Кариотип
Кариотип — это общий вид полного набора хромосом в клетках вида или отдельного организма, в основном включая их размеры, количество и форму. [1] [2] Кариотипирование — это процесс, посредством которого распознается кариотип путем определения хромосомного набора человека, включая количество хромосом и любые аномалии.

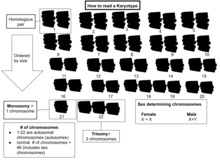
Кариограмма . или идиограмма — это графическое изображение кариотипа, при котором хромосомы обычно организованы парами, упорядоченными по размеру и положению центромеры для хромосом одинакового размера Кариотипирование обычно сочетает в себе световую микроскопию и фотографию в метафазе клеточного цикла и приводит к фотомикрографической (или просто микрографической) кариограмме. Напротив, схематическая кариограмма представляет собой разработанное графическое изображение кариотипа. На схематических кариограммах для краткости обычно отображается только одна из сестринских хроматид каждой хромосомы, а на самом деле они обычно расположены настолько близко друг к другу, что и на микрофотографиях выглядят как одна, если разрешение не достаточно высокое, чтобы их различить. Изучение целых наборов хромосом иногда называют кариологией .
Кариотипы описывают количество хромосом организма и то, как эти хромосомы выглядят под световым микроскопом . Внимание уделяется их длине, положению центромер , характеру полос, любым различиям между половыми хромосомами и любым другим физическим характеристикам. [3] Получение и изучение кариотипов является частью цитогенетики .
Основное число хромосом в соматических клетках особи или вида называется соматическим числом и обозначается 2n . В зародышевой линии (половых клетках) число хромосом равно n (у человека: n = 23). [4] [5] стр. 28 Таким образом, у человека 2n = 46.
Так, у нормальных диплоидных организмов аутосомные хромосомы присутствуют в двух копиях. могут быть, а могут и не быть Половые хромосомы . Полиплоидные клетки имеют несколько копий хромосом, а гаплоидные клетки имеют единичные копии.
Кариотипы можно использовать для многих целей; например, для изучения хромосомных аберраций , клеточных функций, таксономических отношений, медицины и сбора информации о прошлых эволюционных событиях ( кариосистематика ). [6]
Наблюдения за кариотипами [ править ]


Окрашивание [ править ]
Изучение кариотипов возможно с помощью окрашивания . Обычно подходящий краситель , например Гимза , [8] применяется после того, как клетки были остановлены во время клеточного деления раствором колхицина, обычно в метафазе или прометафазе , когда они наиболее конденсированы. Для того чтобы пятно Гимзы прикрепилось правильно, все хромосомные белки должны быть переварены и удалены. Для людей чаще всего используются лейкоциты , поскольку их легко заставить делиться и расти в культуре ткани . [9] Иногда наблюдения можно проводить на неделящихся ( интерфазных ) клетках. Пол будущего плода можно предсказать, наблюдая за интерфазными клетками (см. амниотический центез и тельца Барра ).
Наблюдения [ править ]
Обычно наблюдают и сравнивают шесть различных характеристик кариотипов: [10]
- Различия в абсолютных размерах хромосом. Хромосомы могут различаться по абсолютному размеру до двадцати раз между родами одного и того же семейства. Например, бобовые Lotus tenuis и Vicia faba имеют по шесть пар хромосом, однако хромосомы V. faba во много раз больше. Эти различия, вероятно, отражают разное количество дупликаций ДНК.
- Различия в положении центромер . Эти различия, вероятно, возникли за счет транслокаций .
- Различия в относительных размерах хромосом. Эти различия, вероятно, возникли из-за чередования сегментов неравной длины.
- Различия в основном числе хромосом. Эти различия могли возникнуть в результате последовательных неравных транслокаций, которые удалили весь необходимый генетический материал из хромосомы, что привело к его потере без ущерба для организма (гипотеза дислокации) или в результате слияния. У человека на одну пару хромосом меньше, чем у человекообразных обезьян. Человеческая хромосома 2, по-видимому, возникла в результате слияния двух предковых хромосом, и многие гены этих двух исходных хромосом были транслоцированы в другие хромосомы.
- Различия в количестве и положении спутников. Сателлиты — небольшие тельца, прикрепленные к хромосоме тонкой нитью.
- Различия в степени и распределении содержания ГК ( Гуанин - Цитозин пары и Аденин - Тимин ). В метафазе, когда обычно изучают кариотип, вся ДНК конденсируется, но большую часть времени ДНК с высоким содержанием GC обычно менее конденсирована, то есть имеет тенденцию проявляться в виде эухроматина, а не гетерохроматина . ДНК, богатая GC, имеет тенденцию содержать больше кодирующей ДНК и быть более транскрипционно активной . [11] ДНК, богатая GC, светлее при окрашивании по Гимзе . [12] Области эухроматина содержат большее количество пар гуанин - цитозин (т. е. имеют более высокое содержание GC ). Техника окрашивания с использованием окрашивания по Гимзе называется G-полосами и, следовательно, дает типичные «G-полосы». [12]
Таким образом, полное описание кариотипа может включать количество, тип, форму и полосатость хромосом, а также другую цитогенетическую информацию.
Часто встречаются вариации:
- между полами,
- между зародышевой линией и сомой (между гаметами и остальным телом),
- между членами популяции ( хромосомный полиморфизм ),
- по географической специализации и
- в мозаиках или иным образом ненормальных особях. [13]
Кариограмма человека [ править ]
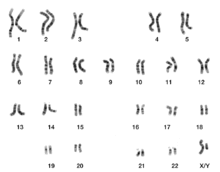
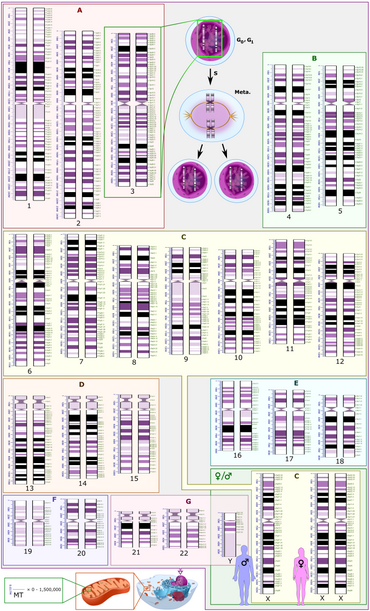
Как микрографические, так и схематические кариограммы, показанные в этом разделе, имеют стандартное расположение хромосом и отображают более темные и светлые области, как видно на G-диапазоне , который представляет собой внешний вид хромосом после обработки трипсином (для частичного переваривания хромосом) и окрашивания по Гимзе. пятно . По сравнению с более темными областями, более светлые области, как правило, более транскрипционно активны, с большим соотношением кодирующей ДНК по сравнению с некодирующей ДНК и более высоким содержанием GC . [11]
Как на микрографических, так и на схематических кариограммах показан нормальный диплоидный кариотип человека, который представляет собой типичный состав генома нормальной клетки человеческого организма и содержит 22 пары аутосомных хромосом и одну пару половых хромосом (аллосом). Серьезным исключением из диплоидии у человека являются гаметы (сперматозоиды и яйцеклетки), которые гаплоидны с 23 непарными хромосомами, и эта плоидность не показана на этих кариограммах. Микрографическая кариограмма преобразуется в оттенки серого , тогда как схематическая кариограмма показывает фиолетовый оттенок, который обычно наблюдается при окраске по Гимзе (и является результатом его лазурного компонента B, который окрашивает ДНК в фиолетовый цвет). [14]
Схематическая кариограмма в этом разделе представляет собой графическое изображение идеализированного кариотипа. Для каждой пары хромосом на шкале слева указана длина в миллионах пар оснований , а на шкале справа показаны обозначения полос и подполос . Такие полосы и поддиапазоны используются Международной системой цитогеномной номенклатуры человека для описания мест хромосомных аномалий . Каждый ряд хромосом вертикально выровнен на уровне центромеры .
Группы хромосом человека [ править ]
На основании особенностей кариограммы размера, положения центромеры и иногда наличия хромосомного сателлита (сегмента, дистального вторичной перетяжки ), хромосомы человека классифицируются на следующие группы: [15]
| Группа | Хромосомы | Функции |
|---|---|---|
| А | 1-3 | Большие, метацентрические или субметацентрические |
| Б | 4-5 | Большой, субметацентрический |
| С | 6-12, Х | Среднего размера, субметацентрический |
| Д | 13-15 | Среднего размера, акроцентрическая, со сателлитом. |
| И | 16-18 | Маленькие, метацентрические или субметацентрические |
| Ф | 19-20 | Очень маленький, метацентрический |
| Г | 21-22 и | Очень маленький, акроцентрический (и 21, 22 со сателлитом ) |
Альтернативно, геном человека можно классифицировать следующим образом на основе парности, половых различий, а также местоположения внутри ядра клетки по сравнению с внутри митохондрий :
- 22 пары гомологичных аутосомных хромосом (хромосомы с 1 по 22). Гомологичность означает, что они имеют одинаковые гены в одних и тех же локусах, а аутосомность означает, что они не являются половыми хромомами.
- Две половые хромосомы (в зеленом прямоугольнике внизу справа на схематической кариограмме с соседними силуэтами типичных репрезентативных фенотипов ): Наиболее распространенные кариотипы у женщин содержат две Х-хромосомы и обозначаются 46,XX; мужчины обычно имеют как X, так и Y-хромосому, обозначаемую 46,XY. Однако примерно 0,018% людей являются интерсексуалами , иногда из-за различий в половых хромосомах. [16]
- Митохондриальный геном человека (показан внизу слева на схематической кариограмме в масштабе по сравнению с ядерной ДНК с точки зрения пар оснований ), хотя в клинической практике он не включается в микрографические кариограммы. Его геном относительно мал по сравнению с остальными.
Номер копии [ изменить ]
схематических кариограммах обычно отображается число копий ДНК, соответствующее фазе клеточного На G0 состояния (вне репликативного клеточного цикла ), которая является наиболее распространенным состоянием клеток. Схематическая кариограмма в этом разделе также показывает это состояние. В этом состоянии (а также во время фазы G1 клеточного цикла ) каждая клетка имеет по две аутосомные хромосомы каждого вида (обозначенные 2n), где каждая хромосома имеет по одной копии каждого локуса , что составляет общее число копий 2 для каждый локус (2c). Вверху в центре схематической кариограммы также показана пара хромосом 3 после синтеза ДНК , происходящего в S-фазе (обозначенной как S) клеточного цикла. Этот интервал включает G 2 фазу и метафазу (обозначенную как «Мета»). В течение этого интервала все еще существует 2n, но каждая хромосома будет иметь по 2 копии каждого локуса, при этом каждая сестринская хроматида (плечо хромосомы) соединена в центромере, всего 4c. [17] Хромосомы на микрографических кариограммах также находятся в этом состоянии, поскольку их обычно микрографируют в метафазе, но на этом этапе две копии каждой хромосомы расположены настолько близко друг к другу, что кажутся едиными, если разрешение изображения не достаточно высокое, чтобы их можно было различить. их. В действительности во время фаз G0 и G1 ядерная ДНК рассредоточена в виде хроматина и не показывает визуально различимых хромосом даже при микрографии.
Число копий митохондриального генома человека на клетку человека варьируется от 0 (эритроциты). [18] до 1 500 000 ( ооцитов ), главным образом, в зависимости от количества митохондрий в клетке. [19]
Разнообразие и эволюция кариотипов [ править ]
Хотя репликация и транскрипция ДНК . высоко стандартизированы у эукариот , этого нельзя сказать об их кариотипах, которые весьма изменчивы Между видами существуют различия в числе хромосом и в детальной организации, несмотря на то, что они построены из одних и тех же макромолекул . Эта вариация обеспечивает основу для ряда исследований в области эволюционной цитологии .
В некоторых случаях наблюдаются даже значительные различия внутри вида. В обзоре Годфри и Мастерс заключают:
По нашему мнению, маловероятно, что тот или иной процесс может независимо объяснить широкий спектр наблюдаемых структур кариотипа... Но в сочетании с другими филогенетическими данными кариотипическое деление может помочь объяснить резкие различия в количестве диплоидов. между близкородственными видами, что ранее было необъяснимо. [20]
Хотя о кариотипах на описательном уровне известно многое, и ясно, что изменения в организации кариотипов оказали влияние на эволюционный ход многих видов, совершенно неясно, каково может быть общее значение.
У нас очень плохое понимание причин эволюции кариотипа, несмотря на многие тщательные исследования... общее значение эволюции кариотипа неясно.
— Мейнард Смит [21]
Изменения в ходе разработки [ править ]
Вместо обычной репрессии генов некоторые организмы занимаются крупномасштабной элиминацией гетерохроматина или другими видами видимой корректировки кариотипа.
- Удаление хромосом. У некоторых видов, как и у многих сциарид , во время развития удаляются целые хромосомы. [22]
- Уменьшение хроматина (отец-основатель: Теодор Бовери ). В ходе этого процесса, обнаруженного у некоторых копепод и круглых червей, таких как Ascaris suum , части хромосом отбрасываются в определенных клетках. Этот процесс представляет собой тщательно организованную перестройку генома, при которой строятся новые теломеры и теряются определенные участки гетерохроматина. [23] [24] У A. suum все предшественники соматических клеток подвергаются уменьшению хроматина. [25]
- Х-инактивация . Инактивация одной Х-хромосомы происходит на ранних стадиях развития млекопитающих (см. « Тело Барра и компенсация дозы »). У плацентарных млекопитающих инактивация случайна между двумя X; таким образом, самка млекопитающего представляет собой мозаику в отношении своих Х-хромосом. У сумчатых всегда инактивирован отцовский X. У женщин около 15% соматических клеток избегают инактивации. [26] а количество генов, затронутых инактивированной Х-хромосомой, варьируется между клетками: в клетках фибробластов около 25% генов в теле Барра избегают инактивации. [27]
Число хромосом в наборе [ править ]
Ярким примером изменчивости между близкородственными видами является мунтжак , которого исследовали Курт Бениршке и Дорис Вурстер . Диплоидное число китайского мунтжака, Muntiacus reevesi , составило 46, все телоцентрические . Когда они изучили кариотип близкородственного индийского мунтжака, Muntiacus muntjak , они были удивлены, обнаружив, что у него женское = 6, мужское = 7 хромосом. [28]
Они просто не могли поверить в то, что видели... Они молчали два-три года, потому что думали, что с их культурой тканей что-то не так... Но когда они получили ещё пару образцов, они подтвердили [свои выводы].
- Сюй п. 73-4 [29]
Число хромосом в кариотипе между (относительно) неродственными видами сильно различается. Низкий рекорд принадлежит нематоде Parascaris univalens , у которой гаплоид n = 1; и муравей: Myrmecia pilosula . [30] Самый высокий рекорд будет где-то среди папоротников , впереди будет папоротник змеиный язык Ophioglossum со средним числом 1262 хромосомы. [31] Наивысшую оценку среди животных может получить коротконосый осетр Acipenser brevirostrum с 372 хромосомами. [32] Существование дополнительных или B-хромосом означает, что число хромосом может варьироваться даже в пределах одной скрещивающейся популяции; и анеуплоиды являются еще одним примером, хотя в этом случае они не будут считаться нормальными членами популяции.
Фундаментальное число [ править ]
Фундаментальное число FN — это количество видимых основных хромосомных плеч на набор хромосом. кариотипа [33] [34] Таким образом, FN ≤ 2 x 2n, разница зависит от количества хромосом, считающихся одноплечими ( акроцентрическими или телоцентрическими ). У человека ФН = 82, [35] за счет наличия пяти пар акроцентрических хромосом: 13 , 14 , 15 , 21 и 22 человека ( Y-хромосома также акроцентрична). Фундаментальное аутосомное число или аутосомное фундаментальное число, FNa. [36] или АН , [37] Кариотип – это количество видимых основных хромосомных плеч на набор аутосом (несцепленных с полом хромосом ).
Плоидность [ править ]
Плоидность – это количество полных наборов хромосом в клетке.
- Полиплоидия , при которой в клетках имеется более двух наборов гомологичных хромосом, встречается преимущественно у растений. это имело большое значение в эволюции растений По мнению Стеббинса, . [38] [39] [40] [41] Доля цветковых растений, являющихся полиплоидами, оценивалась Стеббинсом в 30–35%, но у трав этот средний показатель намного выше, около 70%. [42] Полиплоидия у низших растений ( папоротников , хвощей и psilotales ) также распространена, и некоторые виды папоротников достигли уровней полиплоидии, намного превышающих самые высокие уровни, известные у цветковых растений. Полиплоидия у животных встречается гораздо реже, но в некоторых группах она значительна. [43]
Полиплоидные серии родственных видов, которые полностью состоят из кратных одному основному числу, известны как эуплоидные .
- Гаплодиплоидия , при которой один пол диплоидный , а другой гаплоидный . Это обычное явление у перепончатокрылых и некоторых других групп.
- Эндополиплоидия возникает, когда во взрослых дифференцированных тканях клетки перестают делиться митозом , но ядра содержат большее, чем исходное соматическое число хромосом . [44] В эндоцикле ( эндомитоз или эндоредупликация ) хромосомы в «покоящемся» ядре подвергаются редупликации , при этом дочерние хромосомы отделяются друг от друга внутри интактной ядерной мембраны . [45]
Во многих случаях эндополиплоидные ядра содержат десятки тысяч хромосом (точно сосчитать которые невозможно). Клетки не всегда содержат точные кратные (степени двойки), поэтому простое определение «увеличение числа хромосомных наборов, вызванное репликацией без деления клетки» не совсем точно.
Этот процесс (особенно изученный у насекомых и некоторых высших растений, таких как кукуруза) может быть стратегией развития, направленной на увеличение продуктивности тканей, которые высокоактивны в биосинтезе. [46]
Это явление спорадически встречается во всем царстве эукариот , от простейших до человека; он разнообразен и сложен и служит дифференцировке и морфогенезу . во многих отношениях [47] - См. Палеополиплоидию для изучения дупликаций древних кариотипов.
Анеуплоидия [ править ]
Анеуплоидия — это состояние, при котором число хромосом в клетках не соответствует типичному для данного вида числу. Это может привести к хромосомной аномалии , такой как дополнительная хромосома или потеря одной или нескольких хромосом. Аномалии числа хромосом обычно вызывают дефект в развитии. синдром Дауна и синдром Тернера Примерами этого являются .
Анеуплоидия также может возникать в группе близкородственных видов. Классическими примерами растений являются род Crepis , где гаметические (= гаплоидные) числа образуют ряд x = 3, 4, 5, 6 и 7; и Crocus , где каждое число от x = 3 до x = 15 представлено хотя бы одним видом. Различные данные показывают, что тенденции эволюции в разных группах шли в разных направлениях. [48] У приматов человекообразные обезьяны имеют хромосомы 24x2, тогда как у человека — 23x2. Человеческая хромосома 2 образовалась в результате слияния предковых хромосом, уменьшив их количество. [49]
Хромосомный полиморфизм [ править ]
Некоторые виды полиморфны по разным структурным формам хромосом. [50] Структурная изменчивость может быть связана с разным числом хромосом у разных особей, что встречается у рыльца божьей коровки Chilocorus , некоторых богомолов рода Ameles , [51] Европейская землеройка Sorex araneus . [52] На примере моллюска Thais lapillus ( собачьего трубача ) на побережье Бретани есть некоторые свидетельства того, что две хромосомные морфы адаптированы к разным средам обитания. [53]
Деревья видов [ править ]
Детальное изучение полосатости хромосом у насекомых с политенными хромосомами может выявить взаимоотношения между близкородственными видами: классическим примером является исследование полосатости хромосом у гавайских дрозофилид, проведенное Хэмптоном Л. Карсоном .
Примерно на 6500 квадратных миль (17 000 км²) 2 ), Гавайские острова обладают самой разнообразной в мире коллекцией дрозофилидных мух, обитающих от тропических лесов до субальпийских лугов . Эти примерно 800 гавайских видов дрозофилид обычно относят к двум родам, Drosophila и Scaptomyza , семейства Drosophilaidae .
Политенные полосы группы «картинных крыльев», наиболее изученной группы гавайских дрозофилид, позволили Карсону разработать эволюционное древо задолго до того, как анализ генома стал практически осуществим. В каком-то смысле расположение генов можно увидеть в характере полос каждой хромосомы. Хромосомные перестройки, особенно инверсии , позволяют увидеть, какие виды являются близкими.
Результаты очевидны. Инверсии, построенные в виде дерева (и независимо от какой-либо другой информации), показывают четкий «поток» видов от старых островов к новым. Бывают также случаи обратной колонизации старых островов и пропуска островов, но они встречаются гораздо реже. Используя K-Ar датирование, современные острова датируются периодом от 0,4 миллиона лет назад ( Мауна-Кеа ) до 10 млн лет назад ( Неккер ). Самый старый член Гавайского архипелага, все еще находящийся над морем, — это атолл Куре , возраст которого можно датировать 30 млн лет назад. Сам архипелаг (образованный Тихоокеанской плитой, движущейся над горячей точкой ) существовал гораздо дольше, по крайней мере, в меловой период . Предыдущие острова, ныне находящиеся под водой ( гайоты ), образуют цепь Императорских подводных гор . [54]
Все местные виды Drosophila и Scaptomyza на Гавайях, по-видимому, произошли от одного предкового вида, колонизировавшего острова, вероятно, 20 миллионов лет назад. Последующее адаптивное излучение было вызвано отсутствием конкуренции и широким разнообразием ниш . самка могла бы Хотя одна беременная колонизировать остров, более вероятно, что это была группа одного и того же вида. [55] [56] [57] [58]
На Гавайском архипелаге есть и другие животные и растения, которые подверглись похожему, хотя и менее впечатляющему, адаптивному излучению. [59] [60]
Объединение хромосом [ править ]
Хромосомы при обработке некоторыми красителями приобретают полосатый рисунок. Полосы представляют собой чередующиеся светлые и темные полосы, возникающие по всей длине хромосом. Уникальные образцы полос используются для идентификации хромосом и диагностики хромосомных аберраций, включая поломку хромосом, потерю, дупликацию, транслокацию или инвертированные сегменты. Ряд различных обработок хромосом создает ряд паттернов полос: G-диапазоны, R-диапазоны, C-диапазоны, Q-диапазоны, Т-диапазоны и NOR-диапазоны.
Изображение кариотипов [ править ]
Виды бандажирования [ править ]
Цитогенетика использует несколько методов для визуализации различных аспектов хромосом: [9]
- G-бэндинг получают с помощью окраски по Гимзе после расщепления хромосом трипсином . Он дает серию светло- и темно окрашенных полос — темные области, как правило, гетерохроматические, поздно реплицирующиеся и богатые АТ. Светлые области, как правило, эухроматические, рано реплицирующиеся и богатые GC. Этот метод обычно позволяет получить 300–400 полос в нормальном человеческом геноме . Это наиболее распространенный метод объединения хромосом. [61]
- R-диапазон является противоположностью G-диапазона (R означает «обратный»). Темные области являются эухроматическими (области, богатые гуанином-цитозином), а светлые области - гетерохроматическими (области, богатые тимином-аденином).
- C-бэндинг: Гимза связывается с конститутивным гетерохроматином , поэтому окрашивает центромеры . Название происходит от центромерного или конститутивного гетерохроматина. Перед окрашиванием препараты подвергаются щелочной денатурации, что приводит к практически полному депуринированию ДНК. После промывки зонда оставшуюся ДНК снова ренатурируют и окрашивают раствором Гимзы, состоящим из метиленовой лазури, метиленвиолета, метиленового синего и эозина. Гетерохроматин связывает большую часть красителя, тогда как остальные хромосомы поглощают лишь его небольшую часть. С-связь оказалась особенно подходящей для характеристики хромосом растений.
- Q-бэндинг представляет собой флуоресцентный рисунок, полученный с использованием хинакрина для окрашивания. Рисунок полос очень похож на тот, который наблюдается при G-banding. Их можно узнать по желтой флуоресценции разной интенсивности. Большая часть окрашенной ДНК представляет собой гетерохроматин. Хинакрин (атебрин) связывает обе области, богатые АТ и GC, но флуоресцирует только комплекс АТ-хинакрин. Поскольку регионы, богатые АТ, чаще встречаются в гетерохроматине, чем в эухроматине, эти регионы маркируются предпочтительно. Разная интенсивность отдельных полос отражает различное содержание АТ. Другие флуорохромы, такие как DAPI или Hoechst 33258, также приводят к характерным воспроизводимым рисункам. Каждый из них производит свой определенный узор. Другими словами: свойства связей и специфичность флуорохромов основаны не только на их сродстве к областям, богатым АТ. Скорее, распределение АТ и ассоциация АТ с другими молекулами, такими как, например, гистоны, влияют на связывающие свойства флуорохромов.
- Т-образное соединение: визуализируйте теломеры .
- Окрашивание серебром: нитрат серебра окрашивает белок, связанный с областью ядрышковой организации . Это дает темную область, где откладывается серебро, что указывает на активность генов рРНК в NOR.
Классическая кариотипа цитогенетика

В «классическом» (изображенном) кариотипе краситель , чаще Гимза (G-бэндинг) , реже мепакрин (хинакрин) для окрашивания полос на хромосомах используется . Гимза специфичен для фосфатных групп ДНК . Хинакрин связывается с областями, богатыми аденином и тимином . Каждая хромосома имеет характерный рисунок полос, который помогает их идентифицировать; обе хромосомы в паре будут иметь одинаковый рисунок полос.
Кариотипы расположены так, что короткое плечо хромосомы находится вверху, а длинное плечо — внизу. Некоторые кариотипы называют короткое и длинное плечи p и q соответственно. Кроме того, по-разному окрашенным участкам и субобластям присвоены числовые обозначения от проксимального к дистальному плечу хромосомы. Например, синдром крика дю чата предполагает делецию короткого плеча 5-й хромосомы. Она записывается как 46,XX,5p-. Критической областью этого синдрома является делеция p15.2 ( локуса на хромосоме), которая записывается как 46,XX,del(5)(p15.2). [62]
Многоцветная FISH (mFISH) и спектральный кариотип (техника SKY) [ править ]

Многоцветная FISH и более раннее спектральное кариотипирование — это молекулярные цитогенетические методы, используемые для одновременной визуализации всех пар хромосом в организме в разных цветах. Флуоресцентно- меченные зонды для каждой хромосомы изготавливаются путем мечения ДНК, специфичной для хромосомы, различными флуорофорами . Поскольку существует ограниченное количество спектрально различных флуорофоров, для получения множества различных цветов используется метод комбинаторной маркировки. Комбинации флуорофоров улавливаются и анализируются с помощью флуоресцентного микроскопа с использованием до 7 узкополосных флуоресцентных фильтров или, в случае спектрального кариотипирования, с помощью интерферометра, прикрепленного к флуоресцентному микроскопу. В случае изображения mFISH каждая комбинация флуорохромов из полученных исходных изображений заменяется псевдоцветом в специальном программном обеспечении для анализа изображений. Таким образом, хромосомы или участки хромосом можно визуализировать и идентифицировать, что позволяет проводить анализ хромосомных перестроек. [63] В случае спектрального кариотипирования программное обеспечение для обработки изображений присваивает псевдоцвет каждой спектрально отличной комбинации, позволяя визуализировать индивидуально окрашенные хромосомы. [64]

Многоцветная FISH используется для выявления структурных хромосомных аберраций в раковых клетках и других болезненных состояниях, когда определение полос Гимзы или другие методы недостаточно точны.
Цифровое кариотипирование
Цифровое кариотипирование — это метод, используемый для количественного определения количества копий ДНК в геномном масштабе. Короткие последовательности ДНК из определенных локусов по всему геному выделяют и подсчитывают. [65] Этот метод также известен как виртуальное кариотипирование . Используя этот метод, можно обнаружить небольшие изменения в геноме человека, которые невозможно обнаружить методами, использующими метафазные хромосомы. Известно, что делеции некоторых локусов связаны с развитием рака. Такие делеции обнаруживаются посредством цифрового кариотипирования с использованием локусов, связанных с развитием рака. [66]
Хромосомные аномалии [ править ]
Хромосомные аномалии могут быть числовыми, например, при наличии дополнительных или отсутствующих хромосом, или структурными, как при производных хромосомах , транслокациях , инверсиях , крупномасштабных делециях или дупликациях. Численные аномалии, также известные как анеуплоидия , часто возникают в результате нерасхождения во время мейоза при образовании гаметы ; трисомии , при которых присутствуют три копии хромосомы вместо обычных двух, являются частыми численными аномалиями. Структурные аномалии часто возникают в результате ошибок гомологичной рекомбинации . Оба типа аномалий могут возникать в гаметах и, следовательно, присутствовать во всех клетках тела больного человека, или они могут возникнуть во время митоза и привести к возникновению генетической мозаики индивидуума, в котором есть как нормальные, так и аномальные клетки.
У людей [ править ]
Хромосомные аномалии, которые приводят к заболеваниям у человека, включают:
- Синдром Тернера возникает в результате наличия единственной Х-хромосомы (45,X или 45,X0).
- Синдром Клайнфельтера , наиболее распространенное мужское хромосомное заболевание, также известное как 47,XXY, вызвано лишней Х- хромосомой.
- Синдром Эдвардса вызван трисомией (три копии) хромосомы 18.
- Синдром Дауна — распространенное хромосомное заболевание, вызванное трисомией 21 хромосомы.
- Синдром Патау обусловлен трисомией 13-й хромосомы.
- Трисомия 9 , которая считается четвертой по распространенности трисомией, встречается у многих долгоживущих людей, но только в форме, отличной от полной трисомии, такой как синдром трисомии 9p или мозаичная трисомия 9. Они часто функционируют довольно хорошо, но, как правило, имеют проблемы. с речью.
- Также задокументированы трисомия 8 и трисомия 16, хотя они обычно не доживают до рождения.
Некоторые расстройства возникают в результате потери всего лишь участка одной хромосомы, в том числе
- Cri du Chat (крик кошки) происходит от усеченного короткого плеча на хромосоме 5. Название происходит от характерного крика младенцев, вызванного аномальным формированием гортани.
- 1p36 Делеционный синдром , возникающий из-за потери части короткого плеча хромосомы 1.
- синдром Ангельмана – в 50% случаев отсутствует сегмент длинного плеча 15-й хромосомы; делеция материнских генов, пример нарушения импринтинга .
- синдром Прадера-Вилли – в 50% случаев отсутствует сегмент длинного плеча 15-й хромосомы; делеция отцовских генов, пример нарушения импринтинга.
- Хромосомные аномалии также могут возникать в раковых клетках генетически нормального человека; Одним из хорошо документированных примеров является Филадельфийская хромосома , транслокационная мутация, обычно связанная с хроническим миелогенным лейкозом и реже с острым лимфобластным лейкозом .
История изучения кариотипа [ править ]
Хромосомы были впервые обнаружены в растительных клетках Карлом Вильгельмом фон Нэгели в 1842 году. Их поведение в клетках животных ( саламандры ) было описано Вальтером Флеммингом , первооткрывателем митоза , в 1882 году. Название было придумано другим немецким анатомом, Генрихом фон Вальдейером в 1882 году. 1888. Это неолатинское происхождение от древнегреческого κάρυον karyon , «ядро», «семя» или «ядро», и τύπος typos , «общая форма»).
Следующий этап наступил после развития генетики в начале 20 века, когда было признано, что хромосомы (которые можно наблюдать по кариотипу) являются носителями генов. Термин «кариотип», определяемый фенотипическим внешним видом соматических хромосом, в отличие от их генного содержания, был введен Григорием Левицким, работавшим со Львом Делоне, Сергеем Навашиным и Николаем Вавиловым . [67] [68] [69] [70] Последующую историю концепции можно проследить в работах К. Д. Дарлингтона. [71] и Майкл Джей Ди Уайт . [4] [13]
Исследования кариотипа человека заняли много лет, чтобы решить самый основной вопрос: сколько хромосом содержит нормальная диплоидная клетка человека? [72] В 1912 году Ганс фон Винивартер сообщил о 47 хромосомах в сперматогониях и 48 в оогониях , сделав вывод о XX/XO механизме определения пола . [73] Пейнтер в 1922 году не был уверен, был ли диплоид человека 46 или 48, сначала отдавая предпочтение 46, [74] но пересмотрел свое мнение с 46 на 48 и правильно настаивал на том, что у людей есть система XX/XY . [75] Учитывая методы того времени, эти результаты были поразительными.

Джо Хин Тджио работает в Альберта Левана лаборатории [76] обнаружил, что количество хромосом составляет 46, используя новые методы, доступные в то время:
- Использование клеток в культуре тканей
- Предварительная обработка клеток гипотоническим раствором , который набухает и распределяет хромосомы.
- Задержка митоза в метафазе раствором колхицина
- Сдавливание препарата на предметном стекле, выравнивание хромосом в одной плоскости
- Разрезание микрофотографии и преобразование результата в бесспорную кариограмму.
Работа состоялась в 1955 году, а опубликована в 1956 году. Кариотип человека включает всего 46 хромосом. [77] [29] У других человекообразных обезьян 48 хромосом. Теперь известно, что вторая хромосома человека является результатом слияния конца в конец двух хромосом предков обезьяны. [78] [79]
См. также [ править ]
- Цитогенетические обозначения - символы и сокращения, используемые в цитогенетике.
- Экран генома — лабораторный процесс.
Ссылки [ править ]
- ^ «Кариотип, определение» . Словарь английского языка Коллинза . Проверено 23 декабря 2022 г.
- ^ Джадд, Уолтер С.; Кэмпбелл, Кристофер С.; Келлог, Элизабет А.; Стивенс, Питер Ф.; Донохью, Майкл Дж. (2002). Систематика растений, филогенетический подход (2-е изд.). Сандерленд, Массачусетс, США: Sinauer Associates Inc., с. 544. ИСБН 0-87893-403-0 .
- ^ Кинг, RC; Стэнсфилд, штат Вашингтон; Маллиган, ПК (2006). Генетический словарь (7-е изд.). Издательство Оксфордского университета. п. 242 .
- ↑ Перейти обратно: Перейти обратно: а б Уайт 1973 , с. 35
- ^ Стеббинс, Г.Л. (1950). «Глава XII: Кариотип». Изменчивость и эволюция растений . Издательство Колумбийского университета. ISBN 9780231017336 .
- ^ «Кариосистематика» .
- ^ Ли М. Сильвер (1995). Генетика мышей, концепции и приложения. Глава 5.2: КАРИОТИПЫ, ХРОМОСОМЫ И ТРАНСЛОКАЦИИ . Издательство Оксфордского университета. Пересмотрено в августе 2004 г., январе 2008 г.
- ^ Препарат, в состав которого входят красители метиленовый синий, эозин Y и лазурь-A,B,C.
- ↑ Перейти обратно: Перейти обратно: а б Густашоу К.М. 1991. Хромосомные пятна. В Лабораторном руководстве по цитогенетике ACT , 2-е изд., изд. М. Дж. Барч. Ассоциация цитогенетических технологов, Raven Press, Нью-Йорк.
- ^ Стеббинс, Г.Л. (1971). Хромосомная эволюция у высших растений . Лондон: Арнольд. стр. 85–86 . ISBN 9780713122879 .
- ↑ Перейти обратно: Перейти обратно: а б Ромигье Ж, Ру С (2017). «Аналитические предубеждения, связанные с содержанием GC в молекулярной эволюции» . Фронт Генет . 8 : 16. дои : 10.3389/fgene.2017.00016 . ПМК 5309256 . ПМИД 28261263 .
- ↑ Перейти обратно: Перейти обратно: а б Томпсон и Томпсон Генетика в медицине, 7-е изд.
- ↑ Перейти обратно: Перейти обратно: а б Уайт MJD 1973. Цитология и эволюция животных . 3-е изд., Издательство Кембриджского университета.
- ^ К. Лью (2012). Комплексный отбор проб и подготовка проб. Глава: 3.05 – Сбор и обработка проб крови . Академическая пресса. ISBN 9780123813749 .
- ^ Эрвинсия Р., Рианди и Нурджани М. (2017). «Актуальность анализа хромосом человека в сравнении с концепцией мутаций в курсе генетики. Серия конференций IOP» . Материаловедение и инженерия . дои : 10.1088/1757-899x/180/1/012285 . S2CID 90739754 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сакс, Л. (2002). «Насколько распространен интерсекс?» . Журнал сексуальных исследований . 39 (3): 174–178. дои : 10.1080/00224490209552139 . ПМИД 12476264 . S2CID 33795209 .
- ^ Гомеш Си Джей, Харман М.В., Сентуори С.М., Вольгемут К.В., Мартинес Дж.Д. (2018). «Измерение содержания ДНК в живых клетках методом флуоресцентной микроскопии» . Подразделение ячейки . 13 :6. дои : 10.1186/s13008-018-0039-z . ПМК 6123973 . ПМИД 30202427 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Шустер Р.К., Рубинштейн А.Дж., Уоллес округ Колумбия (1988). «Митохондриальная ДНК в безъядерных клетках крови человека» . Биохимия Биофиз Рес Коммьюнити . 155 (3): 1360–5. дои : 10.1016/s0006-291x(88)81291-9 . ПМИД 3178814 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Чжан Д., Кейлти Д., Чжан З.Ф., Чиан Р.К. (2017). «Митохондрии в старении ооцитов: современное понимание» . Факты Мнения Вис Обгын . 9 (1): 29–38. ПМК 5506767 . ПМИД 28721182 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Годфри Л.Р., Masters JC (август 2000 г.). «Теория размножения кинетохор может объяснить быструю эволюцию хромосом» . Учеб. Натл. акад. наук. США . 97 (18): 9821–3. Бибкод : 2000PNAS...97.9821G . дои : 10.1073/pnas.97.18.9821 . ПМК 34032 . ПМИД 10963652 .
- ^ Мейнард Смит Дж. 1998. Эволюционная генетика . 2-е изд., Оксфорд. р218-9
- ^ Годай С., Эстебан М.Р. (март 2001 г.). «Элиминация хромосом у сциарид». Биоэссе . 23 (3): 242–50. doi : 10.1002/1521-1878(200103)23:3<242::AID-BIES1034>3.0.CO;2-P . ПМИД 11223881 . S2CID 43718856 .
- ^ Мюллер Ф., Бернард В., Тоблер Х. (февраль 1996 г.). «Уменьшение хроматина у нематод». Биоэссе . 18 (2): 133–8. дои : 10.1002/bies.950180209 . ПМИД 8851046 . S2CID 24583845 .
- ^ Вингаард Г.А., Грегори Т.Р. (декабрь 2001 г.). «Временный контроль репликации ДНК и адаптивное значение уменьшения хроматина у копепод». Дж. Эксп. Зоол . 291 (4): 310–6. Бибкод : 2001JEZ...291..310W . дои : 10.1002/jez.1131 . ПМИД 11754011 .
- ^ Гилберт С.Ф. 2006. Биология развития . Sinauer Associates, Стэмфорд, Коннектикут. 8-е изд., Глава 9
- ^ Кинг, Стэнсфилд и Маллиган, 2006 г.
- ^ Каррел Л., Уиллард Х (2005). «Профиль Х-инактивации обнаруживает значительную вариабельность экспрессии Х-связанных генов у женщин». Природа . 434 (7031): 400–404. Бибкод : 2005Natur.434..400C . дои : 10.1038/nature03479 . ПМИД 15772666 . S2CID 4358447 .
- ^ Вурстер Д.Х., Бениршке К. (июнь 1970 г.). «Индийский мунтжак, Muntiacus muntjak : олень с низким диплоидным числом хромосом». Наука . 168 (3937): 1364–6. Бибкод : 1970Sci...168.1364W . дои : 10.1126/science.168.3937.1364 . ПМИД 5444269 . S2CID 45371297 .
- ↑ Перейти обратно: Перейти обратно: а б Сюй TC 1979. Цитогенетика человека и млекопитающих: историческая перспектива . Спрингер-Верлаг, Нью-Йорк.
- ^ Кросланд MWJ; Крозье, Р.Х. (1986). « Myrmecia pilosula , муравей только с одной парой хромосом». Наука . 231 (4743): 1278. Бибкод : 1986Sci...231.1278C . дои : 10.1126/science.231.4743.1278 . ПМИД 17839565 . S2CID 25465053 .
- ^ Хандельвал С. (1990). «Эволюция хромосом рода Ophioglossum L». Ботанический журнал Линнеевского общества . 102 (3): 205–217. дои : 10.1111/j.1095-8339.1990.tb01876.x .
- ^ Ким, Д.С.; Нам, Ю.К.; Нет, Джей Кей; Парк, Швейцария; Чепмен, Ф.А. (2005). «Кариотип североамериканского коротконосого осетра Acipenser brevirostrum с наибольшим числом хромосом среди Acipenseriformes». Ихтиологические исследования . 52 (1): 94–97. Бибкод : 2005IchtR..52...94K . дои : 10.1007/s10228-004-0257-z . S2CID 20126376 .
- ^ Мэтти, Р. (15 мая 1945 г.). «Эволюция хромосомной формулы у позвоночных». «Экспериентия» (Базель) . 1 (2): 50–56. дои : 10.1007/BF02153623 . S2CID 38524594 .
- ^ де Оливейра, РР; Фельдберг, Э.; дос Аньос, МБ; Зуанон, Дж. (июль – сентябрь 2007 г.). «Характеристика кариотипа и гетероморфизм половых хромосом ZZ/ZW у двух видов сома рода Ancistrus Kner, 1854 (Siluriformes: Loricariidae) из бассейна Амазонки» . Неотропическая ихтиология . 5 (3): 301–6. дои : 10.1590/S1679-62252007000300010 .
- ^ Пелличиари, К.; Форменти, Д.; Реди, Калифорния; Манфреди, МГ; Романини (февраль 1982 г.). «Изменчивость содержания ДНК у приматов». Журнал эволюции человека . 11 (2): 131–141. дои : 10.1016/S0047-2484(82)80045-6 .
- ^ Соуза, Алжир; О. Корреа, ММ; из Агилара, Коннектикут; Пессоа, LM (февраль 2011 г.). «Новый кариотип Wiedomys pirrhorhinus (Rodentia: Sigmodontinae) из Чапада Диамантина, северо-восток Бразилии» (PDF) . Зоология . 28 (1): 92–96. дои : 10.1590/S1984-46702011000100013 .
- ^ Векслер, М.; Бонвичино, ЧР (3 января 2005 г.). «Таксономия карликовых рисовых крыс рода Oligoryzomys Bangs, 1900 (Rodentia, Sigmodontinae) бразильского Серрадо, с описанием двух новых видов» (PDF) . Arquivos do Museu Nacional, Рио-де-Жанейро . 63 (1): 113–130. ISSN 0365-4508 . Архивировано из оригинала (PDF) 26 марта 2012 года . Проверено 22 апреля 2012 г.
- ^ Стеббинс, Г.Л. (1940). «Значение полиплоидии в эволюции растений». Американский натуралист . 74 (750): 54–66. дои : 10.1086/280872 . S2CID 86709379 .
- ^ Стеббинс 1950
- ^ Комай Л. (ноябрь 2005 г.). «Преимущества и недостатки полиплоида». Нат. Преподобный Жене . 6 (11): 836–46. дои : 10.1038/nrg1711 . ПМИД 16304599 . S2CID 3329282 .
- ^ Адамс К.Л., Вендел Дж.Ф. (апрель 2005 г.). «Полиплоидия и эволюция генома растений». Курс. Мнение. Растительная биол . 8 (2): 135–41. дои : 10.1016/j.pbi.2005.01.001 . ПМИД 15752992 .
- ^ Стеббинс 1971
- ^ Грегори, ТР; Мейбл, БК (2011). «Гл. 8: Полиплоидия у животных» . У Грегори, Т. Райан (ред.). Эволюция генома . Академическая пресса. стр. 427–517. ISBN 978-0-08-047052-8 .
- ^ Уайт, MJD (1973). Хромосомы (6-е изд.). Лондон: Чепмен и Холл. п. 45 .
- ^ Лилли М.А.; Дуронио Р.Дж. (2005). «Новое понимание контроля клеточного цикла эндоцикла дрозофилы » . Онкоген . 24 (17): 2765–75. дои : 10.1038/sj.onc.1208610 . ПМИД 15838513 .
- ^ Эдгар Б.А., Орр-Уивер Т.Л. (май 2001 г.). «Клеточные циклы эндорепликации: больше за меньшие деньги» . Клетка . 105 (3): 297–306. дои : 10.1016/S0092-8674(01)00334-8 . ПМИД 11348589 . S2CID 14368177 .
- ^ Нагл В. 1978. Эндополиплоидия и политения в дифференциации и эволюции: к пониманию количественных и качественных вариаций ядерной ДНК в онтогенезе и филогении . Эльзевир, Нью-Йорк.
- ^ Стеббинс, Г. Ледли-младший, 1972. Хромосомная эволюция высших растений . Нельсон, Лондон. стр. 18
- ^ Идждо Дж.В., Бальдини А., Уорд, округ Колумбия, Ридерс С.Т., Уэллс Р.А. (октябрь 1991 г.). «Происхождение человеческой хромосомы 2: предковое слияние теломер-теломеров» . Учеб. Натл. акад. наук. США . 88 (20): 9051–5. Бибкод : 1991PNAS...88.9051I . дои : 10.1073/pnas.88.20.9051 . ПМК 52649 . ПМИД 1924367 .
- ^ Ригер, Р.; Михаэлис, А.; Грин, ММ (1968). Глоссарий генетики и цитогенетики: Классический и молекулярный . Нью-Йорк: Springer-Verlag. ISBN 9780387076683 .
- ^ Густавссон, Ингемар (3 марта 1969 г.). «Цитогенетика, распространение и фенотипические эффекты транслокации у шведского крупного рогатого скота». Эредитас . 63 (1–2): 68–169. дои : 10.1111/j.1601-5223.1969.tb02259.x . ПМИД 5399228 .
- ^ Сирл, Дж. Б. (1 июня 1984 г.). «Три новые кариотипические расы обыкновенной бурозубки Sorex Araneus (Mammalia: Insectivora) и филогения». Систематическая биология . 33 (2): 184–194. дои : 10.1093/sysbio/33.2.184 . ISSN 1063-5157 .
- ^ Уайт 1973 , с. 169
- ^ Клэг, Д.А.; Далримпл, Великобритания (1987). «Вулканическая цепь Гавайев и Императора, Часть I. Геологическая эволюция» (PDF) . В Декере, RW; Райт, ТЛ; Стауффер, PH (ред.). Вулканизм на Гавайях . Том. 1. С. 5–54. Профессиональный документ Геологической службы США 1350.
- ^ Карсон Х.Л. (июнь 1970 г.). «Хромосомные трассеры происхождения видов». Наука . 168 (3938): 1414–8. Бибкод : 1970Sci...168.1414C . дои : 10.1126/science.168.3938.1414 . ПМИД 5445927 .
- ^ Карсон Х.Л. (март 1983 г.). «Хромосомные последовательности и межостровные колонизации у гавайской дрозофилы » . Генетика . 103 (3): 465–82. дои : 10.1093/генетика/103.3.465 . ПМК 1202034 . ПМИД 17246115 .
- ^ Карсон Х.Л. (1992). «Инверсии у гавайской дрозофилы ». В Кримбасе, КБ; Пауэлл, младший (ред.). Инверсионный полиморфизм дрозофилы . Бока-Ратон, Флорида: CRC Press. стр. 407–439. ISBN 978-0849365478 .
- ^ Канеширо, Кентукки; Гиллеспи, Р.Г.; Карсон, Х.Л. (1995). «Хромосомы и мужские гениталии гавайской дрозофилы: инструменты интерпретации филогении и географии» . В Вагнере, WL; Фанк, Э. (ред.). Гавайская биогеография: эволюция на архипелаге горячей точки . Вашингтон, округ Колумбия: Издательство Смитсоновского института. стр. 57–71 .
- ^ Крэддок Э.М. (2000). «Процессы видообразования в адаптивной радиации гавайских растений и животных». Ин Хехт, Макс К.; Макинтайр, Росс Дж.; Клегг, Майкл Т. (ред.). Эволюционная биология . Том. 31. стр. 1–43. дои : 10.1007/978-1-4615-4185-1_1 . ISBN 978-1-4613-6877-9 .
- ^ Зиглер, Алан К. (2002). Гавайская естественная история, экология и эволюция . Издательство Гавайского университета. ISBN 978-0-8248-2190-6 .
- ^ Малой, Стэнли Р.; Хьюз, Келли (2013). Энциклопедия генетики Бреннера . Сан-Диего, Калифорния: Academic Press. ISBN 978-0-08-096156-9 . OCLC 836404630 .
- ^ Лиза Г. Шаффер; Нильс Томмерап, ред. (2005). ISCN 2005: Международная система цитогенетической номенклатуры человека . Швейцария: С. Каргер АГ. ISBN 978-3-8055-8019-9 .
- ^ Лиер Т., Старке Х., Вайзе А., Лерер Х., Клауссен У. (январь 2004 г.). «Наборы разноцветных зондов FISH и их применение». Гистол. Гистопатол . 19 (1): 229–237. ПМИД 14702191 .
- ^ Шрек Э., Дю Мануар С., Вельдман Т. и др. (июль 1996 г.). «Многоцветное спектральное кариотипирование хромосом человека». Наука . 273 (5274): 494–7. Бибкод : 1996Sci...273..494S . дои : 10.1126/science.273.5274.494 . ПМИД 8662537 . S2CID 22654725 .
- ^ Ван Т.Л., Майерхофер С., Спейчер М.Р. и др. (декабрь 2002 г.). «Цифровое кариотипирование» . Учеб. Натл. акад. наук. США . 99 (25): 16156–61. Бибкод : 2002PNAS...9916156W . дои : 10.1073/pnas.202610899 . ПМЦ 138581 . ПМИД 12461184 .
- ^ Лири, Ребекка Дж; Камминс, Иордания; Ван, Тянь-Ли; Велкулеску, Виктор Э (август 2007 г.). «Цифровое кариотипирование» . Протоколы природы . 2 (8): 1973–1986. дои : 10.1038/nprot.2007.276 . ISSN 1754-2189 . ПМИД 17703209 . S2CID 33337972 .
- ^ Зеленин А.В.; Родионов А.В.; Большева, Нидерланды; Бадаева, ЭД; Муравенко, ОВ (2016). «Геном: происхождение и эволюция термина» . Молекулярная биология . 50 (4): 542–550. дои : 10.1134/S0026893316040178 . ISSN 0026-8933 . ПМИД 27668601 . S2CID 9373640 .
- ^ Вермиш, Йорис Роберт; Раух, Анита (2006). «Ответ Хохстенбаху и др.» . Европейский журнал генетики человека . 14 (10): 1063–1064. дои : 10.1038/sj.ejhg.5201663 . ISSN 1018-4813 . ПМИД 16736034 . S2CID 46363277 .
- ^ Делоне Л.Н. Сравнительное кариологическое исследование видов Muscari Mill. и Беллевалия Лапейр . Вестник Тифлисского ботанического сада. 1922, т. 2, н. 1, с. 1-32 [на русском языке]
- ^ Батталья, Эмилио (1994). «Нуклеосома и нуклеотип: терминологическая критика». Кариология . 47 (3–4): 193–197. дои : 10.1080/00087114.1994.10797297 .
- ^ Дарлингтон CD 1939. Эволюция генетических систем . Издательство Кембриджского университета. 2-е изд., исправленное и дополненное, 1958 г. Oliver & Boyd, Эдинбург.
- ^ МДж, Коттлер (1974). «От 48 до 46: цитологический метод, предвзятое мнение и подсчет хромосом человека». Булл Хист Мед . 48 (4): 465–502. ПМИД 4618149 .
- ^ фон Винивартер Х. (1912). «Исследования сперматогенеза человека». Архивы биологии . 27 (93): 147–9.
- ^ Художник Т.С. (1922). «Сперматогенез человека». Анат. Рез . 23 :129.
- ^ Художник Т.С. (1923). «Исследования сперматогенеза млекопитающих II». Дж. Эксп. Зоология . 37 (3): 291–336. дои : 10.1002/jez.1400370303 .
- ^ Райт, Пирс (11 декабря 2001 г.). «Джо Хин Тджио. Человек, который взломал подсчет хромосом» . Хранитель .
- ^ Тио Дж. Х.; Леван А. (1956). «Число хромосом человека» . Эредитас . 42 (1–2): 1–6. дои : 10.1111/j.1601-5223.1956.tb03010.x . ПМИД 345813 .
- ^ Человеческая хромосома 2 представляет собой слияние двух предков. хромосомы Алека МакЭндрю; по состоянию на 18 мая 2006 г.
- ^ Доказательства общего происхождения: хромосома 2 человека (видео) 2007 г.
Внешние ссылки [ править ]
 СМИ, связанные с кариотипами, на Викискладе?
СМИ, связанные с кариотипами, на Викискладе? - Создание кариотипа — онлайн-занятие Учебного центра генетики Университета Юты.
- Кариотипирование на основе историй болезни в рамках биологического проекта Университета Аризоны.
- Печатный проект кариотипа от Biology Corner, ресурсного сайта для учителей биологии и естественных наук.
- Методы окрашивания и бандажирования хромосом
- Bjorn Biosystems для кариотипирования и FISH. Архивировано 12 июня 2019 г. в Wayback Machine.