Плетодонтиды
| Безлегочная саламандра Временной диапазон: | |
|---|---|
 | |
| Калифорнийская стройная саламандра ( Batrachoseps attenuatus ) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Амфибия |
| Заказ: | Уродела |
| Подотряд: | Саламандроидея |
| Семья: | Плетодонтиды Грей , 1850 г. |
| Подгруппы | |
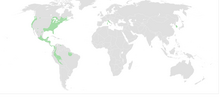 | |
| Местное распространение плетодонтид (зеленым цветом) | |
Plethodontidae , или безлегочные саламандры , — семейство саламандр . [1] [2] Безлегочные саламандры, насчитывающие более 500 видов, на сегодняшний день являются самым большим по разнообразию семейством саламандр. Большинство видов обитают в Западном полушарии , от Британской Колумбии до Бразилии. встречаются только два существующих рода В Восточном полушарии : Speleomantes (родной для Сардинии и материковой Европы к югу от Альп) и Karsenia (родной для Южной Кореи). [3]
Биология
[ редактировать ]
Взрослые безлегочные саламандры имеют четыре конечности, с четырьмя пальцами на передних конечностях и обычно с пятью на задних конечностях. У многих видов спаривание и размножение происходят исключительно на суше. Соответственно, у многих видов также отсутствует водная личиночная стадия - явление, известное как прямое развитие, при котором потомство вылупляется полностью сформировавшимися миниатюрными взрослыми особями. Прямое развитие коррелирует с изменениями в особенностях развития плетодонтид по сравнению с другими семействами саламандр, включая увеличение размера яиц и продолжительности эмбрионального развития. Кроме того, эволюционная утрата водной личиночной стадии связана с уменьшением зависимости размножения от водной среды обитания. Снятие этого ограничения позволило широкомасштабную колонизацию и диверсификацию в широком количестве наземных местообитаний, что является свидетельством высокого успеха и распространения Plethodontidae. [4]
Несмотря на отсутствие легких, некоторые из них могут вырасти довольно большими. Самый крупный вид безлегочных саламандр, саламандра ложного ручья Белла , может достигать длины 36 см (14 дюймов). [5]
У многих видов есть метательный язык и подъязычный аппарат , которыми они могут стрелять почти на всю длину тела с высокой скоростью, чтобы поймать добычу.
Если измерять в индивидуальном количестве, то там, где они встречаются, они являются очень успешными животными. В некоторых местах они составляют доминирующую биомассу позвоночных. [6] обитает 1,88 миллиарда особей южной красноспинной саламандры По оценкам , только в одном районе Национального леса Марка Твена , а биомасса составляет около 1400 тонн. [7] Из-за своих скромных размеров и низкого метаболизма они способны питаться такой добычей, как коллемболы , которые обычно слишком малы для других наземных позвоночных. Это дает им доступ к целой экологической нише с минимальной конкуренцией со стороны других групп.
Ухаживание и спаривание
[ редактировать ]Плетодонтиды демонстрируют весьма стереотипное и сложное брачное поведение и ритуалы ухаживания, которых нет ни в одном другом семействе саламандр. Брачное поведение, как правило, одинаково для всех плетодонтид и обычно включает в себя ходьбу с расставлением хвоста, при которой самка ориентирует голову у основания хвоста самца, одновременно оседлав хвост своим телом. Самец поворачивает свое тело и помещает капсулу со спермой, известную как сперматофор, на субстрат перед мордой самки. Когда самец ведет самку хвостом по сперматофору, самка опускает свою клоаку на сперматофор и помещает массу спермы внутрь, оставляя основание сперматофора на земле. [8]
У многих видов plethodontidae ритуал ухаживания часто сопровождается передачей мужских феромонов во время ходьбы с расставлением хвоста. В период размножения у самцов вырастают увеличенные передние зубы, которыми они царапают кожу на голове самки в рамках ритуала ухаживания. Впоследствии самец втирает в потертое место феромоны, которые выделяются из подушечки ткани, называемой подбородочной железой, расположенной под подбородком самца. [8] [9] [10] [11]
Феромоны ухаживания значительно увеличивают успех самцов в спаривании по ряду причин. В целом выделения феромонов повышают восприимчивость самок к ухаживанию и передаче спермы. Это не только увеличивает вероятность успешного спаривания с конкретной самкой, но и сокращает продолжительность ухаживания, что важно, поскольку сводит к минимуму вероятность того, что самец будет прерван другими конкурирующими самцами. [12]
В научной литературе, обсуждающей различия между подбородочными железами многодонтидных саламандр, было обнаружено, что самцы плетодонтид имели незначительные различия в высоте и диаметре простых трубчатых желез, а основные различия были обнаружены в диаметре секреторных гранул. Это объясняется тем, что самцы могут спариваться в течение всех месяцев года, а самки откладывают яйца сезонно.
Дыхание
[ редактировать ]Ряд особенностей отличает плетодонтид от других саламандр. Самое главное, что у них нет легких , которые осуществляют дыхание через кожу и ткани, выстилающие рот. [3] Некоторые виды пещерных саламандр неотеничны и сохраняют личиночные жабры даже во взрослом возрасте. У всех остальных взрослых плетодонтид жабры отсутствуют. [13] Плетодонтиды имеют реберные бороздки на туловище. Они помогают сохранять кожу влажной благодаря транспортировке воды по поверхности тела. [14]
Плетодонтидные саламандры почти полностью полагаются на кожное дыхание. [15] Приблизительно 83–93% поглощения кислорода происходит благодаря этому методу. [16] Частота дыхания плетодонтидной саламандры ограничена их SA:V , а более высокие значения SA:V коррелируют с более теплым и влажным климатом. [17]
Плетодонтиды постоянно подвергаются воздействию воздуха или воды, что обеспечивает постоянный газообмен, не ограничиваемый вентиляцией. [15] Поглощение кислорода одинаково в воде и воздухе, при условии, что парциальное давление кислорода одинаково. [18] Кислородная и неокисленная кровь смешиваются в венозной системе, что приводит к тому, что парциальное давление кислорода в сердечной крови обычно бывает низким. [15]
Плетодонтиды могут переносить гипоксию в течение длительных периодов времени за счет снижения скорости метаболизма вместо того, чтобы полагаться на анаэробное кожное дыхание, как первоначально предполагалось. [18]
Было замечено, что у плетодонтид в эмбриональном состоянии развиваются рудиментарные легкие. [19] Зачаток легких развивается аналогично таковому у бесплетодонтидных саламандр в течение первых трех недель развития, а затем начинает регрессировать посредством апоптоза . [19]

Паралог гена SFTPC , который экспрессируется исключительно в легких у других позвоночных, у безлегких саламандр экспрессируется вместо этого в личиночных покровах. Проходя метаморфоз, он исчезает из покровов и появляется у взрослых в ротоглотке. Предполагается, что ген облегчают внелегочное дыхание за счет выработки легочного сурфактантоподобного секрета. [20]
Хеморецепция
[ редактировать ]Еще одной отличительной особенностью является наличие вертикальной щели между ноздрей и верхней губой, известной как «носогубная бороздка». саламандры Борозда выстлана железами и усиливает хеморецепцию , что коррелирует с более высокой степенью развития обонятельной доли и слизистой оболочки носа у плетодонтид. [3] [21] Наличие этой специализированной структуры, вероятно, связано с отсутствием легких у этих саламандр. Хотя у некоторых саламандр с выпадом действительно наблюдаются подобные структуры, они уменьшены в размерах и не расположены возле ноздрей (то есть ноздрей) так же, как плетодонтиды. В связи с тем, что плетодонтиды не могут создавать давление воздуха путем вытеснения воздуха из легких и через ноздри, перед ними стоит задача удаления воды и мусора из носовых ходов, что может значительно ограничить обонятельные процессы. Таким образом, носогубные бороздки устроены таким образом, чтобы обеспечить максимальный дренаж из носа. Борозда более глубокая и узкая непосредственно вокруг ноздрей, а отверстия желез слегка приподняты, что способствует гравитационному потоку жидкости из ноздрей и носовой впадины. Кроме того, носогубные железы по краям ноздрей выделяют жировую пленку, которая дополнительно способствует удалению воды из носовых ходов из-за различий в полярности между водными и липидными выделениями. [21]
Эволюционная история
[ редактировать ]По оценкам, Plethodontidae отделились от своей сестринской группы Amphiumidae вокруг границы K-Pg и диверсифицировались во время палеогена . [22] Регионом происхождения семейства является Северная Америка, причем самые старые из европейских членов семейства известны из среднего миоцена Словакии. [23]
Подсемейства и роды
[ редактировать ]Семейство Plethodontidae состоит из двух современных подсемейств и около 516 [2] до 520 [1] виды разделены на эти роды, составляющие большинство известных видов саламандр: [2]
| Подсемейство | Род, научное название и автор. | Общее имя | Разновидность |
|---|---|---|---|
| Гемидактилии Хэллоуэлл, 1856 г. | |||
| Aquiloeurycea Rovito, Parra-Olea, Recuero и Wake, 2015 г. | 6 - 7 | ||
| Батрахозепс Бонапарт, 1839 г. | Стройные саламандры | 23 | |
| Болитоглосса Дюмериль, Биброн и Дюмериль, 1854 г. | Тропические альпинистские саламандры | 139 - 140 | |
| Брадитритон Уэйк и Элиас, 1983 г. | Саламандра Финка Чиблак | 1 | |
| Хироптеротритон Тейлор, 1944 год. | Саламандры с растопыренными ногами | 23 | |
| Криптотритон Гарсия-Париж и Уэйк, 2000 г. | Скрытые саламандры | 7 | |
| Дендротритон Уэйк и Элиас, 1983 г. | Бромелиевые саламандры | 8 | |
| Эврисия Рафинеск, 1822 г. | Североамериканские ручьевые саламандры | 34 | |
| Гиринофил Коуп, 1869 г. | Весенние саламандры | 4 | |
| Гемидактилий Чуди, 1838 г. | Четырёхпалые саламандры | 1 | |
| Истмура Дюбуа и Рафаэлли, 2012 г. | 7 | ||
| Иксалотритон Уэйк и Джонсон, 1989 г. | Прыгающие саламандры | 2 | |
| Нототритон Уэйк и Элиас, 1983 г. | Моховые саламандры | 22 | |
| Никтанолис Элиас и Уэйк, 1983 г. | Длинноногие саламандры | 1 | |
| Эдипина Кеферштейн, 1868 г. | Червь саламандры | 40 | |
| Парвимолдж Тейлор, 1944 год. | Тропические карликовые саламандры | 1 | |
| Псевдоэврикия Тейлор, 1944 год. | Ложные ручьевые саламандры | 41 | |
| Псевдотритон Чуди, 1838 г. | Грязь и красные саламандры | 2 - 4 | |
| Стереохил Коуп, 1869 г. | Многолинейные саламандры | 1 | |
| Ториус Коуп, 1869 г. | Минутные саламандры | 29 | |
| Лагерь Урспелерпес , Петерман, Миланович, Лэмб, Мерц и Уэйк, 2009 г. | Лоскутные саламандры | 1 | |
| Плетодонтина Грей, 1850 г. | |||
| Анеидес Бэрд, 1851 г. | Восхождение на саламандр | 10 | |
| Десмогнат Бэрд, 1850 г. | Темные саламандры | 39 | |
| Энсатина Грей, 1850 г. | Энсатинас | 1 | |
| Гидромантес Гистель, 1848 г. | Саламандры с паутинными пальцами | 5 | |
| Карсения Мин, Ян, Бонетт, Виетес, Брэндон и Уэйк, 2005 г. | Корейские щелевые саламандры | 1 | |
| Феогнат Хайтон, 1961 г. | Саламандры Красных Холмов | 1 | |
| Плетодон Чуди, 1838 г. | Слизистые и горные саламандры | 58 | |
| Спелеомант Дюбуа, 1984 г. | Европейские пещерные саламандры | 8 |
После серьезного пересмотра в 2006 году род Haideotriton был признан синонимом Eurycea , а род Lineatriton стал синонимом Pseudoeurycea . [24]
Одиночный полупалеоплетодон известен из ископаемых останков миоцена , сохранившихся в доминиканском янтаре , что является единственной находкой саламандр в Карибском бассейне . [25]
Статус сохранения
[ редактировать ]| Статус | Количество видов |
|---|---|
| Наименьшее беспокойство | 94 |
| Рядом с угрозой | 39 |
| Уязвимый | 60 |
| Находящийся под угрозой исчезновения | 88 |
| Находящийся под угрозой исчезновения | 68 |
| Вымерший | 1 |
| Недостаточно данных | 40 |
Ссылки
[ редактировать ]- ^ Jump up to: а б Фрост, Даррел Р. (2024). «Plethodontidae Grey, 1850» . Виды земноводных мира: Интернет-справочник. Версия 6.2 . Американский музей естественной истории. дои : 10.5531/db.vz.0001 . Проверено 4 марта 2024 г.
- ^ Jump up to: а б с «Плетодонтиды» . АмфибияВеб . Калифорнийский университет, Беркли. 2024 . Проверено 4 марта 2024 г.
- ^ Jump up to: а б с Ланца, Б.; Ванни, С.; Нистри, А. (1998). Коггер, Х.Г.; Цвайфель, Р.Г. (ред.). Энциклопедия рептилий и амфибий . Сан-Диего: Академическая пресса. стр. 74–75. ISBN 0-12-178560-2 .
- ^ Льюис, Закари Р.; Керни, Райан; Ханкен, Джеймс (2011). «Развитие легких у безлегочных саламандр!» . Биология развития . 356 (1): 250–251. дои : 10.1016/j.ydbio.2011.05.560 .
- ^ Саламандра Ложного Ручья Белла - World Land Trust
- ^ Хейрстон, Н.Г., старший, 1987. Экология сообщества и гильдии саламандр. Издательство Кембриджского университета. Кембридж.
- ^ «Саламандры — более обильный источник пищи в лесных экосистемах, чем считалось ранее» . ScienceDaily . 18 ноября 2014 г.
- ^ Jump up to: а б Север, Дэвид М. (2003). Репродуктивная биология и филогения Уроделы . Энфилд, Нью-Хэмпшир: Научные издательства. стр. 383–424. ISBN 978-1-57808-645-0 . OCLC 427511083 .
- ^ Север, Дэвид М.; Дастин С. Сигел; Майкл С. Тейлор; Кристофер К. Бичи1 (17 марта 2016 г.). «Возвращение к филогении психических желез» . Копейя . 104 (1): 83–93. дои : 10.1643/CH-14-210 . ПМК 6054469 . ПМИД 30034038 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Дэвид М. Север (18 января 2016 г.). «Ультраструктура подбородочной железы красноспинной саламандры Plethodon cinereus (Amphibia: Plethodontidae)» . Акта Зоология . 98 (2): 154–162. дои : 10.1111/azo.12158 .
- ^ Ариана Э. Рупп; Дэвид М. Север (14 февраля 2017 г.). «Гистология подбородочных и хвостовых ухаживающих желез трех родов плетодонтидных саламандр (амфибии: Plethodontidae)» (PDF) . Акта Зоология . 98 (2): 154–162. дои : 10.1111/azo.12188 . Проверено 5 июля 2022 г.
- ^ Хоук, Линн Д.; Рейган, Нэнси Л. (1990). «Феромоны ухаживания самцов повышают восприимчивость самок у многодонтидной саламандры». Поведение животных . 39 (4): 729–734. дои : 10.1016/s0003-3472(05)80384-7 . S2CID 53185123 .
- ^ Холман, Дж. Алан (2006). Ископаемые саламандры Северной Америки . Издательство Университета Индианы. ISBN 0253347327 .
- ^ Доринг, М. «Саламандры» . gbif.org . Проверено 26 апреля 2024 г.
- ^ Jump up to: а б с Гатц, Рэндалл Н.; Кроуфорд, Юджин К.; Пийпер, Йоханнес (1 февраля 1974 г.). «Дыхательные свойства крови безлегочной и безжаберной саламандры Desmognathus fuscus » . Физиология дыхания . 20 (1): 33–41. дои : 10.1016/0034-5687(74)90016-4 . ПМИД 4821655 .
- ^ Уитфорд, Уолтер Г. (1973). «Влияние температуры на дыхание амфибий». Американский зоолог . 13 (2): 505–512. дои : 10.1093/icb/13.2.505 .
- ^ Бакен, Эрика К.; Меллентин, Лорен Э.; Адамс, Дин С. (2020). «Макроэволюция морфологии, связанной с высыханием, у плетодонтидных саламандр, выведенная на основе нового подхода к оценке соотношения площади поверхности к объему» . Эволюция . 74 (2): 476–486. дои : 10.1111/evo.13898 . ПМИД 31849047 . S2CID 209407983 .
- ^ Jump up to: а б Гатц, Рэндалл Н.; Кроуфорд, Юджин К.; Пийпер, Йоханнес (1 февраля 1974 г.). «Метаболическая реакция и реакция сердечного ритма плетодонтидной саламандры Desmognathus fuscus на гипоксию» . Физиология дыхания . 20 (1): 43–49. дои : 10.1016/0034-5687(74)90017-6 . ПМИД 4821656 .
- ^ Jump up to: а б Льюис, Закари Р.; Керни, Райан; Ханкен, Джеймс (19 августа 2022 г.). «Основы развития эволюционной потери легких у плетодонтидных саламандр» . Достижения науки . 8 (33): eabo6108. Бибкод : 2022SciA....8O6108L . дои : 10.1126/sciadv.abo6108 . ПМЦ 9385146 . ПМИД 35977024 .
- ^ Экспрессия нового гена белка сурфактанта связана с участками внелегочного дыхания у безлегочной саламандры.
- ^ Jump up to: а б Браун, Чарльз Э.; Мартоф, Бернард С. (1 октября 1966 г.). «Функция носогубной борозды плетодонтидных саламандр». Физиологическая зоология . 39 (4): 357–367. дои : 10.1086/physzool.39.4.30152358 . S2CID 87787255 .
- ^ Шен, Син-Син; Лян, Дэн; Чен, Мэн-Юнь; Мао, Ронг-Ли; Уэйк, Дэвид Б.; Чжан, Пэн (январь 2016 г.). «Увеличенный набор мультилокусных данных обеспечивает удивительно более молодое время происхождения Plethodontidae, крупнейшего семейства саламандр» . Систематическая биология . 65 (1): 66–81. дои : 10.1093/sysbio/syv061 . ПМИД 26385618 .
- ^ Санчис, Борха; Венцель, Мартон (2005). «Ископаемая плетодонтидная саламандра из среднего миоцена Словакии (Caudata, Plethodontidae)» . Земноводные-рептилии . 26 (3): 408–411. дои : 10.1163/156853805774408586 .
- ^ Фрост, ДР ; Грант, Т.; Фаивович Ю.Н.; Бэйн, Р.Х.; Хаас, А.; Хаддад, ЦФБ; Де Са, Род-Айленд; Ченнинг, А.; Уилкинсон, М.; Доннеллан, Южная Каролина; Раксуорси, CJ; Кэмпбелл, Дж.А.; Блотто, БЛ; Молер, П.; Древес, Колорадо; Нуссбаум, РА; Линч, доктор медицинских наук; Грин, Д.М. и Уиллер, WC (2006). «Древо жизни земноводных» . Бюллетень Американского музея естественной истории . 297 : 1–291. doi : 10.1206/0003-0090(2006)297[0001:TATOL]2.0.CO;2 . hdl : 2246/5781 .
- ^ Пойнар-младший, Г.; Уэйк, Дэвид Б. (2015). « Paeoplethodon hispaniolae gen. n., sp. n. (Amphibia: Caudata), ископаемая саламандра из Карибского моря» (PDF) . Палеоразнообразие . 8 : 21–29.
- ^ Красный список МСОП, (2020). Плетодонтиды. Получено с https://www.iucnredlist.org/search?taxonomies=101246&searchType=species.
Внешние ссылки
[ редактировать ]Хвостатые семейства по подотряду | ||
|---|---|---|
| ||
| Криптобранхоидеа |  | |
| Саламандроидея | ||
| Сиреноидея | ||
| Базы данных органов управления : Национальные |
|---|