Саламандра
| Саламандры | |
|---|---|
 | |
| Пятнистая саламандра , Ambystoma maculatum | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Амфибия |
| Клэйд : | Хвостатый |
| Заказ: | Уродела Дюмериль , 1806 г. |
| Подзаказы | |
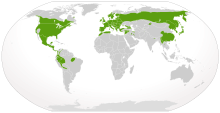 | |
| Местное распространение саламандр (зеленым цветом) | |
Саламандры — это группа амфибий , для которых обычно характерен внешний вид, похожий на ящерицу , со тонким телом, тупой мордой, короткими конечностями, выступающими под прямым углом к телу, и наличием хвоста как у личинок, так и у взрослых особей. Все десять современных семейств саламандр объединены в отряд Urodela из группы Caudata . [2] Уродела — это научный латинский термин, основанный на древнегреческом οὐρά δήλη : Ourà dēlē «заметный хвост». Caudata в переводе с латыни означает «хвостатые», от cauda — «хвост».
Разнообразие саламандр является самым высоким в восточной части Северной Америки , особенно в Аппалачах ; большинство видов обитает в Голарктической области , некоторые виды обитают в Неотропической области . Саламандры никогда не имеют более четырех пальцев на передних ногах и пяти на задних. [3] но у некоторых видов меньше пальцев, а у других отсутствуют задние конечности. Их проницаемая кожа обычно заставляет их полагаться на среду обитания в воде или рядом с ней, а также в других прохладных и влажных местах. Некоторые виды саламандр ведут полностью водный образ жизни на протяжении всей своей жизни, некоторые периодически заходят в воду, а другие во взрослом возрасте полностью ведут наземный образ жизни.
Эта группа земноводных способна регенерировать утраченные конечности, а также другие поврежденные части тела. Исследователи надеются реконструировать регенеративные процессы для потенциальных медицинских применений на людях, таких как лечение травм головного и спинного мозга или предотвращение вредных рубцов во время восстановления после операции на сердце. [4] Замечательная способность саламандр к регенерации не ограничивается только конечностями, но распространяется и на жизненно важные органы, такие как сердце, челюсть и части спинного мозга, что показывает их уникальность по сравнению с другими типами позвоночных. Эта способность наиболее примечательна тем, что возникает без каких-либо рубцов. Это сделало саламандр бесценным модельным организмом в научных исследованиях, направленных на понимание и достижение регенеративных процессов для медицинских достижений в биологии человека и животных. [5]
Представители семейства Salamandridae в основном известны как тритоны , у них отсутствуют реберные бороздки по бокам тела, типичные для других групп. Кожа некоторых видов содержит мощный яд тетродотоксин ; эти саламандры, как правило, медлительны и имеют яркую предупреждающую окраску , демонстрирующую их токсичность. Саламандры обычно откладывают яйца в воде и имеют водных личинок, но в их жизненных циклах происходят большие различия . Некоторые виды в суровых условиях размножаются еще в личиночном состоянии.
Этимология [ править ]
Слово саламандра происходит от старофранцузского salamandre , от латинского salamandra от греческого σαλαμάνδρα : salamándra, неопределенного, возможно, догреческого происхождения. [6] Греческое слово используется для обозначения огненной саламандры . [7]
Описание [ править ]

Кожа лишена чешуи, влажная и гладкая на ощупь, за исключением тритонов Salamandridae, у которых кожа может быть бархатистой или бородавчатой, влажной на ощупь. Кожа может быть тусклой или ярко окрашенной, с различными узорами в виде полос, полос, пятен, пятен или точек. В период размножения самцы тритонов приобретают яркую окраску. Пещерные виды, обитающие в темноте, лишены пигментации и имеют полупрозрачный розовый или перламутровый оттенок. [8]
Размеры саламандр варьируются от мелких саламандр до общей длины От 27 мм ( 1 + 1 ⁄ 8 дюйма), включая хвост, до китайской гигантской саламандры , длина которой достигает 1,8 м (6 футов) и весит до 65 кг (145 фунтов). Все самые крупные виды относятся к четырем семействам: гигантские саламандры , сирены , конголезские угри и протеиды , которые все являются водными и облигатными педоморфами. [9] [10] Некоторые из крупнейших наземных саламандр, прошедших полный метаморфоз, относятся к семейству тихоокеанских гигантских саламандр и значительно мельче. [11] Однако большинство саламандр имеют длину от 10 до 20 см (от 4 до 8 дюймов). [12]
Туловище, конечности и хвост [ править ]
Взрослая саламандра обычно напоминает небольшую ящерицу, имеющую форму тела четвероногих с цилиндрическим туловищем, четырьмя конечностями и длинным хвостом. За исключением семейства Salamandridae, голова, тело и хвост имеют ряд вертикальных впадин на поверхности, которые проходят от средней дорсальной области к брюшной области и известны как реберные бороздки . Их функция, по-видимому, заключается в том, чтобы поддерживать кожу влажной, направляя воду по поверхности тела. [13]

У некоторых водных видов, таких как сирены и амфиумы , задние конечности уменьшены или отсутствуют, что придает им вид угрей , но у большинства видов передние и задние конечности примерно одинаковой длины и выступают в стороны, едва приподнимая туловище над поверхностью тела. земля. Лапы широкие, с короткими пальцами, обычно четыре на передних и пять на задних. У саламандр нет когтей, а форма стопы варьируется в зависимости от среды обитания животного. У альпинистских видов пальцы ног удлиненные, с квадратными кончиками, а у скальных обитателей ступни крупнее с короткими тупыми пальцами. ( Лазающая по деревьям саламандра Bolitoglossa sp .) имеет пластинчатые перепончатые лапы, которые присасываются к гладким поверхностям, тогда как скалолазающие виды Hydromantes из Калифорнии имеют ступни с мясистыми перепонками и короткими пальцами, а хвост используют как дополнительную конечность. При подъеме хвост подпирает заднюю часть тела, в то время как одна задняя нога движется вперед, а затем поворачивается в другую сторону, обеспечивая поддержку при движении другой задней ноги. [14]
У личинок и водных саламандр хвост уплощён с боков, имеет спинные и брюшные плавники и волнообразно колеблется из стороны в сторону, помогая животному двигаться по воде. В семействах Ambystomatidae и Salamandridae хвост самца, который больше, чем у самки, используется во время амплексного объятия , чтобы переместить спаривающуюся пару в укромное место. У наземных видов хвост движется, чтобы уравновесить бегущее животное, тогда как у древесных саламандр и других лазающих по деревьям видов он цепкий . Хвост также используется некоторыми многодонтидными саламандрами , которые умеют прыгать, чтобы подняться в воздух. [14] Хвост используется при ухаживании и как орган хранения белков и липидов. Он также действует как защита от хищников, когда его можно хлестнуть по нападающему или автоматизировать при захвате. В отличие от лягушек, взрослая саламандра способна восстанавливать конечности и хвост, когда они потеряны. [14]
Кожа [ править ]

Кожа саламандр, как и других земноводных, тонкая, проницаемая для воды, служит дыхательной оболочкой, хорошо снабжена железами. Он имеет сильно ороговевшие внешние слои, периодически обновляющиеся в результате процесса отслаивания кожи, контролируемого гормонами гипофиза и щитовидной железы . Во время линьки кожа сначала рвется вокруг рта, и животное продвигается вперед через щель, чтобы сбросить кожу. Когда передние конечности проработаны, серия волн тела подталкивает кожу назад. Задние конечности извлекаются и отталкивают кожу дальше назад, прежде чем она в конечном итоге освобождается в результате трения, когда саламандра движется вперед, прижимая хвост к земле. [15] Затем животное часто поедает образовавшуюся кожуру. [8]
Железы кожи выделяют слизь , которая сохраняет кожу влажной, что является важным фактором кожного дыхания и терморегуляции. Липкий слой защищает от бактериальных инфекций и плесени, уменьшает трение при плавании, делает животное скользким и затрудняет его поимку хищниками. Зернистые железы, разбросанные по верхней поверхности, особенно на голове, спине и хвосте, производят репеллентные или токсичные выделения. [15] Некоторые токсины саламандры особенно сильны. Грубокожий тритон ( Taricha granulosa ) вырабатывает нейротоксин тетродотоксин , самое токсичное из известных небелковых веществ. Обращение с тритонами не причиняет вреда, но проглатывание даже малейшего кусочка кожи смертельно опасно. В ходе испытаний с кормлением было обнаружено, что рыбы, лягушки, рептилии, птицы и млекопитающие восприимчивы. [16]
Взрослые взрослые особи некоторых видов саламандр имеют «брачную» железистую ткань в клоаках , у основания хвоста, на голове или под подбородком. Некоторые самки выделяют химические вещества , возможно, из вентральной клоакальной железы, чтобы привлечь самцов, но самцы, похоже, не используют феромоны для этой цели. [17] У некоторых многодонтов самцы имеют заметные подбородочные железы на подбородке, которые прижимаются к ноздрям самок во время ритуала ухаживания. Они могут ускорять процесс спаривания, снижая риск его прерывания хищником или конкурирующим самцом. [18] Железа у основания хвоста Plethodon cinereus используется для маркировки фекальных шариков, чтобы провозгласить территориальную принадлежность. [17]
Чувства [ править ]


Запах [ править ]
Обоняние саламандр играет роль в поддержании территории, распознавании хищников и ритуалах ухаживания, но, вероятно, вторично по отношению к зрению во время выбора добычи и кормления. У саламандр есть два типа сенсорных областей, которые реагируют на химию окружающей среды. Обонятельный эпителий в носовой полости улавливает запахи воздуха и воды, а прилегающие сошниково-носовые органы улавливают нелетучие химические сигналы, такие как вкусы во рту. У полнодонтов чувствующий эпителий вомероназальных органов доходит до носогубных борозд , которые тянутся от ноздрей к углам рта. Эти расширенные области, по-видимому, связаны с идентификацией объектов добычи, распознаванием сородичей и идентификацией особей. [19]
Видение [ править ]
Глаза большинства саламандр приспособлены прежде всего для видения в ночное время. У некоторых постоянно водных видов они уменьшены в размерах и имеют упрощенное строение сетчатки , а у пещерных обитателей, таких как слепая саламандра Джорджии , они отсутствуют или покрыты слоем кожи. У земноводных глаза являются компромиссом: они близоруки в воздухе , а в воде — дальнозорки . Полностью наземные виды, такие как огненная саламандра, имеют более плоскую линзу, которая может фокусироваться на гораздо более широком диапазоне расстояний. [20] Чтобы найти свою добычу, саламандры используют трехцветное цветовое зрение , простирающееся до ультрафиолетового диапазона, основанное на трех типах фоторецепторов , которые максимально чувствительны к длинам волн около 450, 500 и 570 нм. [21] Личинки и взрослые особи некоторых высоководных видов также имеют орган боковой линии , аналогичный органу рыб, который может обнаруживать изменения давления воды. [8]
Слушание [ править ]
У всех саламандр нет полости среднего уха, барабанной перепонки и евстахиевой трубы , но они имеют покрышку, как у лягушек, и все еще способны улавливать звуки, передающиеся по воздуху. [22] [23] Система opercularis состоит из двух косточек: колумеллы (эквивалента стремечка высших позвоночных ) , сросшейся с черепом, и жаберной крышки. Покрышечная мышца соединяет последнюю с грудным поясом и находится в напряжении, когда животное насторожено. [24] Кажется, система способна обнаруживать низкочастотные вибрации (500–600 Гц), которые могут улавливаться от земли передними конечностями и передаваться во внутреннее ухо. Они могут служить предупреждением животного о приближающемся хищнике. [25]
Вокализация [ править ]
Обычно считается, что саламандры не имеют голоса и не используют звук для общения, как это делают лягушки; однако в системе спаривания они общаются посредством передачи сигналов феромонов; некоторые виды могут издавать тихое тиканье. щелчки, скрипы или хлопающие звуки, [26] возможно, путем открытия и закрытия клапанов в носу. У большинства саламандр отсутствуют голосовые связки, но у илистого щенка (Necturus) и некоторых других видов имеется гортань, а у тихоокеанских гигантских саламандр и некоторых других есть большая гортань и полосы, известные как голосовые складки. [27] Калифорнийская гигантская саламандра может издавать лай или хрипеть, а некоторые виды могут пищать, сокращая мышцы горла. Древесная саламандра может пищать, используя другой механизм; он втягивает глаза в голову, вытесняя воздух изо рта. Саламандра энсатина иногда издает шипящий звук, а сирены иногда издают тихие щелчки и в случае нападения могут издавать слабые вопли. Подобное щелкающее поведение наблюдалось у двух европейских тритонов Lissotriton vulgaris и Ichthyosaura alpestris в водной фазе. [26] Вокализация саламандр мало изучена, и предполагается, что цель этих звуков — отпугнуть хищников. [28]

Дыхание [ править ]
Дыхание различается у разных видов саламандр и может затрагивать жабры, легкие, кожу и оболочки рта и горла. Личинки саламандр дышат преимущественно с помощью жабр , которые обычно внешние и перистые. Вода всасывается через рот и вытекает через жаберные щели. Некоторые неотенические виды, такие как илистый щенок ( Necturus maculosus ), сохраняют жабры на протяжении всей жизни, но большинство видов теряют их при метаморфозе . Эмбрионы , подвергающихся прямому развитию , некоторых наземных безлегочных саламандр, таких как Ensatina имеют большие жабры, лежащие близко к поверхности яйца. [29]
Легкие, присутствующие у взрослых саламандр, сильно различаются у разных видов по размеру и строению. У водных, холодноводных видов, таких как речные саламандры ( Ryacotriton ), легкие очень маленькие с гладкими стенками, в то время как виды, живущие в теплой воде с небольшим количеством растворенного кислорода, такие как малая сирена ( Siren intermedia ), имеют большие легкие с извилистыми стенками. поверхности. У безлёгочных саламандр ( семейство Plethodontidae и когтистые саламандры семейства азиатских саламандр ) отсутствуют лёгкие и жабры, и газообмен в основном происходит через кожу, так называемое кожное дыхание , дополняемое тканями, выстилающими рот. Для облегчения этого у этих саламандр имеется густая сеть кровеносных сосудов под кожей и во рту. [29] [30] [31]
У амфиумов метаморфоз неполный, и во взрослом возрасте они сохраняют одну пару жаберных щелей с полностью функционирующими внутренними легкими. [32] Некоторые виды, у которых отсутствуют легкие, дышат через жабры. В большинстве случаев это внешние жабры, видимые в виде пучков по обе стороны головы. У некоторых наземных саламандр для дыхания используются легкие, хотя они простые и похожи на мешочки, в отличие от более сложных органов млекопитающих . У многих видов, таких как олм , во взрослом состоянии есть и легкие, и жабры. [8]

У Necturus наружные жабры начинают формироваться как средство борьбы с гипоксией яйца, поскольку яичный желток превращается в метаболически активную ткань. [33] Однако молекулярные изменения у щенков во время постэмбрионального развития, в первую очередь за счет щитовидной железы, предотвращают интернализацию наружных жабр, как это наблюдается у большинства саламандр, претерпевающих метаморфоз. [34] Внешние жабры саламандр сильно отличаются от жабр земноводных с внутренними жабрами. В отличие от земноводных с внутренними жабрами, которые обычно полагаются на изменение давления в щечной и глоточной полостях, чтобы обеспечить диффузию кислорода на жаберную занавеску, неотенические саламандры, такие как Necturus, используют специальную мускулатуру, такую как levatores arcuum, для перемещения внешних жабр и поддержания дыхательные поверхности постоянно контактируют с новой насыщенной кислородом водой. [35] [36]
Кормление и диета [ править ]
Саламандры – оппортунистические хищники . Обычно они не ограничиваются определенной пищей, а питаются практически любым организмом разумного размера. [37] Крупные виды, такие как японская гигантская саламандра ( Andrias japonicus ), питаются крабами, рыбой, мелкими млекопитающими, амфибиями и водными насекомыми. [38] В ходе исследования более мелких темных саламандр ( Desmognathus ) в Аппалачах их рацион включает дождевых червей , мух , жуков , личинок жуков цикадок , ногохвостов , мотыльков , пауков , , кузнечиков и клещей . [37] Иногда имеет место каннибализм , особенно когда ресурсы ограничены или время ограничено. Головастики тигровой саламандры в эфемерных лужах иногда прибегают к поеданию друг друга и, по-видимому, способны нападать на неродственных особей. [39] Взрослые чернобрюхие саламандры ( Desmognathus Quadramulatus ) охотятся на взрослых особей и детенышей других видов саламандр, а их личинки иногда поедают более мелких личинок. [40]

У большинства видов саламандр маленькие зубы как на верхней, так и на нижней челюсти. В отличие от лягушек , такими зубами обладают даже личинки саламандр. [8] Хотя зубы личинок имеют форму заостренных конусов, зубы взрослых особей приспособлены для быстрого захвата добычи. Коронка коллагеновыми , имеющая два бугорка (двустворчатая), прикреплена к ножке волокнами . Сустав, образующийся между премоляром и ножкой, является частично гибким, так как может изгибаться внутрь, но не наружу. Когда борющаяся добыча продвигается в рот саламандры, кончики зубов расслабляются и изгибаются в одном направлении, стимулируя движение к горлу и препятствуя побегу добычи. [41] У многих саламандр есть участки зубов, прикрепленные к сошнику и небным костям нёба, которые помогают удерживать добычу. Все типы зубов резорбируются и периодически заменяются на протяжении всей жизни животного. [42]
Наземная саламандра ловит свою жертву, высовывая липкий язык , что занимает менее полсекунды. У некоторых видов язык прикрепляется спереди ко дну рта, а у других - на ножке. Он становится липким из-за выделений слизи желез на его кончике и нёбе. [43] Высокоскоростная кинематография показывает, как тигровая саламандра ( Ambystoma tigrinum ) располагается мордой близко к добыче. Затем его рот широко раскрывается, нижняя челюсть остается неподвижной, а язык выпячивается и меняет форму, выбрасываясь вперед. Высунутый язык имеет углубление в центре, и его край загибается внутрь при ударе по цели, запирая добычу в наполненной слизью корыте. Здесь его удерживают, пока шея животного согнута, язык втянут и челюсти сомкнуты. Крупную или устойчивую добычу удерживают зубы, в то время как повторяющиеся высовывания и втягивания языка втягивают ее. Глотание включает попеременное сокращение и расслабление мышц горла, чему способствует нажатие глазных яблок в нёбо. [44] Многие безлегочные саламандры семейства Plethodontidae имеют более сложные методы питания. Мышцы, окружающие подъязычную кость, сокращаются, сохраняя упругую энергию в упругой соединительной ткани, и фактически «выстреливают» подъязычную кость изо рта, удлиняя таким образом язык. [45] [46] Мышцы, которые берут начало в области таза и прикрепляются к языку, используются для возвращения языка и подъязычной кости в исходное положение. [47]
У водной саламандры отсутствуют мышцы языка, и она ловит добычу совершенно другим способом. Он хватает пищу, сжимает ее зубами и применяет своего рода инерционное кормление. Это включает в себя встряхивание головой, резкое втягивание и вытягивание воды изо рта и щелканье челюстями, и все это имеет тенденцию разрывать и мацерировать добычу, которую затем проглатывают. [47]
часто питаются медлительными животными, такими как улитки , креветки и черви , Хотя сирены они уникальны среди саламандр тем, что у них развились растительноядные виды, такие как клювообразные концы челюстей и обширный кишечник. В дикой природе они питаются водорослями и другими мягкими растениями и легко поедают предложенный им салат . [48]
Защита [ править ]
У саламандр тонкая кожа и мягкое тело, они двигаются довольно медленно и на первый взгляд могут показаться уязвимыми для оппортунистических хищников. Однако у них есть несколько эффективных линий защиты. Слизистый налет на влажной коже затрудняет захват, а слизистый налет может иметь неприятный вкус или быть токсичным. При нападении хищника саламандра может расположиться так, чтобы основные ядовитые железы были обращены к агрессору. Часто они находятся на хвосте, который может вилять или загибаться вверх и выгибаться над спиной животного. Жертва хвоста может оказаться стоящей стратегией, если саламандра уцелеет и хищник научится избегать этого вида саламандр в будущем. [49]
Апосематизм [ править ]

кожные выделения тигровой саламандры ( Ambystoma tigrinum ), скармливаемые крысам, вызывают отвращение к вкусу, и крысы избегали презентационной среды, когда ее предлагали им снова. Было показано, что [50] Огненная саламандра ( Salamandra salamandra ) имеет гребень крупных зернистых желез вдоль позвоночника, которые способны выплеснуть тонкую струю токсичной жидкости в нападающего. Правильно наклонив корпус, он может точно направить струю на расстояние до 80 см (30 дюймов). [51]
У иберийского ребристого тритона ( Pleurodeles waltl ) есть еще один метод отпугивания агрессоров. Его кожа выделяет ядовитую вязкую жидкость, при этом тритон поворачивает острые ребра на угол от 27 до 92° и принимает надутую позу. В результате этого действия ребра прокалывают стенку тела, при этом каждое ребро выступает через оранжевую бородавку, расположенную в боковой ряд. Это может служить апосематическим сигналом, который делает шипы более заметными. Когда опасность миновала, ребра втягиваются и кожа заживает. [52]
Камуфляж и мимикрия [ править ]
Хотя многие саламандры имеют загадочную окраску, чтобы быть незаметными, другие сигнализируют о своей токсичности своей яркой окраской . Обычно используются желтый, оранжевый и красный цвета, часто с черным для большей контрастности. Иногда в случае нападения животное принимает позу, демонстрируя вспышку предупреждающего оттенка на нижней стороне. Красный эфт, ярко окрашенная наземная молодая форма восточного тритона ( Notophthalmus viridescens ), очень ядовит. Птицы и змеи избегают его, и он может выжить до 30 минут после проглатывания (позже срыгивания). [53] Красная саламандра ( Pseudotriton Ruber ) — приятный на вкус вид, окраска которого похожа на красную эфту. Было показано, что хищники, которые ранее питались им, избегают его после встречи с красными эфтами, что является примером бейтсовской мимикрии . [53] Другие виды демонстрируют аналогичную мимикрию. В Калифорнии вкусная желтоглазая саламандра ( Ensatina eschscholtzii ) очень напоминает ядовитого калифорнийского тритона ( Taricha torosa ) и грубокожего тритона ( Tricha granulosa ), тогда как в других частях ареала она имеет загадочную окраску. [54] Существует корреляция между токсичностью калифорнийских видов саламандр и дневным образом жизни: относительно безобидные виды, такие как калифорнийская стройная саламандра ( Batrachoseps attenuatus ), ведут ночной образ жизни и поедаются змеями, тогда как калифорнийский тритон имеет в коже множество крупных ядовитых желез, ведет дневной, и его избегают змеи. [55]
Автотомия [ править ]
Некоторые виды саламандр используют автотомию хвоста, чтобы спастись от хищников. Хвост после нападения отпадает и некоторое время извивается, а саламандра либо убегает, либо остается достаточно неподвижной, чтобы ее не заметили, пока хищник отвлекается. Хвост со временем отрастает, и саламандры регулярно регенерируют другие сложные ткани, включая хрусталик или сетчатку глаза. Всего за несколько недель после потери части конечности саламандра прекрасно восстанавливает недостающую структуру. [56]
Распространение и среда обитания [ править ]
Саламандры отделились от других амфибий в середине и конце перми и первоначально были похожи на современных представителей Cryptobranchoidea . Их сходство с ящерицами является результатом симплезиоморфии , общего сохранения примитивного строения тела четвероногих, но они не более тесно связаны с ящерицами, чем с млекопитающими. Их ближайшие родственники — лягушки и жабы, обитающие в Батрахии .
Самая старая известная общей группы ( Caudata саламандра ) — Triassurus из триаса Кыргызстана . [57] саламандр известны из средней юры Англии . Другие окаменелости [58] Шотландия , [59] Китай , [60] и Казахстан . [61] Самый старый известный саламандра кроновой группы ( Urodela но недавние анализы показывают, что это Вальдотритон из поздней юры Испании ) остается неопределенным , . [59]
Саламандры встречаются только в Голарктической и Неотропической областях, не достигая юга Средиземноморского бассейна , Гималаев или в Южной Америке бассейна Амазонки . Они не простираются к северу от границы арктических деревьев , а самый северный азиатский вид Salamandrellakeyserlingii может пережить длительное замораживание при температуре -55 °C. [62] Встречается в сибирских лиственничных лесах Саха и самый северный вид Северной Америки — Ambystoma Laterale , не доходящий на север дальше лабрадора и Taricha granulosa и не дальше Аляскинского Панхандла . [63] Они имели исключительно лавразийское распространение, пока Bolitoglossa не вторглись в Южную Америку из Центральной Америки, вероятно, к началу раннего миоцена , около 23 миллионов лет назад. [64] Они также жили на Карибских островах в эпоху раннего миоцена , что подтверждается открытием Palaeoplethodon hispaniolae , [65] найден в ловушке в янтаре в Доминиканской Республике . Окаменелости позвонков, обнаруженные на окаменелостях Мургона, предположительно принадлежат саламандре. [66] хотя его истинная идентичность оспаривается. Если бы позвонки действительно принадлежали саламандрам, они были бы единственными саламандрами в Австралии .
Существует около 760 ныне живущих видов саламандр. [67] [68] Треть известных видов саламандр обитает в Северной Америке. Самая высокая их концентрация наблюдается в районе Аппалачей, где, как полагают, Plethodontidae произошли от горных ручьев. Здесь растительные зоны и близость к воде имеют большее значение, чем высота. Лишь виды, принявшие более наземный образ жизни, смогли расселиться в другие местности. Северная склизкая саламандра ( Plethodon Glutinosus ) имеет широкий ареал и занимает ареал, сходный с ареалом южной серощекой саламандры ( Plethodon metcalfi ). Последний ограничен немного более прохладными и влажными условиями в лесах бухт , обращенных на север , в южных Аппалачах, а также на высотах выше 900 м (3000 футов), в то время как первый более адаптируем и вполне способен обитать в этих местах. , но какой-то неизвестный фактор, похоже, мешает этим двум видам сосуществовать. [37]
Один из видов, саламандра Андерсона , является одним из немногих видов живых земноводных, обитающих в солоноватой или соленой воде. [69]
и развитие Размножение

Многие саламандры не используют вокализацию. [70] и у большинства видов полы похожи, поэтому они используют обонятельные и тактильные сигналы для идентификации потенциальных партнеров, и половой отбор происходит . Феромоны играют важную роль в этом процессе и могут вырабатываться брюшными железами у мужчин и клоакальными железами и кожей у обоих полов. Иногда можно увидеть самцов, исследующих потенциальных партнеров мордой. У тритонов Старого Света Triturus spp. самцы половодиморфны и располагаются впереди самок. Считается, что визуальные сигналы также важны для некоторых видов плетодонтов . [71]
За исключением наземных видов трех семейств Plethodontidae , Ambystomatidae и Salamandridae , саламандры спариваются в воде. [72] Спаривание варьируется от ухаживания между одиночными самцом и самкой до взрывного группового размножения. [73] У клады Salamandroidea , составляющей около 90% всех видов, оплодотворение внутреннее. [74] Как правило, саламандры с внутренним оплодотворением имеют непрямую передачу спермы, но у таких видов, как сардинская ручьевая саламандра , корсиканская ручьевая саламандра , кавказская саламандра и пиренейская ручьевая саламандра , самец передает свою сперму непосредственно в клоаку самки. [75] [76] [77] У видов с непрямой передачей спермы самец откладывает сперматофор на землю или в воду в зависимости от вида, а самка подбирает его своим выходным отверстием. Сперматофор имеет пакет со спермой, поддерживаемый на коническом студенистом основании, и часто его отложение и сбор включают в себя тщательно продуманное ухаживание. Попав внутрь клоаки, сперматозоиды перемещаются в сперматеку , одну или несколько камер в крыше клоаки, где они хранятся иногда в течение длительного периода времени, пока яйцеклетка не будет отложена. У азиатских саламандр , гигантских саламандр и сиренид , являющихся наиболее примитивными группами, оплодотворение наружное. В ходе репродуктивного процесса, аналогичного процессу типичных лягушек, самец выделяет сперму в яйцеклетку. У этих саламандр также есть самцы, проявляющие родительскую заботу , которая в противном случае возникает только у самок при внутреннем оплодотворении. [71] [78]
Встречаются три различных типа откладывания яиц. Ambystoma и Taricha spp. откладывают большое количество мелких икринок в тихих водоемах, где маловероятно появление крупных хищников. Большинство темных саламандр ( Desmognathus ) и тихоокеанских гигантских саламандр ( Dicamptodon ) откладывают небольшие порции яиц среднего размера в скрытом месте в проточной воде, и их обычно охраняет взрослая особь, обычно самка. Многие тропические вьющиеся саламандры ( Bolitoglossa ) и безлегочные саламандры (Plethodontinae) откладывают небольшое количество крупных яиц на суше в хорошо укромном месте, где их также охраняет мать. [71] Некоторые виды, такие как огненные саламандры ( Salamandra ), являются яйцеживородящими , при этом самка сохраняет яйца внутри своего тела до тех пор, пока они не вылупятся либо в личинки, которые будут отложены в водоеме, либо в полностью сформировавшуюся молодь. [8]
В регионах с умеренным климатом размножение обычно носит сезонный характер, и саламандры могут мигрировать в места размножения. Самцы обычно прибывают первыми и в некоторых случаях осваивают территории . Обычно следует личиночная стадия, на которой организм полностью водный. У головастика три пары наружных жабр, нет век, длинное тело, уплощенный с боков хвост со спинными и брюшными плавниками, а у некоторых видов конечности-зачатки или конечности. Личинки прудового типа могут иметь пару стержнеобразных балансиров по обе стороны головы, длинные жаберные нити и широкие плавники. Личинки ручьевого типа более стройные, с короткими жаберными нитями — у Rhyacotriton и Onychodactylus, а у некоторых видов у Batrachuperus жабры и жаберные тычинки чрезвычайно редуцированы, [79] более узкие плавники и отсутствие балансиров, вместо этого задние конечности уже развиты, когда они вылупляются. [80] Головастики плотоядны , и личиночная стадия может длиться от нескольких дней до лет, в зависимости от вида. Иногда эту стадию полностью пропускают, и яйца большинства безлегочных саламандр (Plethodontidae) развиваются непосредственно в миниатюрные версии взрослых особей без промежуточной личиночной стадии. [81]
К концу личиночной стадии у головастиков уже есть конечности и метаморфоз протекает нормально. У саламандр это происходит в течение короткого периода времени и включает закрытие жаберных щелей и потерю таких структур, как жабры и хвостовые плавники, которые не требуются во взрослом возрасте. При этом развиваются веки, рот становится шире, появляется язык, формируются зубы. Водная личинка выходит на сушу уже взрослой наземной особью. [82]

Не все виды саламандр идут по этому пути. Неотения , также известная как педоморфоз, наблюдалась во всех семействах саламандр и может быть повсеместно возможна у всех видов саламандр. В этом состоянии особь может сохранять жабры или другие ювенильные черты при достижении репродуктивной зрелости. Изменения, происходящие при метаморфозе, находятся под контролем гормонов щитовидной железы , а у облигатных неотенов, таких как аксолотль ( Ambystoma mexicanum ), ткани, по-видимому, не реагируют на гормоны. У других видов изменения могут не запускаться из-за недостаточной активности гипоталамо-гипофизарно-щитовидного механизма, что может возникнуть, когда условия наземной среды слишком негостеприимны. [82] Это может быть связано с холодом или резкими колебаниями температуры, засушливостью, недостатком еды, отсутствием укрытия или недостатком йода для образования гормонов щитовидной железы. Генетика также может сыграть свою роль. у личинок тигровых саламандр ( Ambystoma tigrinum Например, ) конечности развиваются вскоре после вылупления, и в сезонных водоемах они быстро подвергаются метаморфозу. Другие личинки, особенно в постоянных водоемах и в более теплом климате, могут не претерпевать метаморфоза до тех пор, пока не станут взрослыми. Другие популяции в более холодном климате могут вообще не метаморфизоваться и стать половозрелыми, находясь в личиночной форме. Неотения позволяет этому виду выжить, даже если земная среда слишком сурова для того, чтобы взрослые особи могли жить на суше. [80]
Сохранение [ править ]

Общее сокращение численности ныне живущих видов земноводных связано с грибковым заболеванием хитридиомикозом . Более высокая доля видов саламандр, чем лягушек или червяг, находится в одной из категорий риска, установленных МСОП . За последние несколько десятилетий 20-го века численность саламандр значительно сократилась, хотя прямой связи между грибком и сокращением популяции пока не обнаружено. [83] В 2005 году МСОП предпринял дальнейшие усилия, учредив План действий по сохранению амфибий (ACAP), за которым впоследствии последовали Amphibian Ark (AArk), Группа специалистов по амфибиям (ASG) и, наконец, головная организация, известная как Альянс по выживанию амфибий (ASA). ). [84] Исследователи также называют вырубку лесов , приводящую к фрагментации подходящей среды обитания, и изменение климата в качестве возможных факторов, способствующих этому. Такие виды, как Pseudoeurycea brunnata и Pseudoeurycea goebeli , которые в изобилии встречались в облачных лесах Гватемалы и Мексики в 1970-е годы, к 2009 году оказались редкими. [85] Однако за прошедшие годы было собрано мало данных о размерах популяций, и благодаря интенсивному обследованию исторических и подходящих новых мест удалось обнаружить особей других видов, таких как Parvimolge Townsendi , которые считались вымершими . [83] В настоящее время основные меры по сохранению саламандр включают методы сохранения как in situ , так и ex situ . Предпринимаются усилия по сохранению некоторых представителей семейства саламандр в рамках программы природоохранного разведения (CBP), но важно отметить, что необходимо заранее провести исследование, чтобы определить, действительно ли вид саламандр будет иметь ценность от CBP, поскольку исследователи отмечают, что некоторые виды амфибий полностью не справляются с этой средой. [84]
Во всем мире предпринимаются различные инициативы по сохранению. Китайская гигантская саламандра , самая крупная амфибия в мире высотой 1,8 м (6 футов), находится под угрозой исчезновения , поскольку ее собирают для употребления в пищу и для использования в традиционной китайской медицине . Осуществляется программа экологического образования, направленная на поощрение устойчивого управления дикими популяциями в горах Циньлин , а также созданы программы разведения в неволе. [86] Повелитель ада — еще один крупный и долгоживущий вид, численность которого сокращается и меньше молодых особей, достигающих зрелости, чем раньше. [87] Еще одним тревожным открытием является рост аномалий у 90% популяции магов ада в водоразделе реки Спринг в Арканзасе. [88] Утрата среды обитания, заиливание рек, загрязнение окружающей среды и болезни — все это привело к упадку численности, и в зоопарке Сент-Луиса была успешно реализована программа разведения в неволе. [89] Считается , что из 20 видов мельчайших саламандр ( Thorius spp.) в Мексике половина вымерла, а большинство остальных находятся под угрозой исчезновения. Конкретными причинами сокращения могут быть изменение климата, хитридиомикоз или вулканическая активность, но главной угрозой является разрушение среды обитания , поскольку лесозаготовки, сельскохозяйственная деятельность и населенные пункты сокращают их зачастую крошечные и фрагментированные ареалы. В настоящее время проводится исследовательская работа для оценки состояния этих саламандр и лучшего понимания факторов, связанных с сокращением их популяции, с целью принятия мер. [90]
Ambystoma mexicanum , водная саламандра, является видом, охраняемым Мексиканским UMA (Отделом по управлению и сохранению дикой природы) с апреля 1994 года. Еще одним пагубным фактором является то, что аксолотль потерял свою роль главного хищника с момента появления местных экзотических видов. такие как нильская тилапия и карп. Тилапия и карп напрямую конкурируют с аксолотлями, поедая их икру, личинки и молодь. Изменение климата также сильно повлияло на аксолотлей и их популяцию на юге Мексики. Из-за близости к Мехико чиновники в настоящее время работают над программами на озере Сочимилько, направленными на развитие туризма и обучение местного населения восстановлению естественной среды обитания этих существ. [91] Эта близость является важным фактором, повлиявшим на выживание аксолотлей, поскольку город расширился и захватил регион Сочимилько, чтобы использовать его ресурсы для воды, продовольствия и канализации. [92] Тем не менее, аксолотль имеет то преимущество, что его выращивают на фермах для исследовательских целей. Так что шанс, что они смогут вернуться в свою естественную среду обитания, все еще есть. Недавнее сокращение популяции существенно повлияло на генетическое разнообразие популяций аксолотлей, что затруднило дальнейший научный прогресс. Важно отметить, что, хотя существует определенный уровень генетического разнообразия из-за того, что популяции амбистом , таких как аксолотль, являются педеоморфными видами, это не объясняет общего отсутствия разнообразия. Есть свидетельства, указывающие на историческое узкое место амбистомы , которое усугубляет проблемы вариаций. К сожалению, в отличие от исторических времен, у этого вида нет большого генетического фонда, из которого можно было бы извлечь пользу. Таким образом, существует серьезная обеспокоенность по поводу инбридинга из-за отсутствия потока генов. [93] Одним из способов сохранения генетического разнообразия в популяции является криоконсервация сперматофоров самца аксолотля. Это безопасный и неинвазивный метод, который требует сбора сперматофоров и помещения их в глубокую заморозку для сохранения. Самое главное, они обнаружили, что при оттаивании сперматофорам наносится лишь ограниченный ущерб, и поэтому этот вариант является жизнеспособным. По состоянию на 2013 год этот метод используется для спасения не только аксолотля, но и многих других представителей семейства саламандр. [92] [94] [95]
В настоящее время проводятся исследования экологических сигналов, которые необходимо воспроизвести, прежде чем можно будет убедить содержащихся в неволе животных размножаться. Распространенным видам, таким как тигровая саламандра и грязевой щенок, вводят гормоны для стимуляции выработки спермы и яйцеклеток, а также роль аргинин -вазотоцина исследуется в брачном поведении. Другое направление исследований — искусственное оплодотворение , либо in vitro , либо путем введения сперматофоров в клоаки самок. Результаты этого исследования могут быть использованы в программах по разведению в неволе исчезающих видов. [96]
Таксономия [ править ]
Название отряда Urodela происходит от имени Urodeles, данного Андре Мари Констан Дюмерилем в 1805 году. [2] оно происходит от греческих слов οὐρά Ourā «хвост» и δῆλος delos «видимый, заметный» из-за их «стойких» хвостов. [97]
Среди различных авторитетов существуют разногласия относительно определения терминов Caudata и Urodela. Некоторые утверждают, что Urodela следует ограничить кроновой группой , а Caudata использовать для всей группы. [2] [ нужна ссылка ] Другие ограничивают название Caudata кроновой группой и используют Urodela для всей группы. [98] [99] Первый подход, по-видимому, получил наиболее широкое распространение и используется в этой статье. [68]
Десять семейств, принадлежащих Urodela, разделены на три подотряда. [98] Кладу Neocaudata часто используют для отделения Cryptobranchoidea и Salamandroidea от Sirenoidea.
| Cryptobranchoidea (Гигантские саламандры) | |||
| Семья | Общие имена | Пример вида | Пример изображения |
|---|---|---|---|
| Криптобранхиды | Гигантские саламандры | Повелитель ада ( Cryptobranchus alleganiensis ) | |
| Гинобииды | Азиатские саламандры | Саламандра Хида ( Hynobius kimurae ) |  |
| Salamandroidea (Продвинутые саламандры) | |||
| Амбистоматиды | Кротовые саламандры | Саламандра мраморная ( Ambystoma opacum ) |  |
| Амфиумиды | Амфиумы или конголезские угри | Двупалая амфиума ( Амфиума означает ) |  |
| Плетодонтиды | Безлегочные саламандры | Красноспинная саламандра ( Plethodon cinereus ) |  |
| Протеиды | Mudpuppies и olms | Ольм ( Proteus anguinus ) | |
| Риакотритониды | Торрент-саламандры | Южная речная саламандра ( Rhyacotriton variegatus ) |  |
| Саламандриды | Тритоны и настоящие саламандры | Альпийский тритон ( Ichthyosaura alpestris ) | |
| Сиреноидея (Сирены) | |||
| Сирениды | Сирены | Сирена большая ( Sirenlacertina ) |
и эволюция Филогения
Происхождение и эволюционные взаимоотношения между тремя основными группами амфибий (гимнофионами, хвостатыми и бесхвостыми) являются предметом дискуссий. Молекулярная филогения 2005 года, основанная на анализе рДНК , показала, что первое расхождение между этими тремя группами произошло вскоре после того, как они отделились от лопастных рыб в девоне (около 360 миллионов лет назад) и до распада суперконтинента. Пангея . Кратковременность этого периода и скорость распространения радиации могут помочь объяснить относительную нехватку окаменелостей амфибий, которые, по-видимому, тесно связаны с лиссамфибиями . [100] Однако более поздние исследования, как правило, обнаружили, что более поздние (поздний карбон) [101] в Пермь [102] ) возраст наибольшей дивергенции среди лисамфибий.

Самым ранним известным лиссамфибией линии саламандры является Triassurus из среднего-позднего триаса Кыргызстана. [103] Другие ископаемые саламандры известны из средней-поздней юры Евразии, в том числе Kokartus Honorarius из средней юры Кыргызстана, два вида явно неотенического водного Marmorerpeton из средней юры Англии и Шотландии, [104] и Karaurus из средней-поздней юры Казахстана по морфологии напоминали современных кротовых саламандр и, вероятно, вели сходный роющий образ жизни. [68] Они выглядели как крепкие современные саламандры, но не имели ряда анатомических особенностей, характерных для всех современных саламандр. [105]
Две группы современных саламандр — это Cryptobranchoidea (в которую входят азиатские и гигантские саламандры) и Salamandroidea (в которую входят все другие ныне живущие саламандры), также известные как Diadectosalamandroidei. Обе группы известны из средней-поздней юры Китая. примером первого являются Chunerpeton tianyiensis , Pangerpeton sinensis , Jeholotriton paradoxus , Regalerpeton weichangensis , Liaoxitriton daohugouensis и Iridotriton hechti , а второго — Beiyanerpeton jianpingensis . К верхнему мелу , вероятно, появилось большинство или все ныне живущие семейства саламандр. [68]
Следующая кладограмма показывает взаимоотношения между семействами саламандр на основе молекулярного анализа Пайрона и Винса (2011). [106] Положение Sirenidae оспаривается, но положение сестры Salamandroidea лучше всего соответствует молекулярным и ископаемым данным. [68]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Геном и генетика [ править ]
Саламандры обладают гигантскими геномами размером от 14 до 120 ГБ. [107] ( длина генома человека составляет 3,2 Гб). геномы Pleurodeles waltl (20 Гб) и Ambystoma mexicanum (32 Гб). Секвенированы [108] [109]
Их гигантские геномы сильно повлияли на их физиологию. Это касается их скелета и системы кровообращения и привело к упрощению работы мозга, слабости сердца и замедлению метаболизма. [110] Клеточные механизмы, препятствующие накоплению транспозонов, по-видимому, частично нарушены у саламандр. Некоторые виды с крупнейшими геномами утратили способность к метаморфозу. Развитие тела происходит медленнее, чем его рост по сравнению с их предками, и прекращается в определенном возрасте, оставляя им эмбриональные черты. Ткани саламандры содержат клетки, которые дифференцируются медленно, слабо или вообще не дифференцируются из-за задержки интронов, что придает им регенеративные свойства, включая регенерирующие части лица и глаз, легких, печени, сердца и даже спинного и головного мозга. и их описывают как «ходячие мешки со стволовыми клетками». [111] [112] [113] Исследования также показали, что у них не развиваются типичные признаки старения и не накапливаются возрастные заболевания, такие как рак. [114]
В человеческом обществе [ править ]
Миф и легенда [ править ]

На протяжении веков вокруг саламандры складывались легенды, многие из которых связаны с огнем. Эта связь, вероятно, связана с тенденцией многих саламандр обитать внутри гниющих бревен. Когда бревно бросали в огонь, саламандра пыталась убежать, что подтверждало веру в то, что саламандры были созданы из пламени. [115]
Ассоциация саламандры с огнем впервые появилась в древности у Аристотеля ( «История животных» 5, 17) и у Плиния Старшего, который в своей «Естественной истории» (10, 86) писал, что «саламандра настолько холодна, что при контакте гасит огонь». ... Он рвет изо рта молочную жидкость; если эта жидкость касается какой-либо части человеческого тела, у него выпадают все волосы, а кожа меняет цвет и покрывается сыпью». [116] Умение тушить огонь повторяют святой Августин в пятом веке и Исидор Севильский в седьмом веке. [117] [118]

Мифический правитель пресвитер Иоанн якобы имел одеяние, сделанное из якобы волос саламандры, на самом деле из асбестового волокна, известного уже в Древней Греции и Риме ( linum vivum Плиния Старшего Naturalis historia , 19, 4). [119] «Император Индии» имел костюм, сделанный из тысячи шкур; У Папы Александра III была туника, которую он очень ценил, и Уильям Кэкстон (1481 г.) писал: «Это Салемандра berithe wulle, из которой сделаны ткань и пояса, которые нельзя сжечь в огне». [120] Говорят, что саламандра настолько токсична, что, обвивая дерево, она могла отравить плоды и убить любого, кто их съел, а упав в колодец, могла убить всех, кто пил из них. [120]
В своей автобиографии рассказывает Бенвенуто Челлини :
Когда мне было около пяти лет, мой отец сидел один в одной из наших маленьких комнат, пел и играл на скрипке. Там только что закончили стирку, и хорошие дрова все еще горели. Было очень холодно, и он подошел к огню. Затем, когда он смотрел на пламя, его взгляд упал на маленькое животное, похожее на ящерицу, которое весело бегало в самой горячей части огня. Внезапно сообразив, что это такое, он позвонил мне и моей сестре и показал это нам. А потом он дал мне такую жестокую пощечину по ушам, что я закричала и расплакалась. При этом он успокоил меня так любезно, как только мог, и сказал: «Мой дорогой маленький мальчик, я ударил тебя не потому, что ты поступил неправильно. Я сделал это только для того, чтобы вы никогда не забывали, что ящерица, которую вы видели в огне, — это саламандра, и, насколько нам известно, никто никогда ее раньше не видел». [121]
Японская гигантская саламандра была предметом легенд и произведений искусства в Японии (например, укиё-э произведение Утагавы Куниёси ). Знаменитое японское мифологическое существо, известное как каппа , возможно, было вдохновлено этой саламандрой. [122]
Медицинские исследования
саламандр Регенерация конечностей уже давно находится в центре внимания ученых. Первое обширное исследование на клеточном уровне было проведено Винченцо Колуччи в 1886 году. [123] Исследователи пытались выяснить условия, необходимые для роста новых конечностей, и надеются, что такую регенерацию можно будет воспроизвести у людей с помощью стволовых клеток . Аксолотли использовались в исследованиях и были генетически модифицированы таким образом, что в клетках ног присутствует флуоресцентный белок, что позволяет отслеживать процесс деления клеток под микроскопом. Похоже, что после потери конечности клетки собираются вместе, образуя комок, известный как бластема . На первый взгляд это кажется недифференцированным, но клетки, возникшие в коже, позже развиваются в новую кожу, мышечные клетки — в новые мышцы, а хрящевые клетки — в новый хрящ. Только клетки, находящиеся непосредственно под поверхностью кожи, являются плюрипотентными и способны развиваться в клетки любого типа. [124] Исследователи из Австралийского института регенеративной медицины обнаружили, что когда макрофаги были удалены, саламандры потеряли способность к регенерации и вместо этого образовали рубцовую ткань. Если процессы, связанные с формированием новой ткани, можно будет реконструировать в организме человека, возможно, станет возможным излечивать травмы спинного или головного мозга, восстанавливать поврежденные органы и уменьшать рубцы и фиброз после операции. [125]
Пятнистая саламандра (Amblystoma maculatum) живет в симбиотических отношениях с зелеными водорослями, известными как Oophila amblystomatis. Клетки водорослей проникают в клетки тканей по всему телу эмбриона и, по-видимому, избегают отторжения, активируя гены, которые подавляют иммунный ответ эмбриона. Механизм, который можно использовать в лечении аутоиммунных заболеваний у людей. [126]
Бренди [ править ]
В статье 1995 года в словенском еженедельном журнале «Младина» рекламировался бренди из саламандры, ликер , предположительно родом из Словении . Говорят, что он сочетает в себе галлюциногенный эффект с эффектом афродизиака и изготавливается путем помещения нескольких живых саламандр в бочку с ферментирующими фруктами. Стимулированные алкоголем, они выделяют токсичную слизь для защиты и в конечном итоге умирают. не только вызывают галлюцинации, но и Говорят, что нейротоксины , присутствующие в напитке, вызывают сильное сексуальное возбуждение . [127]
Более поздние исследования словенского антрополога Михи Козорога ( Университет Любляны ) рисуют совершенно иную картину: саламандра в бренди, похоже, традиционно рассматривалась как примесь , вызывающая ухудшение здоровья. Его также использовали как термин клеветы. [128]
Ссылки [ править ]
Цитаты [ править ]
- ^ Андерсон, Дж. С. (2012). «Окаменелости, молекулы, время расхождения и происхождение Salamandroidea» . Труды Национальной академии наук . 109 (15): 5557–5558. Бибкод : 2012PNAS..109.5557A . дои : 10.1073/pnas.1202491109 . ПМЦ 3326514 . ПМИД 22460794 .
- ↑ Перейти обратно: Перейти обратно: а б с Фрост, Даррел Р. (2023) [1998]. «Каудата» . Виды амфибий мира . Американский музей естественной истории . Архивировано из оригинала 22 марта 2023 года . Проверено 22 марта 2023 г.
- ^ Следы жизни побережья Джорджии: раскрытие невидимой жизни растений и животных
- ^ «Недостающие части? Раскрыт секрет регенерации саламандры» . Живая наука . 20 мая 2013 г. Архивировано из оригинала 16 июля 2018 г. . Проверено 15 июля 2018 г.
- ^ Чжун, Цзисин; Айрес, Рита; Цисиос, Георгиос; Скуфа, Евангелия; Брандт, Керстин; Сандовал-Гузман, Татьяна; Ацтекин, Джан (10 октября 2023 г.). «Многовидовой атлас разрешает парадокс развития и регенерации конечностей аксолотля» . Природные коммуникации . 14 (1): 6346. Бибкод : 2023NatCo..14.6346Z . doi : 10.1038/s41467-023-41944-w . ПМЦ 10564727 . ПМИД 37816738 .
- ^ Харпер, Дуглас. «саламандра» . Интернет-словарь этимологии . Проверено 22 марта 2023 г.
- ^ Лидделл, Генри Джордж; Скотт, Роберт (1940). «σαλαμάνδρα» . Греко-английский лексикон . Цифровая библиотека Персея. Архивировано из оригинала 12 марта 2023 года . Проверено 22 марта 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Ланца, Б.; Ванни, С.; Нистри, А. (1998). Коггер, Х.Г.; Цвайфель, Р.Г. (ред.). Энциклопедия рептилий и амфибий . Академическая пресса. стр. 60–68. ISBN 978-0-12-178560-4 .
- ^ Повторяющиеся экологические и жизненные циклы делают саламандр идеальной моделью для эволюции и развития.
- ^ Обмен веществ, газообмен и кислотно-щелочной баланс гигантских саламандр.
- ^ Полевой справочник по амфибиям и рептилиям Калифорнии: исправленное издание
- ^ Стеббинс и Коэн (1995), с. 3
- ^ Лопес, Карл Х.; Броди, Эдмунд Д. младший (1972). «Функция реберных бороздок у саламандр (амфибий, уроделов)». Журнал герпетологии . 11 (3): 372–374. дои : 10.2307/1563252 . JSTOR 1563252 .
- ↑ Перейти обратно: Перейти обратно: а б с Стеббинс и Коэн (1995), стр. 26–30.
- ↑ Перейти обратно: Перейти обратно: а б Стеббинс и Коэн (1995), стр. 10–16.
- ^ Броди, Эдмунд Д. младший (1968). «Исследования кожного токсина взрослого грубокожего тритона Taricha granulosa ». Копейя . 1968 (2): 307–313. дои : 10.2307/1441757 . JSTOR 1441757 . S2CID 52235877 .
- ↑ Перейти обратно: Перейти обратно: а б Уэллс, Кентвуд, Д. (2010). Экология и поведение амфибий . Издательство Чикагского университета. стр. 411–417. ISBN 978-0-226-89333-4 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Леон, Эсекьель Гонсалес; Рамирес-Пинилья, Марта Патрисия (2009). «Ментальная железа Bolitoglossa nicefori (Caudata: Plethodontidae)» . Земноводные-рептилии . 30 (4): 561–569. дои : 10.1163/156853809789647013 .
- ^ Стеббинс и Коэн (1995), стр. 37–40.
- ^ Стеббинс и Коэн (1995), стр. 42–44.
- ^ Прзырембель, К.; Келлер, Б.; Ноймайер, К. (1995). «Трихроматическое цветовое зрение саламандры ( Salamandra salamandra )». Журнал сравнительной физиологии . 176 (4): 575–586. дои : 10.1007/BF00196422 . S2CID 749622 .
- ^ Шох, Райнер Р. (19 марта 2014 г.). Эволюция земноводных: жизнь ранних наземных позвоночных . Джон Уайли и сыновья. ISBN 9781118759134 . Проверено 15 июля 2018 г. - через Google Книги.
- ^ «Исследователи показывают, как развивался слух» . sciencedaily.com . Архивировано из оригинала 16 июля 2018 года . Проверено 15 июля 2018 г.
- ^ Беккер, Р.П.; Ломбард, RE (1977). «Структурные корреляты функции «жаберной мышцы» амфибий». Исследования клеток и тканей . 175 (4): 499–522. дои : 10.1007/bf00222415 . ПМИД 830429 . S2CID 25650919 .
- ^ Стеббинс и Коэн (1995), стр. 69–72.
- ↑ Перейти обратно: Перейти обратно: а б Губачек Ю.; Шугеркова, М.; Гвождик, Л. (2019). «Подводное звучание у симпатрических тритонов варьируется не между видами» . ПерДж . 7 : е6649. дои : 10.7717/peerj.6649 . ПМК 6441559 . ПМИД 30944780 .
- ^ Биология позвоночных: систематика, таксономия, естественная история и сохранение
- ^ Стеббинс и Коэн (1995), стр. 76–77.
- ↑ Перейти обратно: Перейти обратно: а б Стеббинс и Коэн (1995), стр. 17–25.
- ^ Коггер и Сомнение (1998), стр. 74–75.
- ^ Какехаши, Рёске; Курабаяси, Ацуши (2021). «Закономерности естественного отбора генов, кодирующих митохондриальные белки, у безлегких саламандр: расслабленный очищающий отбор и наличие положительно отобранных кодоновых сайтов в семействе Plethodontidae» . Международный журнал геномики . 2021 : 1–12. дои : 10.1155/2021/6671300 . ПМК 8053045 . ПМИД 33928143 .
- ^ Тэйвс, Дэниел П. (1974). «Дыхательные механизмы водной саламандры Amphiuma tridactylum». Копейя . 1974 (4): 917–920. дои : 10.2307/1442591 . JSTOR 1442591 .
- ^ Рогге, Джессика Р.; Варкентин, Карен М. (15 ноября 2008 г.). «Внешние жабры и адаптивное поведение эмбриона способствуют синхронному развитию и пластичности вылупления при ограничении дыхания» . Журнал экспериментальной биологии . 211 (22): 3627–3635. дои : 10.1242/jeb.020958 . ISSN 0022-0949 . ПМИД 18978228 .
- ^ Дуэллман, Уильям Эдвард (1994). Биология амфибий . Издательство Университета Джонса Хопкинса.
- ^ В., Кардонг, Кеннет (2012). Позвоночные животные: сравнительная анатомия, функции, эволюция . МакГроу-Хилл. ISBN 9780073524238 . OCLC 939087630 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Залиско, Эдвард Дж. (2015). Сравнительная анатомия позвоночных: руководство по лабораторному вскрытию . Макгроу-Хилл Образование. ISBN 9780077657055 . OCLC 935173274 .
- ↑ Перейти обратно: Перейти обратно: а б с Хейрстон, Нельсон Г. (1949). «Местное распространение и экология многодонтидных саламандр южных Аппалачей». Экологические монографии . 19 (1): 47–73. Бибкод : 1949ЭкоМ...19...47Н . дои : 10.2307/1943584 . JSTOR 1943584 .
- ^ «Плетодонтиды» . АмфибияВеб. Архивировано из оригинала 22 февраля 2014 года . Проверено 9 февраля 2014 г.
- ^ Коппес, Стив (24 сентября 2009 г.). «Он невкусный, он мой брат» . Спросите биолога . Университет штата Аризона. Архивировано из оригинала 22 февраля 2014 года . Проверено 8 февраля 2014 г.
- ^ « Десмогнат квадратный » . АмфибияВеб. Архивировано из оригинала 22 февраля 2014 года . Проверено 9 февраля 2014 г.
- ^ Кардонг (2009), стр. 505–506.
- ^ Стеббинс и Коэн (1995), стр. 57–58.
- ^ Стеббинс и Коэн (1995), стр. 58–60.
- ^ Ларсен, Джон Х. младший; Гатри, Дэн Дж. (1975). «Система питания наземных тигровых саламандр ( Ambystoma tigrinum melanostictum baird)». Журнал морфологии . 147 (2): 137–153. дои : 10.1002/jmor.1051470203 . ПМИД 30309060 . S2CID 52959566 .
- ^ Дебан, С.М.; Уэйк, ДБ; Рот, Г. (1997). «Саламандра с баллистическим языком». Природа . 389 (6646): 27–28. Бибкод : 1997Natur.389...27D . дои : 10.1038/37898 . S2CID 205026166 .
- ^ Дебан, С.М.; О'Рейли, У. Дике (2007). «Чрезвычайно мощная проекция языка у многодонтидных саламандр» . Журнал экспериментальной биологии . 210 (4): 655–667. дои : 10.1242/jeb.02664 . ПМИД 17267651 .
- ↑ Перейти обратно: Перейти обратно: а б Регал, Филип Дж. (1966). «Специализации питания и классификация наземных саламандр». Эволюция . 20 (3): 392–407. дои : 10.2307/2406638 . JSTOR 2406638 . ПМИД 28562974 .
- ^ Хилл, РЛ; Мендельсон-младший; Стабиле, Дж.Л. (2015). «Прямое наблюдение и обзор травоядных Sirenidae (Amphibia: Caudata)». Юго-восточный натуралист . 14 : N5–N9. дои : 10.1656/058.014.0104 . S2CID 86233204 .
- ^ Стеббинс и Коэн (1995), стр. 110–120.
- ^ Мейсон, Дж. Рассел; Рабин, Майкл Д.; Стивенс, Дэвид А. (1982). «Обусловленное отвращение вкуса: выделения кожи, используемые тигровыми саламандрами, Ambystoma tigrinum, для защиты ». Копейя . 1982 (3): 667–671. дои : 10.2307/1444668 . JSTOR 1444668 .
- ^ Броди, Эдмунд Д. младший; Сматреск, Нил Дж . (1990). «Арсенал огненных саламандр против хищников: распыление секрета из спинных кожных желез, находящихся под высоким давлением». Герпетологика . 46 (1): 1–7. JSTOR 3892595 .
- ^ Хейсс, Э.; Натчев, Н.; Салабергер, Д.; Гумпенбергер, М.; Рабансер, А.; Вейсграм, Дж. (2010). «Навреди себе, чтобы навредить своему врагу: новые сведения о функции причудливого механизма защиты от хищников у саламандр Pleurodeles waltl » . Журнал зоологии . 280 (2): 156–162. дои : 10.1111/j.1469-7998.2009.00631.x .
- ↑ Перейти обратно: Перейти обратно: а б Ховард, Ронни Р.; Броди, Эдмунд Д. младший (1973). «Бейтсовский миметический комплекс у саламандр: реакция птичьих хищников». Герпетологика . 29 (1): 33–41. JSTOR 3891196 .
- ^ Кухта, СР; Кракауэр, АХ; Синерво, Б (2008). «Почему у желтоглазой энсатины желтые глаза? Бейтсовская мимикрия тихоокеанских тритонов (род Trachia ) со стороны саламандры Ensatina eschscholtzii xanthoptica » . Эволюция . 62 (4): 984–990. дои : 10.1111/j.1558-5646.2008.00338.x . ПМИД 18248632 . S2CID 998486 .
- ^ Котт, 1940. Страница 204.
- ^ Монаган, Джеймс Р.; Эпп, Леонард Г.; Путта, Шрикришна; Пейдж, Роберт Б.; Уокер, Джон А.; Бичи, Крис К.; Чжу, Вэй; Пао, Джеральд М.; Верма, Индер М.; Хантер, Тони; Брайант, Сьюзен В.; Гардинер, Дэвид М.; Харкинс, Тим Т.; Восс, С. Рэндал (2009). «Анализ транскрипции на микрочипах и кДНК во время нервно-зависимой регенерации конечностей» . БМК Биология . 7 (1): 1. дои : 10.1186/1741-7007-7-1 . ПМК 2630914 . ПМИД 19144100 .
- ^ Шох, Райнер Р.; Вернебург, Ральф; Фойгт, Себастьян (2020). «Триасовый ствол-саламандра из Кыргызстана и происхождение саламандр» . Труды Национальной академии наук . 117 (21): 11584–11588. Бибкод : 2020PNAS..11711584S . дои : 10.1073/pnas.2001424117 . ПМК 7261083 . ПМИД 32393623 .
- ^ Эванс, SE; Милнер, Арканзас; Массетт, Ф. (1988). «Самые ранние известные саламандры (амфибия, хвостатые): записи средней юры Англии». Геобиос . 21 (5): 539–552. Бибкод : 1988Geobi..21..539E . дои : 10.1016/s0016-6995(88)80069-x .
- ↑ Перейти обратно: Перейти обратно: а б Джонс, Марк Э.Х.; Бенсон, Роджер Б.Дж.; Скутщас, Павел; Хилл, Люси; Панчироли, Эльза; Шмитт, Армин Д.; Уолш, Стиг А.; Эванс, Сьюзен Э. (11 июля 2022 г.). «Окаменелости средней юры документируют раннюю стадию эволюции саламандр» . Труды Национальной академии наук . 119 (30): e2114100119. Бибкод : 2022PNAS..11914100J . дои : 10.1073/pnas.2114100119 . ISSN 0027-8424 . ПМЦ 9335269 . ПМИД 35858401 .
- ^ Гао, Кэ-Цинь; Шубин, Нил Х. (2012). «Позднеюрский саламандроид из западного Ляонина, Китай» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (15): 5767–5772. Бибкод : 2012PNAS..109.5767G . дои : 10.1073/pnas.1009828109 . ПМЦ 3326464 . ПМИД 22411790 .
- ^ Марьянович, Д.; Лаурин, М. (2014). «Обновленное палеонтологическое древо лисамфибий с комментариями по анатомии саламандр юрской кронной группы (Urodela)». Историческая биология . 26 (4): 535–550. Бибкод : 2014HBio...26..535M . дои : 10.1080/08912963.2013.797972 . S2CID 84581331 .
- ^ Shekhovtsov, Sergei V.; Bulakhova, Nina A.; Tsentalovich, Yuri P.; Zelentsova, Ekaterina A.; Meshcheryakova, Ekaterina N.; Poluboyarova, Tatiana V.; Berman, Daniil I. (12 November 2021). "Biochemical Response to Freezing in the Siberian Salamander Salamandrella keyserlingii" . Biology . 10 (11): 1172. doi : 10.3390/biology10111172 . PMC 8614755 . PMID 34827165 .
- ^ Уэллс, Кентвуд Д. (2007). Экология и поведение амфибий . Издательство Чикагского университета. п. 130. ИСБН 978-0-226-89334-1 .
- ^ Элмер, КР; Бонетт, РМ; Уэйк, ДБ; Лохид, Южная Каролина (4 марта 2013 г.). «Раннемиоценовое происхождение и загадочная диверсификация южноамериканских саламандр» . Эволюционная биология BMC . 13 (1): 59. Бибкод : 2013BMCEE..13...59E . дои : 10.1186/1471-2148-13-59 . ПМК 3602097 . ПМИД 23497060 .
- ^ «Первое в истории открытие саламандры в янтаре проливает свет на эволюцию Карибских островов | Новости и исследовательские коммуникации | Университет штата Орегон» . oregonstate.edu . 17 августа 2015 года. Архивировано из оригинала 19 февраля 2017 года . Проверено 25 марта 2016 г.
- ^ Болес, МЫ (1997). «Ископаемые певчие птицы (Воробьиные) из раннего эоцена Австралии». Эму . 97 (1): 43–50. Бибкод : 1997EmuAO..97...43B . дои : 10.1071/MU97004 . ISSN 0158-4197 .
- ^ «Виды по количеству» . АмфибияВеб. Архивировано из оригинала 12 января 2021 года . Проверено 11 января 2021 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и Нэйш, Даррен (1 октября 2013 г.). «Удивительный мир саламандр» . Научный американец . Архивировано из оригинала 16 января 2014 года . Проверено 14 января 2014 г.
- ^ Брэд Шаффер; Оскар Флорес-Виллела; Габриэла Парра-Олеа; Дэвид Уэйк (2004). «Амбистома андерсони». Красный список видов, находящихся под угрозой исчезновения МСОП. Версия 2013.2. Международный союз охраны природы
- ^ Брэдли, Дж. Гэвин; Исон, Перри К. (2018). « Eurycea lucifuga (Пещерная саламандра) Вокализация» . Герпетологическое обозрение . 49 (4): 725.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ↑ Перейти обратно: Перейти обратно: а б с Стеббинс и Коэн (1995), стр. 143–154.
- ^ Бюшинг, Кристина Д. (19 июля 2019 г.). Химические сигналы у позвоночных 14 . Спрингер. ISBN 9783030176167 .
- ^ «Изучение влияния систем спаривания на размер семенников у саламандр» . Архивировано из оригинала 16 февраля 2023 года . Проверено 16 февраля 2023 г.
- ^ Цзя, Цзя; Андерсон, Джейсон С.; Гао, Кэ-Цинь (2021). «Среднеюрские стволовые гинобииды из Китая проливают свет на эволюцию базальных саламандр» . iScience . 24 (7). Бибкод : 2021iSci...24j2744J . дои : 10.1016/j.isci.2021.102744 . ПМЦ 8264161 . ПМИД 34278256 .
- ^ «Эволюционная репродуктивная морфология амфибий» (PDF) . Архивировано (PDF) из оригинала 7 марта 2023 года . Проверено 15 февраля 2023 г.
- ^ Боверо, С.; Сотджиу, Г.; Кастеллано, С.; Джакома, К. (2003). «Возрастной и половой диморфизм в популяции Euproctus platycephalus (Caudata: Salamandridae) с Сардинии». Копейя . 2003 : 149–154. doi : 10.1643/0045-8511(2003)003[0149:AASDIA]2.0.CO;2 . S2CID 85909830 .
- ^ Уэллс, Кентвуд Д. (15 февраля 2010 г.). Экология и поведение амфибий . Издательство Чикагского университета. ISBN 9780226893334 .
- ^ Ваги, Балаж; Марш, Дэниел; Катона, Грегори; Вегвари, Жолт; Фреклтон, Роберт П.; Лайкер, Андраш; Секели, Тамаш (5 октября 2022 г.). «Эволюция родительской заботы у саламандр - Природа» . Научные отчеты . 12 (1): 16655. doi : 10.1038/s41598-022-20903-3 . ПМЦ 9535019 . ПМИД 36198742 .
- ^ Репродуктивная биология и филогения Уроделы
- ↑ Перейти обратно: Перейти обратно: а б Стеббинс и Коэн (1995), стр. 175–179.
- ^ «Плетодонтиды» . АмфибияВеб. Архивировано из оригинала 9 июня 2015 года . Проверено 9 января 2014 г.
- ↑ Перейти обратно: Перейти обратно: а б Киёнага, Робин Р. «Метаморфоза против неотении (педоморфоза) у саламандр (Caudata)» . Архивировано из оригинала 1 декабря 2008 года . Проверено 4 мая 2008 г.
- ↑ Перейти обратно: Перейти обратно: а б Сандовал-Конте, Адриана; Пинеда, Эдуардо; Агилар-Лопес, Хосе Л. (2012). «В поисках видов, находящихся под угрозой исчезновения: текущая ситуация с двумя крошечными видами саламандр в неотропических горах Мексики» . ПЛОС ОДИН . 7 (4): e34023. Бибкод : 2012PLoSO...734023S . дои : 10.1371/journal.pone.0034023 . ПМЦ 3317776 . ПМИД 22485155 .
- ↑ Перейти обратно: Перейти обратно: а б Тэпли, Бенджамин; Брэдфилд, Кей С.; Майклс, Кристофер; Бунгард, Майк (24 июля 2015 г.). «Амфибии и программы сохранения разведения: все ли амфибии, находящиеся под угрозой исчезновения, принадлежат ковчегу?». Биоразнообразие и сохранение . 24 (11): 2625–2646. Бибкод : 2015BiCon..24.2625T . дои : 10.1007/s10531-015-0966-9 . ISSN 0960-3115 . S2CID 11824410 .
- ^ Фонтан, Генри (16 февраля 2009 г.). «Еще одна амфибия в опасности: саламандры» . Нью-Йорк Таймс . Архивировано из оригинала 26 ноября 2013 года . Проверено 28 июня 2013 г.
- ^ «Китайская гигантская саламандра» . Сохранение ZSL . Зоологическое общество Лондона. Архивировано из оригинала 28 июня 2013 года . Проверено 21 июля 2013 г.
- ^ Уилер, Бенджамин А.; Прозен, Итан; Матис, Алисия; Уилкинсон, Роберт Ф. (2003). «Сокращение популяции долгоживущей саламандры: более чем 20-летнее исследование магов ада Cryptobranchus alleganiensis ». Биологическая консервация . 109 (1): 151–156. Бибкод : 2003BCons.109..151W . дои : 10.1016/s0006-3207(02)00136-2 .
- ^ Уилер, Бенджамин А.; МакКаллум, Малкольм Л.; Траут, Стэнли Э. (2002). «Аномалии у Озарка-повелителя ада ( Ctyptobranchm alleganiensis Bishopi ) в Арканзасе: сравнение двух рек с исторической точки зрения» . Журнал Академии наук Арканзаса . 58 : 250–252.
- ^ «Первое в мире разведение магов ада Озарка в неволе» . Наука Дейли . 1 декабря 2011 года. Архивировано из оригинала 11 мая 2013 года . Проверено 21 июля 2013 г.
- ^ «Безлегкие саламандры Мексики» . EDGE: Эволюционно обособленный вид, находящийся под угрозой исчезновения во всем мире . Зоологическое общество Лондона. Архивировано из оригинала 29 мая 2013 года . Проверено 21 июля 2013 г.
- ^ «Грань существования» . ГРАНЬ Существования . Архивировано из оригинала 17 ноября 2015 года . Проверено 29 октября 2015 г.
- ↑ Перейти обратно: Перейти обратно: а б Контрерас, Виктория; Мартинес-Мейер, Энрике; Валиенте, Эльза; Самбрано, Луис (1 декабря 2009 г.). «Недавнее сокращение и потенциальное распространение в последней зоне остатков микроэндемичного мексиканского аксолотля ( Ambystoma mexicanum )». Биологическая консервация . 142 (12): 2881–2885. Бибкод : 2009BCons.142.2881C . doi : 10.1016/j.biocon.2009.07.008 .
- ^ Парра-Олеа, Г.; Самудио, КР; Рекуэро, Э.; Агилар-Мигель, X.; Уакуз, Д.; Самбрано, Л. (1 февраля 2012 г.). «Генетика сохранения находящихся под угрозой исчезновения мексиканских аксолотлей ( Амбистома )». Охрана животных . 15 (1): 61–72. Бибкод : 2012AnCon..15...61P . дои : 10.1111/j.1469-1795.2011.00488.x . ISSN 1469-1795 . S2CID 46992721 .
- ^ Алькарас, Гильермина; Лопес-Портела, Харини; Роблес-Мендоса, Сесилия (1 февраля 2015 г.). «Реакция местного находящегося под угрозой исчезновения аксолотля Ambystoma mexicanum (Amphibia) на экзотического хищника-рыбу». Гидробиология . 753 (1): 73–80. дои : 10.1007/s10750-015-2194-4 . ISSN 0018-8158 . S2CID 17468971 .
- ^ Фигил, Честер (2013). «Криоконсервация спермы аксолотля AmbystomA MexicAnum: последствия для сохранения» (PDF) . Герпетологическая охрана и биология . Архивировано (PDF) из оригинала 22 марта 2016 г. Проверено 26 октября 2015 г.
- ^ Марсец, Рут; Бемент, Ханна. «Сохранение саламандр» . Сохранение амфибий . Мемфисский зоопарк. Архивировано из оригинала 31 июля 2013 года . Проверено 21 июля 2013 г.
- ^ Райс, Эдвард Лоранус (1935). Введение в биологию . Бостон, Массачусетс, США: Джинн и компания. п. 253.
- ↑ Перейти обратно: Перейти обратно: а б Ларсон, А.; Диммик, В. (1993). «Филогенетические взаимоотношения семейств саламандр: анализ соответствия морфологических и молекулярных признаков». Герпетологические монографии . 7 (7): 77–93. дои : 10.2307/1466953 . JSTOR 1466953 .
- ^ Блэкберн, Дэвид К.; Уэйк, Дэвид Б. (23 декабря 2011 г.). «Класс Амфибия Серые, 1825 г. В: Чжан, З.-К. (Ред.) Биоразнообразие животных: Очерк классификации более высокого уровня и обзор таксономического богатства». Зоотакса . 3148 (1). дои : 10.11646/zootaxa.3148.1.8 .
- ^ Сан-Мауро, Диего; Вансес, Мигель; Алькобендас, Марина; Зардоя, Рафаэль; Мейер, Аксель (2005). «Первоначальное разнообразие современных амфибий предшествовало распаду Пангеи» (PDF) . Американский натуралист . 165 (5): 590–599. дои : 10.1086/429523 . ПМИД 15795855 . S2CID 17021360 .
- ^ Сан-Мауро, Д. (2010). «Многолокусная временная шкала происхождения современных амфибий». Молекулярная филогенетика и эволюция . 56 (3): 554–561. Бибкод : 2010МОЛПЭ..56..554С . дои : 10.1016/j.ympev.2010.04.019 . ПМИД 20399871 .
- ^ Марьянович Д., Лаурин М. (2007). «Ископаемые, молекулы, время расхождения и происхождение лисамфибий» . Систематическая биология . 56 (3): 369–388. дои : 10.1080/10635150701397635 . ПМИД 17520502 .
- ^ Шох, Райнер Р.; Вернебург, Ральф; Фойгт, Себастьян (26 мая 2020 г.). «Триасовый ствол-саламандра из Кыргызстана и происхождение саламандр» . Труды Национальной академии наук . 117 (21): 11584–11588. Бибкод : 2020PNAS..11711584S . дои : 10.1073/pnas.2001424117 . ISSN 0027-8424 . ПМК 7261083 . ПМИД 32393623 .
- ^ де Буффрениль В., Кановиль А., Эванс С.Е., Лорин М. (2014). «Гистологическое исследование караурид, старейших известных (стволовых) уроделей». Историческая биология . 27 (1): 109–114. дои : 10.1080/08912963.2013.869800 . S2CID 83557507 .
- ^ Марьянович Д., Лаурин М. (2014). «Обновленное палеонтологическое древо лисамфибий с комментариями по анатомии саламандр юрской кронной группы (Urodela)». Историческая биология . 26 (4): 535–550. Бибкод : 2014HBio...26..535M . дои : 10.1080/08912963.2013.797972 . S2CID 84581331 .
- ^ Р. Александр Пайрон; Джон Дж. Винс (2011). «Крупномасштабная филогения земноводных, включающая более 2800 видов, и пересмотренная классификация современных лягушек, саламандр и червяг» . Молекулярная филогенетика и эволюция . 61 (2): 543–583. Бибкод : 2011МОЛПЭ..61..543А . дои : 10.1016/j.ympev.2011.06.012 . ПМИД 21723399 .
- ^ Сунь, Ченг; Шепард, Дональд Б.; Чонг, Ребекка А.; Лопес Арриаса, Хосе; Холл, Кэтрин; Касто, Тодд А.; Фешотт, Седрик; Поллок, Дэвид Д.; Мюллер, Рэйчел Локридж (1 января 2012 г.). «LTR-ретротранспозоны способствуют геномному гигантизму у плетодонтидных саламандр» . Геномная биология и эволюция . 4 (2): 168–183. дои : 10.1093/gbe/evr139 . ПМК 3318908 . ПМИД 22200636 .
- ^ Элева, Ахмед; Ван, Хэн; Талавера-Лопес, Карлос; Ховен, Альберто; Брито, Гонсалу; Кумар, Ануп; Хамид, Л. Шахул; Пенрад-Мобаед, май; Яо, Зею (22 декабря 2017 г.). «Чтение и редактирование генома Pleurodeles Waltl раскрывает новые особенности регенерации четвероногих» . Природные коммуникации . 8 (1): 2286. Бибкод : 2017NatCo...8.2286E . дои : 10.1038/s41467-017-01964-9 . ISSN 2041-1723 . ПМЦ 5741667 . ПМИД 29273779 .
- ^ Новошилов, Сергей; Шлойсниг, Зигфрид; Фей, Цзи-Фэн; Даль, Андреас; Панг, Энди У.К.; Пиппель, Мартин; Винклер, Силке; Хасти, Алекс Р.; Янг, Джордж; Росцито, Джулиана Г.; Сокол, Франциско; Кнапп, Дуня; Пауэлл, Шон; Круз, Альфредо; Цао, Хан; Хаберманн, Бьянка; Хиллер, Майкл; Танака, Элли М.; Майерс, Юджин В. (2018). «Геном аксолотля и эволюция ключевых регуляторов тканевого образования» . Природа . 554 (7690): 50–55. Бибкод : 2018Natur.554...50N . дои : 10.1038/nature25458 . hdl : 21.11116/0000-0003-F659-4 . ПМИД 29364872 .
- ^ Сан, К.; Мюллер, Р.Л. (17 июля 2014 г.). «Последовательности генома Хеллбендера проливают свет на расширение генома у основания коронованных саламандр» . Геномная биология и эволюция . 6 (7): 1818–1829. дои : 10.1093/gbe/evu143 . ПМК 4122941 . ПМИД 25115007 .
- ^ Сешнс, Стэнли К.; Уэйк, Дэвид Б. (июнь 2021 г.). «Вечно молодой: связь регенерации и размера генома саламандр». Динамика развития . 250 (6): 768–778. дои : 10.1002/dvdy.279 . ПМИД 33320991 .
- ^ Фокс, Дуглас (февраль 2022 г.). «Мусорная ДНК деформирует тела саламандр» . Научный американец . 326 (2): 40.
- ^ Фокс, Дуглас (29 сентября 2022 г.). «Живые тайны: у этого существа в 38 раз больше ДНК, чем у вас» . Новости науки исследуют .
- ^ Преимущества клеток «зомби»: стареющие клетки способствуют регенерации саламандр.
- ^ Эшкрофт, Фрэнсис (2002). Жизнь в крайностях: наука выживания . Издательство Калифорнийского университета. п. 112 . ISBN 978-0-520-22234-2 .
- ^ Плиний Старший (ок. 100). Естественная история . стр. Книга 10, 86. Архивировано из оригинала 18 октября 2008 года . Проверено 8 января 2014 г.
- ^ Святой Августин (ок. 500 г.). Город Божий . с. Книга 21, 4.
- ^ Исидор Севильский (ок. 700). Этимологии . стр. Книга 12, 4:36. Архивировано из оригинала 18 октября 2008 года . Проверено 8 января 2014 г.
- ^ Браун, Клэр (2003). «Шерсть саламандры: историческое свидетельство использования текстиля, сотканного из асбестового волокна». Текстильная история . 34 : 64–73. дои : 10.1179/004049603235001454 . S2CID 191599472 .
- ↑ Перейти обратно: Перейти обратно: а б Уайт, TH (1992) [1954]. Книга зверей: перевод латинского бестиария двенадцатого века . Алан Саттон . стр. 183–184. ISBN 978-0-7509-0206-9 .
- ^ Челлини, Бенвенуто (1998). Автобиография Бенвенуто Челлини . Англия: Книги Пингвинов. п. 34. ISBN 0140447180 .
- ^ Режиссер: Дункан Чард (14 февраля 2012 г.). «Речные монстры». Программа 6, серия 3 . ИТВ . ИТВ1 .
- ^ Холланд, Николас (2021), «Мемуары Виченцо Колуччи 1886 года, О регенерации конечностей и хвоста у тритонов, аннотированные и переведенные на английский язык как: О регенерации конечностей и хвоста у саламандр», Европейский зоологический журнал , 88 : 837 –890, дои : 10.1080/24750263.2021.1943549
- ^ Кейм, Брэндон (1 июля 2009 г.). «Открытие саламандры может привести к регенерации конечностей человека» . Проводной . Проверено 7 мая 2010 г.
- ^ «Является ли иммунная система саламандр ключом к регенерации?» . ScienceDaily . Архивировано из оригинала 6 декабря 2013 года . Проверено 21 мая 2013 г.
- ^ Симбиотические отношения саламандры и водорослей могут вдохновить на создание новых лекарств.
- ^ Голдсмит, Ричард (11 июня 2010 г.). «Сногсшибательные галлюциногенные напитки» . Фокс Ньюс . Архивировано из оригинала 5 декабря 2014 года . Проверено 2 января 2014 г.
- ^ Козорог, Миха (2003). «Бренди Саламандра:« психоделический напиток »между мифом СМИ и практикой домашней дистилляции алкоголя в Словении» . Обзор антропологии Восточной Европы . 21 (1): 63–71. Архивировано из оригинала 31 марта 2014 года . Проверено 22 марта 2014 г.
Цитируемые тексты [ править ]
- Коггер, Х.Г.; Цвайфель, Р.Г. , ред. (1998). Энциклопедия рептилий и земноводных (2-е изд.). Академическая пресса. ISBN 978-0-12-178560-4 .
- Котт, Хью Б. (1940). Адаптивная окраска животных . Издательство Оксфордского университета.
- Дорит, РЛ; Уокер, ВФ; Барнс, Р.Д. (1991). Зоология . Издательство Колледжа Сондерса. ISBN 978-0-03-030504-7 .
- Кардонг, Кеннет В. (2009). Позвоночные животные: сравнительная анатомия, функции, эволюция (5-е изд.). МакГроу-Хилл. ISBN 978-0-07-304058-5 .
- Стеббинс, Роберт С .; Коэн, Натан В. (1995). Естественная история амфибий . Издательство Принстонского университета. ISBN 978-0-691-03281-8 .
Внешние ссылки [ править ]
- Древо жизни: Каудата
- Саламандра Галерея
- Каудата Культура
- Critter Crossings: туннели Саламандры в Министерстве транспорта США
- ArchéoZooThèque: рисунок скелета уродели : доступен в векторном формате, формате изображений и PDF.




Хвостатые семейства по подотряду |
|---|

| Базы данных органов управления : Национальные |
|---|