) — главный хищник болотной рисовой крысы [ 102 ] а остатки O. antillarum , O. couesi и O. gorgasi были обнаружены в отложениях пометов сов . [ 103 ] Известно, что несколько других животных охотятся на Oryzomys . [ 104 ] множество паразитов. известно У O. couesi [ 105 ] и болотная рисовая крыса [ 106 ] и две паразитические нематоды были обнаружены у O. gorgasi . [ 107 ]
Человеческое взаимодействие
[ редактировать ]Два вида Oryzomys , O. antillarum и O. nelsoni , вымерли с 19 века. [ 108 ] и третий, O. peninsulae , вряд ли до сих пор сохранился. [ 69 ] Их исчезновение могло быть вызвано разрушением среды обитания и интродуцированными видами, такими как небольшой азиатский мангуст , коричневая и черная крыса. [ 109 ] Эти же причины могут угрожать O. gorgasi , который Красный список МСОП оценивает как « находящийся под угрозой исчезновения ». [ 110 ] O. albiventer пострадал от изменения человеком среды обитания, но, вероятно, все еще выживает. [ 111 ] Напротив, широко распространенные виды, болотная рисовая крыса и O. couesi , являются обычными и не представляют опасности для сохранения (действительно, оба считаются вредителями), но некоторые популяции находятся под угрозой. [ 112 ] Как и эти два вида, O. dimidiatus внесен как « вызывающий наименьшие опасения ». в Красный список [ 113 ]
Болотная рисовая крыса является естественным резервуаром вируса Байу , второй наиболее распространенной причины хантавирусного легочного синдрома в Соединенных Штатах. [ 114 ] Два других хантавируса, вирус Катакамас и вирус Плайя-де-Оро , встречаются у O. couesi в Гондурасе и западной Мексике соответственно, но не известно, что они заражают людей. [ 115 ]
Примечания
[ редактировать ]- ^ Как подрод Hesperomys .
- ^ Первое использование как рода. [ 1 ] Голдман [ 4 ] и Тейт [ 5 ] приписывают первое использование Oryzomys как рода Coues также в 1890 году. [ 6 ]
- ^ Голое имя («Обнаженное имя»). [ 8 ]
- ^ Количество измеренных образцов.
- ^ Шесть для общей длины, восемь для длины хвоста и десять для длины задней части стопы.
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж Карлтон и Арройо-Кабралес, 2009, с. 116
- ^ Бэрд, 1857, с. 482, ср. п. 459
- ^ Аллен, 1890, с. 187
- ^ Гольдман, 1918, с. 11
- ^ Тейт, 1932, с. 4
- ^ Куэс, 1890, с. 4164
- ^ Гершовиц, 1948, с. 54
- ^ Jump up to: а б с Массер и Карлтон, 2005, с. 1144
- ^ Векслер, 2006, стр. 1, 10; Векслер и др., 2006, стр. 1, таблица 1.
- ^ Массер и Карлтон, 2005 г.
- ^ Векслер, 2006, рис. 34–39
- ^ Jump up to: а б с д Векслер, 2006, стр. 131.
- ^ Бэрд, 1857, с. 482
- ^ Шварц и Шварц, 2001, с. 192
- ^ ὄρυζα , μῦς . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей» .
- ^ Куэ, 1874, стр. 183–184; 1877, с. 113
- ^ Тейт, 1932, стр. 4 и далее.
- ^ Труссар, 1898, стр. 523–527
- ^ Тейт, 1932, стр. 6–7; Массер и Карлтон, 2005, стр. 1105, 1108, 1128, 1130, 1144, 1179, 1180.
- ^ Векслер, 2006, таблица 1; Массер и Карлтон, 2005, с. 1144
- ^ Эллерман, 1941, с. 340
- ^ Эллерман, 1940, стр. 345–359
- ^ Гершковиц, 1948, с. 54, сноска 1
- ^ Рэй, 1962, стр. 16–26.
- ^ Векслер, 2006, с. 10, таблица 1; Массер и Карлтон, 2005, с. 1144
- ^ Массер и Карлтон, 2005, с. 900
- ^ Массер и Карлтон, 2005, с. 1144; Векслер, 2006, с. 10
- ^ Векслер, 2006, стр. 82.
- ^ Векслер, 2006, стр. 75.
- ^ Векслер, 2006, рис. 42, стр. 77.
- ^ Векслер и др., 2006, стр. 1–2
- ^ Карлтон и Арройо-Кабралес, 2009, стр. 115–116; Векслер и др., 2006, таблица 1.
- ^ Jump up to: а б Векслер, 2006, стр. 87.
- ^ Черчер, 1984, с. 149
- ^ Элиот, 1904, с. 240
- ^ Jump up to: а б с Карлтон и Арройо-Кабралес, 2009 г.
- ^ Гольдман, 1918, с. 16
- ^ Jump up to: а б Массер и Карлтон, 2005, с. 1147
- ^ Гершковитц, 1970, с. 700
- ^ Гершковитц, 1971, стр. 789, 791.
- ^ Санчес и др., 2001, стр. 209–210
- ^ Векслер, 2006, таблица 1, сноска e.
- ^ Jump up to: а б Восс и Векслер, 2009, стр. 73.
- ^ Jump up to: а б с Хэнсон и др., 2010, стр. 342–343.
- ^ Мерриам, 1901, с. 275
- ^ Jump up to: а б Гольдман, 1918, с. 20
- ^ Векслер, 2006, таблица 4.
- ^ Восс и Векслер, 2009, рис. 1
- ^ Hanson et al., 2010, рис. 2–5, таблица 1
- ^ Jump up to: а б Карлтон и Арройо-Кабралес, 2009, стр. 118
- ^ Карлтон и Арройо-Кабралес, 2009, стр. 117–118
- ^ Jump up to: а б Морган, 1993, с. 439
- ^ Гольдман, 1918, с. 44
- ^ Морган, 1993, стр. 439; Weksler et al., 2006, таблица 1, сноска e.
- ^ Массер и Карлтон, 2005, с. 1147; Карлтон и Арройо-Кабралес, 2009, с. 119
- ^ Гольдман, 1918, с. 29
- ^ Массер и Карлтон, 2005, с. 1147; Карлтон и Арройо-Кабралес, 2009 г., стр. 101–1. 94–95
- ^ Jump up to: а б Массер и Карлтон, 2005, с. 1148
- ^ Рид, 2009, с. 207
- ^ Гершковитц, 1948, стр. 54–55.
- ^ Массер и Карлтон, 2005, с. 1149; Восс и Векслер, 2009, с. 73
- ^ Санчес и др., 2001, с. 210
- ^ Jump up to: а б с Массер и Карлтон, 2005, с. 1152
- ^ Карлтон и Арройо-Кабралес, 2009, стр. 121–122
- ^ Массер и Карлтон, 2005, с. 1152; Шмидт и Энгстрем, 1994, с. 914; Ричардс, 1980, рис. 1
- ^ Шмидт и Энгстрем, 1994, с. 917
- ^ Массер и Карлтон, 2005, с. 1153
- ^ Хансон и др., 2010, с. 342
- ^ Jump up to: а б с Карлтон и Арройо-Кабралес, 2009, с. 114
- ^ Jump up to: а б Карлтон и Арройо-Кабралес, 2009, с. 122
- ^ Jump up to: а б с д Карлтон и Арройо-Кабралес, 2009 г., таблица 2.
- ^ Рэй, 1962, таблица 3.
- ^ Джонс и Энгстром, 1986, с. 13; Рид, 2009, с. 207
- ^ Санчес и др., 2001, таблица 1.
- ^ Jump up to: а б Вульф, 1982, с. 1
- ^ Гольдман, 1918, с. 19; Рид, 2009, с. 206; Карлтон и Арройо-Кабралес, 2009, с. 116
- ^ Гольдман, 1918, с. 19; Рид, 2009, с. 206; Карлтон и Арройо-Кабралес, 2009, с. 116; Санчес и др., 2001, с. 209
- ^ Векслер, 2006, стр. 59.
- ^ Векслер, 2006, стр. 58–59
- ^ Ray, 1962, plate V
- ^ Jump up to: а б Гольдман, 1918, с. 19; Карлтон и Арройо-Кабралес, 2009, с. 116
- ^ Гольдман, 1918, с. 19
- ^ Вулф, 1982, с. 1; Уитакер и Гамильтон, 1998, стр. 278–279.
- ^ Векслер, 2006, стр. 55–56
- ^ Векслер, 2006, стр. 56–57
- ^ Хупер и Массер, 1964, с. 13; Векслер, 2006, с. 57
- ^ Хупер и Массер, 1964, с. 7
- ^ Векслер, 2006, стр. 57.
- ^ Олстон, 1882, пластина 15.
- ^ Карлтон и Арройо-Кабралес, 2009, с. 116; Восс и Векслер, 2009, с. 73
- ^ Reid, 2009, p. 207; Kruchek, 2004, p. 269
- ^ Векслер, 2006, стр. 87–88
- ^ Векслер, 2006, стр. 88.
- ^ Линдси, 2008, с. 473
- ^ Рид, 2009, с. 205
- ^ Эшер и др., 1978, с. 556; Кук и др., 2001; Уитакер и Гамильтон, 1998, с. 279; Рид, 2009, с. 279
- ^ Рид, 2009, с. 207; Уитакер и Гамильтон, 1998, с. 279; Вульф, 1982, с. 4; Холл и Далквест, 1963, с. 289
- ^ Блох и Роуз, 2005, с. 303; Медельин и Медельин, 2006, с. 710
- ^ Джонс и Энгстром, 1986, с. 12; Медельин и Медельин, 2006, с. 710; Рид, 2009, с. 207; Уитакер и Гамильтон, 1998, с. 280; Вульф, 1982, с. 2; Линзи и Хаммерсон, 2008 г.
- ^ Медельин и Медельин, 2006, с. 710; Вульф, 1982, с. 2
- ^ Медельин и Медельин, 2006, с. 710; Рид, 2006, с. 207; Санчес и др., 2001, с. 211; Уитакер и Гамильтон, 1998, с. 280
- ^ Вулф, 1982, с. 2
- ^ Энтони, 1920, с. 166; Вудман, 1995, с. 1, таблица 1; Макфарлейн и Дебро, 2001, с. 182
- ^ Уитакер и Гамильтон, 1998, с. 281; Вега и др., 2004, с. 217
- ^ Холл и Далквест, 1963, с. 288; Экерлин, 2005, с. 155; Андервуд и др., 1986; Барнард и др., 1971, с. 1294
- ^ Вулф, 1982, с. 3
- ^ Санчес и др., 2001, с. 211
- ^ Морган, 1993, с. 239; Массер и Карлтон, 2005, с. 1152
- ^ Морган, 1993, с. 239; Рэй, 1962, с. 33–34; Карлтон и Арройо-Кабралес, 2009 г., стр. 101–1. 114–115
- ^ Санчес и др., 2001, стр. 205, 211; Векслер и Тимм, 2017 г.
- ^ Карлтон и Арройо-Кабралес, 2009, с. 115
- ^ Линзи и Хаммерсон, 2008; Линзи и др., 2016; Вега и др., 2004, с. 218; Шмидли и Дэвис, 2004, с. 281; Уитакер и Гамильтон, 1998, стр. 278–280; Хофманн и др., 1990, с. 162
- ^ Линзи и Хаммерсон, 2008; Линзи и др., 2016; Тимм и Рид, 2019 г.
- ^ Макинтайр и др., 2005, с. 1083
- ^ Милаццо и др., 2006; Чу и др., 2008 г.
Цитируемая литература
[ редактировать ]- Аллен, Дж. А. 1890. Заметки о коллекциях млекопитающих, сделанных в Центральной Америке и южной Мексике доктором Одли К. Буллером, с описаниями новых видов родов Vespertilio , Sciurus и Lepus . Бюллетень Американского музея естественной истории 3 (11): 175–194.
- Олстон, Э. Р. 1882. Биология Центральной Америки. Млекопитающие . Р. Х. Портер, 220 стр.
- Энтони, HE, 1920 г. Зоолог с Ямайки . Естественная история 20: 157–168.
- Бэрд, С.Ф. 1857. Млекопитающие: общий отчет о зоологии нескольких тихоокеанских железнодорожных маршрутов . Отчеты об исследованиях и изысканиях для определения наиболее практичного и экономичного маршрута железной дороги от реки Миссисипи до Тихого океана (исполнительный документ Сената 78, Вашингтон, округ Колумбия) 8 (1): 1–757.
- Барнард, В.П., Эрнст, Дж.В. и Стивенс, Р.О. 1971. Eimeria palustris sp. н. и Isospora hammondi sp. н. (Coccidia: Eimeriidae) от болотной рисовой крысы Oryzomys palustris (Harlan)] (требуется подписка). Журнал паразитологии 57(6):1293–1296 JSTOR 3277983 .
- Блох, К.П. и Роуз, Р.К. 2005. Динамика популяций Oryzomys palustris и Microtus pennsylvanicus в приливных болотах Вирджинии (требуется подписка). Северо-восточный натуралист 12 (3): 295–306 JSTOR 3858686. doi : 10.1656/1092-6194(2005)012[0295:PDOOPA 2.0.CO;2].
- Карлтон, доктор медицинских наук и Арройо-Кабралес, Дж. 2009. Обзор комплекса Oryzomys couesi (Rodentia: Cricetidae: Sigmodontinae) в Западной Мексике . Бюллетень Американского музея естественной истории 331: 94–127.
- Чу, Ю.-К., Оуэн, Р.Д., Санчес-Эрнандес, К., Ромеро-Альмарес, М. де Л. и Йонссон, CB 2008. Генетическая характеристика и филогения хантавируса из Западной Мексики (требуется подписка). Вирусные исследования 131:180–188.
- Черчер, К.С. 1984. Фаунистические корреляции плейстоценовых отложений в западной Канаде. стр. 145–158 в Махани, штат Вашингтон (ред.). Корреляция четвертичных хронологий . Норидж, Великобритания: Geo Books, 517 стр. ISBN 978-0-86094-172-9
- Кук, В.М., Тимм, Р.М. и Хайман, Д.Э. 2001. Способность плавать у трёх грызунов сухих лесов Коста-Рики . Revista de Biologia Tropical 49 (3–4): 1177–1181.
- Куэс, Э. 1874. Краткое описание мюридов Северной Америки . Труды Академии естественных наук Филадельфии 26: 173–196.
- Coues, E. 1877. Muridae . стр. 100-1 x+264 в Куес, Э. и Аллен, Дж. А. Монографии североамериканских грызунов. Отчет Геологической службы территорий США 11:xii+x+1091 стр.
- Куес, Э. 1890. Оризомис . п. 4164 в Уитни, штат Вашингтон (ред.). Словарь и циклопедия Century , Vol. V. Компания «Век».
- Экерлин, Р.П. 2005. Блохи (Siphonaptera) полуострова Юкатан (Кампече, Кинтана-Роо и Юкатан), Мексика . Карибский научный журнал 41 (1): 152–157.
- Эллерман, младший, 1941. Семейства и роды ныне живущих грызунов . Том II. Семейство Муриды . Лондон: напечатано по заказу попечителей Британского музея, 690 стр.
- Элиот, Д.Г. 1904. Наземные и морские млекопитающие Средней Америки и Вест-Индии . Полевой Колумбийский музей, Зоологическая серия 4 (1): i – xxi, 1–439.
- Эшер, Р.Дж., Вулф, Дж.Л. и Лэйн, Дж.Н. 1978. Плавательное поведение рисовых крыс ( Oryzomys palustris ) и хлопковых крыс ( Sigmodon hispidus ) (требуется подписка). Журнал маммологии 59(3):551–558 JSTOR 1380231. дои : 10.2307/1380231 .
- Гольдман, Е.А. 1918. Рисовые крысы Северной Америки . Североамериканская фауна 43: 1–100.
- Холл, ER и Далквест, WW 1963. Млекопитающие Веракруса . Публикации Канзасского университета, Музей естественной истории 14:165–362.
- Хэнсон, Дж.Д., Индорф, Дж.Л., Свир, В.Дж. и Брэдли, Р.Д. 2010. Молекулярная дивергенция внутри комплекса Oryzomys palustris : свидетельства существования нескольких видов. Журнал маммологии 91 (2): 336–347. два : 10.1644/08-МАММ-А-342.1 .
- Гершковитц, П. 1948. Млекопитающие северной Колумбии. Предварительное сообщение № 3: Водяные крысы (род Nектомии ) с дополнительными примечаниями о родственных формах . Труды Национального музея США 98: 49–56.
- Гершковиц, П. 1970. Дополнительные примечания к неотропическим Oryzomys dimidiatus и Oryzomys hammondi (Cricetinae) (требуется подписка). Журнал маммологии 51(4):789–794 JSTOR 1378303. дои : 10.2307/1378303 .
- Гершковиц, П. 1971. Новая рисовая крыса из группы Oryzomys palustris (Cricetinae, Muridae) из северо-западной Колумбии, с замечаниями по распространению (требуется подписка). Журнал маммологии 52(4):700–709 JSTOR 1378917. дои : 10.2307/1378917 .
- Хофманн Дж. Э., Гарднер Дж. Э. и Морис М. Дж. 1990. Распространение, численность и среда обитания болотной рисовой крысы ( Oryzomys palustris ) в южном Иллинойсе. Труды Академии наук штата Иллинойс 83 (3–4): 162–180.
- Хупер, Э.Т. и Массер, Г.Г., 1964. Головка полового члена неотропических крицитинов (семейство Muridae) с комментариями по классификации муроидных грызунов . Разные публикации Зоологического музея Мичиганского университета 123:1–57.
- Джонс, Дж. К. младший и Энгстрем, доктор медицинских наук, 1986. Краткий обзор рисовых крыс (род Oryzomys ) Никарагуа. Периодические статьи, Музей Техасского технологического университета, 103:1–23.
- Кручек, Б.Л. 2004. Использование болотной и горной среды обитания болотной рисовой крысой ( Oryzomys palustris ). Журнал маммологии 85(3):569–575 JSTOR 1383957. дои : 10.1644/BEH-016 .
- Линдсей, Э.Х. 2008. Cricetidae. стр. 456–479 в книге Дженис, К.М., Ганнелла, Г.Ф. и Уэна, доктора медицины (ред.). Эволюция третичных млекопитающих Северной Америки. Том 2: Мелкие млекопитающие, ксенартраны и морские млекопитающие . Издательство Кембриджского университета, 802 стр. ISBN 978-0-521-78117-6
- Линзи А.В. и Хаммерсон Г. 2008. Oryzomys palustris . В МСОП. Красный список исчезающих видов МСОП . Версия 2009.2. < www.iucnredlist.org >. Загружено 30 ноября 2009 г.
- Линзи А.В., Тимм Р., Вудман Н., Мэтсон Дж. и Самудио Р. 2016. Oryzomys couesi (версия с ошибками опубликована в 2017 г.). Красный список исчезающих видов МСОП 2016. doi : 10.2305/IUCN.UK.2016-3.RLTS.T15592A22388237.en .
- Макфарлейн Д.А. и Дебро А.О. 2001. Новый вид вымерших оризомииновых грызунов из четвертичного периода Кюрасао, Нидерландские Антильские острова . Карибский научный журнал 37 (3–4): 182–184.
- Макинтайр Н.Е., Чу Ю.-К., Оуэн Р.Д., Абузейне А., де ла Санча Н., Дик К.В., Холсомбак Т. Нисбетт Р.А. и Йонссон К. 2005. Продольное исследование Вирус Байу, хозяева и среда обитания . Американский журнал тропической медицины и гигиены 73: 1043–1049. ПМИД 16354810 .
- Медельин, XL и Медельин, РА, 2006. Oryzomys couesi (Alston, 1877). стр. 709–710 в Себальосе, Г. и Оливе, Г. (ред.). Дикие млекопитающие Мексики . Мехико: Национальная комиссия по изучению и использованию биоразнообразия и Фонд экономической культуры, 986 стр. ISBN 978-970-9000-30-6
- Мерриам, CH 1901. Краткий обзор рисовых крыс (род Oryzomys ) США и Мексики . Труды Вашингтонской академии наук 3: 273–295.
- Милаццо М.Л., Кахимат М.Н., Хэнсон Д.Д., Брэдли Р.Д., Кинтана М., Шерман К., Веласкес Р.Т. и Фулхорст К.Ф. 2006. Вирус катакамас, хантавирусный вид, естественным образом связанный с Oryzomys couesi (oryzomys Coues). ) в Гондурасе . Американский журнал тропической медицины и гигиены 75 (5): 1003–1010.
- Морган, Г.С. 1993. Четвертичные наземные позвоночные Ямайки. Мемуары Геологического общества Америки 182: 417–442.
- Массер, Г.Г. и Карлтон, доктор медицинских наук, 2005. Надсемейство Muroidea . стр. 894–1531 в Уилсоне, Д.Э. и Ридере, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник . 3-е изд . Балтимор: Издательство Университета Джона Хопкинса, 2 тома, 2142 стр. ISBN 978-0-8018-8221-0
- Пардиньяс, У. Ф. Дж., Д'Элия, Г. и Ортис, П. Е. 2002. Ископаемые сигмодонтины (Rodentia, Muroidea, Sigmodontinae) Южной Америки: современное состояние их знаний и перспективы . Неотропическая мастозоология 9 (2): 209–252.
- Рэй, CE 1962. Оризомииновые грызуны Антильского субрегиона. Докторская диссертация, Гарвардский университет, 211 стр.
- Рид, Ф. 2009. Полевой справочник по млекопитающим Центральной Америки и Юго-Восточной Мексики. 2-е издание . Издательство Оксфордского университета, США, 346 стр. ISBN 978-0-19-534322-9
- Ричардс, Р.Л., 1980. Останки рисовой крысы ( Oryzomys cf. palustris ) из пещер южной Индианы . Труды Академии наук Индианы 89: 425–431.
- Санчес Х., Дж., Очоа Г., Дж. и Восс, Р.С. 2001. Повторное открытие Oryzomys gogasi (Rodentia: Muridae) с примечаниями по таксономии и естественной истории (требуется подписка). Млекопитающие 65: 205–214. два : 10.1515/мамм.2001.65.2.205 .
- Шмидли, DJ и Дэвис, WB 2004. Млекопитающие Техаса. 2-е издание . Техасский университет Press, 501 стр. ISBN 978-0-292-70241-7
- Шмидт, Калифорния и Энгстрем, доктор медицинских наук, 1994. Генетическая изменчивость и систематика рисовых крыс ( группа видов Oryzomys palustris ) в южном Техасе и северо-восточном Тамаулипасе, Мексика. Журнал маммологии 75(4):914-928 JSTOR 1382473 дои : 10.2307/1382473 .
- Шварц, К.В. и Шварц, Э.Р. 2001. Дикие млекопитающие Миссури . Университет Миссури Press, 368 стр. ISBN 978-0-8262-1359-4
- Тейт, GHH 1932. Таксономическая история крицетидных грызунов Южной и Центральной Америки рода Oryzomys . Часть 1. Подрод Oryzomys . Новитаты Американского музея 579: 1–18.
- Тимм Р. и Рид Ф. 2019. Oryzomys dimidiatus . 2019 Красный список видов, находящихся под угрозой исчезновения МСОП . doi : 10.2305/IUCN.UK.2019-1.RLTS.T15594A22387864.en .
- Труессар, Э.Л. 1898. Каталог млекопитающих, как живых, так и ископаемых. Том 2 Берлин: Р. Фридлендер и Зон, 1469 стр. (на латыни).
- Андервуд, Х.Т., Оуэн, Дж.Г. и Энгстром, доктор медицинских наук, 1986. Эндогельминты трех видов Oryzomys (Rodentia: Cricetidae) из Сан-Луис-Потоси, Мексика (требуется подписка). Юго-западный натуралист 31(3):410–411 JSTOR 3671854 .
- Вега Р., Васкес-Домингес Э., Мехиа-Пуэнте А. и Куаро А.Д. 2004. Неожиданно высокий уровень генетической изменчивости и структура популяции островного эндемичного грызуна ( Oryzomys couesi cozumelae ) . Биологическая охрана 137: 210–222. doi : 10.1016/j.biocon.2007.02.007 .
- Восс, Р.С. и Векслер, М.В. 2009. О таксономическом статусе Oryzomys curasoae McFarlane и Debrot, 2001, (Rodentia: Cricetidae: Sigmodontinae) с замечаниями о филогенетических отношениях O. gogasi Hershkovitz, 1971 . Карибский научный журнал 45(1):73–79.
- Векслер, М. 2006. Филогенетические взаимоотношения оризомииновых грызунов (Muroidea: Sigmodontinae): отдельный и комбинированный анализ морфологических и молекулярных данных . Бюллетень Американского музея естественной истории 296: 1–149.
- Векслер М., Персекильо А.Р. и Восс Р.С. 2006. Десять новых родов оризомииновых грызунов (Cricetidae: Sigmodontinae) . Новитаты Американского музея 3537: 1–29.
- Векслер М. и Тимм Р. 2017. Oryzomys gogasi (версия с ошибками опубликована в 2018 году). Красный список исчезающих видов МСОП 2017. doi : 10.2305/IUCN.UK.2017-3.RLTS.T115554360A22387922.en .
- Уитакер, Дж. О. и Гамильтон, У. Дж. 1998. Млекопитающие восточной части Соединенных Штатов . Издательство Корнельского университета, 583 стр. ISBN 978-0-8014-3475-4
- Вулф, Дж. Л. 1982. Oryzomys palustris . Виды млекопитающих 176:1–5. дои : 10.2307/3503921 .
- Вудман, Н. 1995. Морфологические различия между плейстоценовыми и современными образцами Cryptotis (Insectivora: Soricidae) с полуострова Юкатан, Мексика. Журнал маммологии 76(1):223–231 JSTOR 1382330. дои : 10.2307/1382330 .
| Базы данных органов управления : Национальные |
|---|