Хищничество


Хищничество — это биологическое взаимодействие , при котором один организм, хищник , убивает и поедает другой организм, свою добычу . Это одно из семейств распространенных форм пищевого поведения , которое включает паразитизм и микрохищничество (которые обычно не убивают хозяина ) и паразитоидизм (который в конечном итоге всегда приводит к гибели). Это отличается от поедания мертвой добычи, хотя многие хищники также питаются ; он частично совпадает с травоядными , поскольку семенные хищники и разрушительные плодоядные животные являются хищниками.
Хищники могут активно искать или преследовать добычу или поджидать ее, часто скрываясь. При обнаружении добычи хищник оценивает, стоит ли нападать на нее. Это может включать засаду или преследование хищников , иногда после преследования добычи. Если атака успешна, хищник убивает добычу, удаляет все несъедобные части, такие как панцирь или шипы, и съедает ее.
Хищники адаптированы и часто узкоспециализированы для охоты, обладая острыми чувствами, такими как зрение , слух или обоняние . Многие хищные животные , как позвоночные , так и беспозвоночные , имеют острые когти или челюсти, позволяющие хватать, убивать и разделывать добычу. Другие адаптации включают скрытность и агрессивную мимику , которые повышают эффективность охоты.
Хищничество оказывает мощное избирательное воздействие на добычу, и у жертвы развиваются антихищнические адаптации, такие как предупреждающая окраска , сигналы тревоги и другие сигналы , маскировка , имитация хорошо защищенных видов, а также защитные шипы и химические вещества. Иногда хищник и жертва оказываются в эволюционной гонке вооружений , цикле адаптаций и контрадаптаций. Хищничество было основным двигателем эволюции, по крайней мере, с кембрийского периода.
Определение [ править ]

На самом базовом уровне хищники убивают и поедают другие организмы. Однако концепция хищничества широка, определяется по-разному в разных контекстах и включает в себя широкий спектр методов питания; более того, некоторые отношения, приводящие к гибели жертвы, не обязательно называются хищничеством. Паразитоид ; , такой как оса-наездник , откладывает яйца в хозяина или на него из яиц вылупляются личинки, которые поедают хозяина, и он неизбежно погибает. Зоологи обычно называют это формой паразитизма , хотя традиционно считается, что паразиты не убивают своих хозяев. Хищника можно определить как отличающегося от паразитоида тем, что у него есть много жертв, пойманных за всю его жизнь, тогда как у личинки паразитоида есть только одна, или, по крайней мере, запас пищи для нее обеспечивается только один раз. [1] [2]

Есть и другие сложные и пограничные случаи. Микрохищники — мелкие животные, которые, как и хищники, полностью питаются другими организмами; к ним относятся блохи и комары , питающиеся кровью живых животных, и тля , питающаяся соком живых растений. Однако, поскольку они обычно не убивают своих хозяев, их теперь часто считают паразитами. [3] [4] Животные, которые пасутся на фитопланктоне или матах микробов, являются хищниками, поскольку они поедают и убивают свои пищевые организмы, в то время как травоядные животные, которые щиплют листья, таковыми не являются, поскольку их пищевые растения обычно переживают нападение. [5] Когда животные поедают семена ( семенное хищничество или зерноядство ) или яйца ( яичное хищничество ), они поедают целые живые организмы, что по определению делает их хищниками. [6] [7] [8]
Падальщики , организмы, которые поедают только уже мертвые организмы, не являются хищниками, но многие хищники, такие как шакал и гиена, собирают мусор, когда появляется возможность. [9] [10] [5] Среди беспозвоночных социальные осы, такие как желтые куртки, являются одновременно охотниками и падальщиками других насекомых. [11]
Таксономический диапазон [ править ]


Хотя примеры хищников среди млекопитающих и птиц хорошо известны, [12] Хищников можно встретить среди самых разных таксонов, включая членистоногих. Они распространены среди насекомых, включая богомолов, стрекоз , златоглазок и скорпионов . У некоторых видов, таких как ольха , хищными являются только личинки (взрослые особи не едят). Пауки являются хищниками, как и другие наземные беспозвоночные, например скорпионы ; многоножки ; некоторые клещи , улитки и слизни ; нематоды ; и планарии . [13] В морской среде большинство книдарий (например, медузы , гидроиды ), гребневиков (гребневиков), иглокожих (например, морские звезды , морские ежи , песчаные доллары и голотурии ) и плоских червей . хищниками являются [14] Среди омары , крабы , ракушки креветки . и являются ракообразных хищниками [15] а на ракообразных, в свою очередь, охотятся почти все головоногие моллюски (включая осьминогов , кальмаров и каракатиц ). [16]

Семенное хищничество характерно только для млекопитающих, птиц и насекомых, но встречается почти во всех наземных экосистемах. [8] [6] Хищничество с яйцами включает в себя как специализированных хищников, таких как некоторые колубридные змеи , так и универсалов, таких как лисы и барсуки, которые оппортунистически забирают яйца, когда находят их. [17] [18] [19]
Некоторые растения, такие как кувшинник , венерина мухоловка и росянка , плотоядны и питаются насекомыми . [12] Методы хищничества растений сильно различаются, но часто включают в себя пищевую ловушку, механическую стимуляцию и электрические импульсы, чтобы в конечном итоге поймать и съесть добычу. [20] Некоторые хищные грибы ловят нематод, используя либо активные ловушки в виде сжимающих колец, либо пассивные ловушки с адгезивными структурами. [21]
Многие виды простейших ( эукариоты ) и бактерий ( прокариоты ) питаются другими микроорганизмами; способ питания, очевидно, древний и неоднократно менялся в обеих группах. [22] [12] [23] Среди пресноводного и морского зоопланктона , как одноклеточного, так и многоклеточного, хищнический поедание фитопланктона распространен и более мелкого зоопланктона, который встречается у многих видов нанофлагеллят , динофлагеллят , инфузорий , коловраток , разнообразного ряда личинок меропланктонных животных и двух групп. ракообразных, а именно веслоногих и ветвистоусых . [24]
Собирательство [ править ]

Чтобы прокормиться, хищник должен искать, преследовать и убивать свою жертву. Эти действия образуют цикл поиска пищи . [26] [27] Хищник должен решить, где искать добычу, исходя из ее географического распределения; и как только он обнаружил добычу, он должен решить, преследовать ее или подождать лучшего выбора. Если он выбирает преследование, его физические возможности определяют способ преследования (например, засада или погоня). [28] [29] Захватив добычу, ему также может потребоваться потратить энергию на ее обработку (например, на ее убийство, удаление панциря или шипов и проглатывание). [25] [26]
Поиск [ править ]
У хищников есть выбор режимов поиска: от сидения и ожидания до активного или широкого поиска пищи . [30] [25] [31] [32] Метод «сиди-жди» наиболее подходит, если добыча плотная и подвижная, а у хищника низкие энергетические потребности. [30] Широкий поиск пищи требует больше энергии и используется, когда добыча малоподвижна или распределена редко. [28] [30] Существует континуум режимов поиска с интервалами между периодами движения от секунд до месяцев. Акулы, рыба-луна , насекомоядные птицы и землеройки почти всегда движутся, тогда как пауки, строящие паутину, водные беспозвоночные, богомолы и пустельги перемещаются редко. Между тем ржанки и другие кулики , пресноводные рыбы, включая краппи , и личинки жуков-кокцинеллид (божьих коровок) попеременно активно ищут и сканируют окружающую среду. [30]

Распределение добычи часто бывает сбивчивым, и хищники в ответ ищут участки, где добыча плотна, а затем ищут их внутри участков. [25] Там, где пища встречается пятнами, например, редкие косяки рыб в почти пустом океане, этап поиска требует от хищника путешествовать в течение значительного времени и затрачивать значительное количество энергии, чтобы найти каждый участок с пищей. [33] Например, чернобровый альбатрос регулярно совершает полеты в поисках пищи на расстояние около 700 километров (430 миль), а максимальная дальность поиска пищи составляет 3000 километров (1860 миль) для размножения птиц, собирающих пищу для своих птенцов. [а] [34] Имея статическую добычу, некоторые хищники могут узнать подходящие места и время от времени возвращаться к ним для кормления. [33] Оптимальная стратегия поиска пищи была смоделирована с использованием теоремы о предельной ценности . [35]
Шаблоны поиска часто кажутся случайными. Одним из таких является ходьба Леви , которая, как правило, включает в себя группы коротких шагов, а иногда и длинные шаги. Он хорошо подходит для поведения самых разных организмов, включая бактерии, медоносных пчел, акул и людей-охотников-собирателей. [36] [37]
Оценка [ править ]

Найдя добычу, хищник должен решить, преследовать ее или продолжать поиск. Решение зависит от связанных с этим затрат и выгод. Птица, добывающая насекомых, тратит много времени на поиски, но поймать и съесть их можно быстро и легко, поэтому эффективная стратегия для птицы — съесть каждое вкусное насекомое, которое она найдет. Напротив, хищник, такой как лев или сокол, легко находит добычу, но для ее поимки требуется много усилий. В этом случае хищник более избирателен. [28]
Одним из факторов, который следует учитывать, является размер. Слишком маленькая добыча может не стоить хлопот из-за количества энергии, которую она дает. Слишком большой, и его может быть слишком сложно поймать. Например, богомол захватывает добычу передними ногами, и они оптимизированы для захвата добычи определенного размера. Богомолы неохотно нападают на добычу, далекую от такого размера. Существует положительная корреляция между размером хищника и его жертвой. [28]
Хищник также может оценить участок и решить, стоит ли тратить на него время на поиск добычи. [25] Это может потребовать некоторого знания предпочтений жертвы; например, божьи коровки могут выбрать участок растительности, подходящий для их добычи тли . [38]
Захватить [ править ]
Чтобы поймать добычу, у хищников есть целый спектр режимов преследования, которые варьируются от явной погони ( хищничество преследования ) до внезапного нападения на ближайшую добычу ( хищничество из засады ). [25] [39] [12] Другая стратегия, промежуточная между засадой и преследованием, — это баллистический перехват , когда хищник наблюдает и предсказывает движение жертвы, а затем соответствующим образом начинает атаку. [40]
Засада [ править ]


Хищники, устраивающие засаду или сидящие и ждущие, - это плотоядные животные, которые ловят добычу незаметно или неожиданно. У животных хищничество из засады характеризуется тем, что хищник сканирует окружающую среду из скрытого положения до тех пор, пока не будет обнаружена жертва, а затем быстро совершает фиксированную внезапную атаку. [41] [40] К позвоночным хищникам, устраивающим засады, относятся лягушки, такие рыбы, как акула-ангел , северная щука и восточная рыба-лягушка . [40] [42] [43] [44] Среди многих беспозвоночных хищников, устраивающих засады, есть пауки-люки и австралийские пауки-крабы на суше, а также креветки-богомолы в море. [41] [45] [46] Хищники, устраивающие засаду, часто строят норы, в которых можно спрятаться, улучшая маскировку за счет уменьшения поля зрения. Некоторые хищники, устраивающие засады, также используют приманки, чтобы привлечь добычу на расстояние удара. [40] Чтобы поймать добычу, движение захвата должно быть быстрым, поскольку после начала атаки изменить ее невозможно. [40]
Баллистический перехват [ править ]
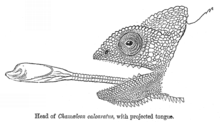
Баллистический перехват — это стратегия, при которой хищник наблюдает за движением добычи, прогнозирует ее движение, определяет путь перехвата, а затем атакует добычу на этом пути. Это отличается от хищничества из засады тем, что хищник корректирует свою атаку в зависимости от того, как движется добыча. [40] Баллистический перехват предполагает короткий период планирования, дающий жертве возможность сбежать. Некоторые лягушки ждут, пока змеи начнут атаковать, прежде чем прыгнуть, сокращая время, доступное змее для повторной калибровки атаки, и максимизируя угловую регулировку, которую змее необходимо будет сделать, чтобы перехватить лягушку в реальном времени. [40] К баллистическим хищникам относятся насекомые, такие как стрекозы, и позвоночные, такие как рыбы-стрелки (атакующие струей воды), хамелеоны (атакующие языком) и некоторые змеи-колобриды . [40]
Преследование [ править ]


Преследуя хищников, хищники преследуют убегающую добычу. Если жертва бежит по прямой, поимка зависит только от того, будет ли хищник быстрее добычи. [40] Если добыча маневрирует, поворачиваясь при бегстве, хищник должен реагировать в реальном времени, чтобы рассчитать и следовать по новому пути перехвата, например, с помощью параллельной навигации , когда он приближается к добыче. [40] Многие хищники-преследователи используют камуфляж, чтобы приблизиться к жертве как можно ближе незамеченным ( преследование ), прежде чем начать преследование. [40] Хищники-преследователи включают наземных млекопитающих, таких как люди, африканские дикие собаки, пятнистые гиены и волки; морские хищники, такие как дельфины, косатки и многие хищные рыбы, такие как тунец; [47] [48] хищные птицы (рапторы), например соколы; и насекомые, такие как стрекозы . [49]
Крайней формой преследования является охота на выносливость или настойчивость , при которой хищник утомляет добычу, преследуя ее на большом расстоянии, иногда в течение нескольких часов. Этот метод используется людьми -охотниками-собирателями , а также псовыми, такими как африканские дикие собаки и домашние гончие. Африканская дикая собака — чрезвычайно настойчивый хищник, утомляющий отдельную добычу, преследуя ее на протяжении многих миль с относительно низкой скоростью. [50]
Специализированной формой преследования хищников является кормление с усатых китов выпадом . Эти очень крупные морские хищники питаются планктоном , особенно крилем , ныряя и активно плавая в скоплениях планктона, а затем заглатывая воду огромным глотком и фильтруя ее через свои перистые пластинки уса . [51] [52]
Хищники-преследователи могут быть социальными , как лев и волк, которые охотятся группами, или одиночными. [2]
Обработка [ править ]


После того, как хищник поймал добычу, он должен обращаться с ней: очень осторожно, если добыча опасна для употребления в пищу, например, если она обладает острыми или ядовитыми шипами, как у многих хищных рыб. У некоторых сомов, таких как Ictaluridae, есть шипы на спине (спинной части) и брюхе (грудной), которые фиксируются в вертикальном положении; поскольку сом мечется во время поимки, они могут пронзить пасть хищника, возможно, смертельно. Некоторые рыбоядные птицы, такие как скопа, избегают опасности шипов, разрывая добычу перед тем, как ее съесть. [53]
хищничество Одиночное и социальное
При социальном хищничестве группа хищников сотрудничает, чтобы убить добычу. Это позволяет убивать существ более крупных, чем те, которых они могли бы одолеть в одиночку; например, гиены и волки сотрудничают, чтобы поймать и убить травоядных животных размером с буйвола, а львы даже охотятся на слонов. [54] [55] [56] Он также может сделать добычу более доступной с помощью таких стратегий, как смыв добычи и перегонка ее на меньшую территорию. Например, когда кормятся смешанные стаи птиц, птицы впереди спугивают насекомых, которых ловят птицы сзади. Дельфины-спиннеры образуют круг вокруг косяка рыбы и движутся внутрь, концентрируя рыбу в 200 раз. [57] Охотясь социально, шимпанзе могут поймать обезьян-колобусов , которые легко ускользнут от отдельного охотника, а сотрудничающие с ними ястребы Харриса могут поймать кроликов. [54] [58]

Хищники разных видов иногда сотрудничают, чтобы поймать добычу. На коралловых рифах , когда такие рыбы, как морской окунь и коралловая форель, замечают недоступную для них добычу, они подают сигнал гигантским муренам , губанам-наполеонам или осьминогам . Эти хищники способны проникать в небольшие щели и выманивать добычу. [59] [60] усатых Известно, что косатки помогают китобоям охотиться на китов . [61]
Социальная охота позволяет хищникам ловить более широкий круг добычи, но при этом существует риск конкуренции за добытую пищу. У одиночных хищников больше шансов съесть то, что они поймают, за счет увеличения затрат энергии на поимку и повышенного риска того, что добыча сбежит. [62] [63] Хищники, устраивающие засаду, часто ведут одиночный образ жизни, чтобы снизить риск самим стать добычей. [64] Из 245 наземных представителей хищных ( группа, в которую входят кошки, собаки и медведи) 177 — одиночки; и 35 из 37 диких кошек ведут одиночный образ жизни. [65] включая пуму и гепарда. [62] [2] Тем не менее, одинокая пума позволяет другим пумам участвовать в добыче. [66] а койот может быть как одиночным, так и социальным. [67] Другие хищники-одиночки включают северную щуку, [68] пауки-волки и все тысячи видов одиночных ос среди членистоногих, [69] [70] и многие микроорганизмы и зоопланктон . [22] [71]
Специализация [ править ]
Физические адаптации [ править ]
Под давлением естественного отбора хищники развили множество физических приспособлений для обнаружения, ловли, убийства и переваривания добычи. К ним относятся скорость, ловкость, скрытность, острые чувства, когти, зубы, фильтры и подходящая пищеварительная система. [72]
Для обнаружения добычи у хищников хорошо развито зрение , обоняние или слух . [12] Такие разнообразные хищники, как совы и пауки-прыгуны , имеют глаза, обращенные вперед, что обеспечивает точное бинокулярное зрение в относительно узком поле зрения, тогда как животные-жертвы часто имеют менее острое круговое зрение. Такие животные, как лисы, чувствуют запах своей добычи, даже если она спрятана под снегом или землей на глубине 2 футов (60 см). Многие хищники обладают острым слухом, а некоторые, например -эхолокаторы летучие мыши , охотятся исключительно за счет активного или пассивного использования звука. [73]
Хищники, в том числе большие кошки , хищные птицы и муравьи, имеют общие мощные челюсти, острые зубы или когти, которые они используют, чтобы схватить и убить свою добычу. Некоторые хищники, такие как змеи и рыбоядные птицы, такие как цапли и бакланы, заглатывают добычу целиком; некоторые змеи могут раздвигать челюсти, чтобы проглотить крупную добычу, в то время как у рыбоядных птиц есть длинные копьевидные клювы, которые они используют, чтобы наносить удары и хватать быстро движущуюся и скользкую добычу. [73] Рыбы и другие хищники развили способность раздавливать или вскрывать панцирные раковины моллюсков. [74]
Многие хищники имеют мощное телосложение и могут ловить и убивать животных крупнее их самих; это относится как к мелким хищникам, таким как муравьи и землеройки, так и к большим и заметно мускулистым хищникам, таким как пума и лев . [73] [2] [75]
 Череп бурого медведя имеет большие заостренные клыки, предназначенные для убийства добычи, и самозатачивающиеся плотоядные зубы сзади, позволяющие разрезать плоть, подобно ножницам.
Череп бурого медведя имеет большие заостренные клыки, предназначенные для убийства добычи, и самозатачивающиеся плотоядные зубы сзади, позволяющие разрезать плоть, подобно ножницам.

 Краснохвостый ястреб использует острые крючковатые когти и клюв, чтобы убивать и разрывать добычу.
Краснохвостый ястреб использует острые крючковатые когти и клюв, чтобы убивать и разрывать добычу. Специалист: большая голубая цапля с копьем.
Специалист: большая голубая цапля с копьем. Индийский питон раздвигает челюсти, чтобы проглотить такую крупную добычу, как этот читал
Индийский питон раздвигает челюсти, чтобы проглотить такую крупную добычу, как этот читал






Диета поведение и


Хищники часто узкоспециализированы в своем питании и охотничьем поведении; например, евразийская рысь охотится только на мелких копытных . [76] Другие, такие как леопарды, являются более оппортунистическими универсалами, охотящимися как минимум на 100 видов. [77] [78] Специалисты могут быть хорошо приспособлены к поимке предпочитаемой ими добычи, тогда как универсалы могут лучше переключиться на другую добычу, когда предпочтительной цели не хватает. Когда добыча распределена сгруппировано (неравномерно), оптимальная стратегия для хищника, по прогнозам, будет более специализированной, поскольку добыча более заметна и ее можно найти быстрее; [79] это кажется правильным для хищников, питающихся неподвижной добычей, но сомнительно для подвижной добычи. [80]
При хищничестве, избирательном по размеру, хищники выбирают добычу определенного размера. [81] Крупная добыча может оказаться неприятной для хищника, тогда как мелкую добычу может оказаться трудно найти, и в любом случае она принесет меньшую награду. Это привело к корреляции между размером хищников и их добычей. Размер также может служить убежищем для крупной добычи. Например, взрослые слоны относительно защищены от нападения львов, но молодые особи уязвимы. [82]
Камуфляж и мимикрия [ править ]


Представители семейства кошачьих , такие как снежный барс (безлесные высокогорья), тигр (травянистые равнины, тростниковые болота), оцелот (лес), кошка-рыболов (прибрежные заросли) и лев (открытые равнины), замаскированы с помощью окраски и разрушительных узоров, подходящих для места их обитания. [83]
При агрессивной мимикрии некоторые хищники, в том числе насекомые и рыбы, используют окраску и поведение для привлечения добычи. Самки Photuris светлячков , например, копируют световые сигналы других видов, тем самым привлекая самцов светлячков, которых они ловят и поедают. [84] Цветочные богомолы - хищники, устраивающие засаду; маскируясь под цветы, например орхидеи , они привлекают добычу и хватают ее, когда она оказывается достаточно близко. [85] Рыбы-лягушки чрезвычайно хорошо маскируются и активно приманивают свою добычу, используя эску , приманку на конце стержнеобразного придатка на голове, которой они осторожно машут, имитируя маленького животного, заглатывая добычу чрезвычайно быстрыми движениями. когда он находится в пределах досягаемости. [86]
Веном [ править ]
Многие более мелкие хищники, такие как коробчатая медуза, используют яд , чтобы усмирить свою добычу. [87] а яд также может способствовать пищеварению (как в случае гремучих змей и некоторых пауков ). [88] [89] У мраморной морской змеи, приспособившейся к поеданию яиц, атрофированы ядовитые железы, а ген ее трехпальцевого токсина содержит мутацию (удаление двух нуклеотидов ), которая инактивирует ее. Эти изменения объясняются тем, что его добычу не нужно подчинять себе. [90]
Электрические поля [ править ]
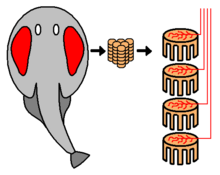
Некоторые группы хищных рыб обладают способностью обнаруживать, отслеживать, а иногда, как в случае с электрическим скатом , выводить из строя свою жертву, ощущая и генерируя электрические поля . [91] [92] [93] Электрический орган получен из модифицированной нервной или мышечной ткани. [94]
Физиология [ править ]
Физиологическая адаптация к хищничеству включает способность хищных бактерий переваривать сложный полимер пептидогликана из клеточных стенок бактерий, на которых они охотятся. [23] Плотоядные позвоночные всех пяти основных классов (рыбы, амфибии, рептилии, птицы и млекопитающие) имеют более низкую относительную скорость транспорта сахара к аминокислотам , чем травоядные или всеядные, предположительно потому, что они получают большое количество аминокислот из животных белков в своем рационе. . [95]
Адаптации против хищников


Чтобы противостоять хищникам, жертва выработала защитные механизмы, которые можно использовать на каждом этапе атаки. [96] [12] Они могут попытаться избежать обнаружения, [97] например, с помощью камуфляжа и мимикрии . [98] Они могут обнаружить хищников [99] и предупреждать других о своем присутствии. [100] [101] В случае обнаружения они могут попытаться избежать нападения, например, сигнализируя о том, что они токсичны или неприятны на вкус . [102] [103] [104] сигнализируя, что погоня будет невыгодной, [105] [106] или путем формирования групп. [107] [108] Если они становятся целью, они могут попытаться отразить атаку с помощью таких средств защиты, как броня, иглы , неприятности или толпы; [109] [110] [111] и они часто могут избежать нападения, напугав хищника, [112] [113] [114] притворяясь мертвым , теряя части тела, такие как хвосты, или просто убегая. [115] [116]
Коэволюция [ править ]

Хищники и жертвы — естественные враги, и многие из их приспособлений, похоже, созданы для противодействия друг другу. Например, летучие мыши обладают сложной системой эхолокации для обнаружения насекомых и другой добычи, а насекомые выработали множество защитных механизмов, включая способность слышать эхолокационные сигналы. [117] [118] Многие хищники-преследователи, бегающие по суше, например волки, развили длинные конечности в ответ на увеличение скорости своей добычи. [119] Их адаптации были охарактеризованы как эволюционная гонка вооружений , пример совместной эволюции двух видов. [120] С геноцентрированной точки зрения на эволюцию гены хищника и жертвы можно рассматривать как конкурирующих за тело жертвы. [120] Однако принцип «жизни-ужина» Докинза и Кребса предсказывает, что эта гонка вооружений асимметрична: если хищнику не удается поймать добычу, он теряет обед, а если ему это удается, жертва теряет жизнь. [120]

Метафора гонки вооружений подразумевает постоянно растущие успехи в нападении и обороне. Однако за эти адаптации приходится платить; например, более длинные ноги имеют повышенный риск перелома, [121] тогда как специализированный язык хамелеона, обладающий способностью действовать как снаряд, бесполезен для лакания воды, поэтому хамелеону приходится пить росу с растительности. [122]
Принцип «жизни-ужина» подвергался критике по многим причинам. Степень асимметрии естественного отбора частично зависит от наследуемости адаптивных признаков. [122] Кроме того, если хищник потеряет достаточное количество обедов, он тоже потеряет свою жизнь. [121] [122] С другой стороны, стоимость потерянного ужина непредсказуема, поскольку хищник может быстро найти лучшую добычу. Кроме того, большинство хищников являются универсалами, что снижает влияние конкретной адаптации жертвы на хищника. Поскольку специализация вызвана коэволюцией хищник-жертва, редкость специалистов может означать, что гонка вооружений хищник-жертва редка. [122]
Трудно определить, действительно ли данные адаптации являются результатом совместной эволюции, когда адаптация жертвы приводит к адаптации хищника, которой противостоит дальнейшая адаптация жертвы. Альтернативное объяснение — эскалация , когда хищники приспосабливаются к конкурентам, своим собственным хищникам или опасной добыче. [123] Очевидная адаптация к хищничеству могла возникнуть и по другим причинам, а затем быть использована для нападения или защиты. У некоторых насекомых, на которых охотятся летучие мыши, слух развился до появления летучих мышей и использовался для восприятия сигналов, используемых для территориальной защиты и спаривания. [124] Их слух развился в ответ на хищничество летучих мышей, но единственным ярким примером взаимной адаптации у летучих мышей является скрытая эхолокация. [125]
Более симметричная гонка вооружений может возникнуть, когда жертва опасна, поскольку у нее есть шипы, иглы, токсины или яд, которые могут нанести вред хищнику. Хищник может реагировать уклонением, что, в свою очередь, стимулирует эволюцию мимикрии. Избегание не обязательно является эволюционной реакцией, поскольку оно обычно вырабатывается на основе неудачного опыта общения с добычей. Однако, когда добыча способна убить хищника (как это может сделать коралловая змея своим ядом), возможности для обучения нет, и избегание должно передаваться по наследству. Хищники также могут реагировать на опасную добычу контрадаптациями. В западной части Северной Америки обыкновенная подвязочная змея выработала устойчивость к токсину кожи грубокожего тритона . [122]
в экосистемах Роль
Хищники влияют на свои экосистемы не только напрямую, поедая свою собственную добычу, но и косвенными способами, такими как сокращение хищничества со стороны других видов или изменение пищевого поведения травоядных, как в случае с влиянием на биоразнообразие волков на прибрежную растительность или каланов на леса из водорослей. . Это может объяснить эффекты динамики популяций, такие как циклы, наблюдаемые у рыси и зайцев-снегоступов. [126] [127] [128]
Трофический уровень [ править ]
Один из способов классификации хищников — по трофическому уровню . Хищники , питающиеся травоядными, являются вторичными потребителями; их хищники являются третичными потребителями и так далее. [129] На вершине этой пищевой цепи находятся высшие хищники, такие как львы . [130] Однако многие хищники питаются на нескольких уровнях пищевой цепи; Хищник может поедать как вторичных, так и третичных потребителей. [131] Это означает, что многим хищникам приходится бороться с внутригильдейским хищничеством , когда другие хищники убивают и поедают их. Например, койоты конкурируют с серыми лисами и рыси , а иногда и убивают их . [132]
Трофический перенос [ править ]
Трофический перенос внутри экосистемы относится к транспортировке энергии и питательных веществ в результате хищничества. Энергия переходит с одного трофического уровня на другой по мере того, как хищники потребляют органическое вещество из тела другого организма . В рамках каждой передачи наряду с использованием энергии существуют и ее потери.
Уровни морской трофики варьируются в зависимости от местности и размера первичных производителей . Обычно существует до шести трофических уровней в открытом океане, четыре над континентальными шельфами и около трех в зонах апвеллинга. [133] Например, морскую среду обитания с пятью трофическими уровнями можно представить следующим образом: травоядные (питаются преимущественно фитопланктоном ); Хищники (питаются в основном другим зоопланктоном /животными); Детритофаги (питаются в основном мертвым органическим веществом/ детритом ; всеядные (питаются смешанной диетой, состоящей из фито- и зоопланктона и детрита); и миксотрофы , которые сочетают автотрофность (использование энергии света для роста без потребления каких-либо дополнительных органических соединений или питательных веществ) с гетеротрофностью . (питание другими растениями и животными для получения энергии и питательных веществ — травоядные, всеядные, плотоядные и детритофаги).
Эффективность трофической передачи измеряет, насколько эффективно энергия передается или проходит через более высокие трофические уровни морской пищевой сети . По мере продвижения энергии вверх по трофическим уровням она уменьшается из-за тепла, отходов и естественных метаболических процессов , которые происходят, когда хищники поедают свою добычу. В результате только около 10% энергии на любом трофическом уровне передается на следующий уровень. Это часто называют «правилом 10%», которое ограничивает количество трофических уровней, которые способна поддерживать отдельная экосистема. [134]
хищников за счет поддерживается Биоразнообразие
Хищники могут увеличить биоразнообразие сообществ, предотвращая доминирование одного вида. Такие хищники известны как ключевые виды и могут оказывать глубокое влияние на баланс организмов в конкретной экосистеме . [135] Интродукция или удаление этого хищника, а также изменение плотности его популяции могут иметь радикальные каскадные последствия для равновесия многих других популяций в экосистеме. Например, пасущиеся на пастбищах могут помешать одному доминирующему виду захватить власть. [136]

Искоренение волков в Йеллоустонском национальном парке оказало глубокое воздействие на трофическую пирамиду . В этой области волки являются одновременно ключевыми видами и высшими хищниками. Без хищников травоядные животные начали чрезмерно выпасать многие виды древесных побегов, что повлияло на популяции растений в этом районе. Кроме того, волки часто не позволяли животным пастись возле ручьев, защищая источники пищи бобров . Удаление волков оказало прямое влияние на популяцию бобров, поскольку их среда обитания стала территорией для выпаса скота. Увеличение посещений ив и хвойных пород вдоль ручья Блэктейл из-за отсутствия хищников привело к разрыву канала, поскольку сократившаяся популяция бобров больше не могла замедлять течение воды и удерживать почву на месте. Таким образом, было продемонстрировано, что хищники имеют жизненно важное значение в экосистеме. [137]
населения Динамика
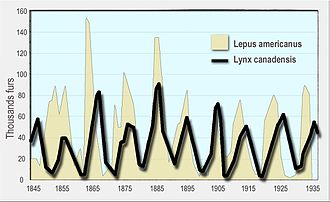
В отсутствие хищников популяция вида может расти в геометрической прогрессии, пока не достигнет несущей способности окружающей среды. [138] Хищники ограничивают рост добычи, как поедая ее, так и изменяя ее поведение. [139] Увеличение или уменьшение популяции жертв также может привести к увеличению или уменьшению численности хищников, например, за счет увеличения количества детенышей, которых они вынашивают.
В популяциях хищников и жертв наблюдались циклические колебания, часто со смещением между циклами хищника и добычи. Хорошо известный пример — заяц -беляк и рысь . На обширной территории бореальных лесов Аляски и Канады популяция зайцев колеблется почти синхронно с 10-летним периодом, а популяция рыси колеблется в ответ на это. Впервые это было замечено в исторических записях о животных, пойманных охотниками за мехом для компании Гудзонова залива за более чем столетие. [140] [128] [141] [142]

Простая модель системы с одним видом хищников и жертв, уравнения Лотки-Вольтерра , предсказывает популяционные циклы. [143] Однако попытки воспроизвести предсказания этой модели в лаборатории часто терпели неудачу; например, когда простейшее Didinium nasutum добавляется к культуре, содержащей его добычу, Paramecium caudatum , последняя часто оказывается на грани исчезновения. [144]
Уравнения Лотки-Вольтерра основаны на нескольких упрощающих предположениях, и они структурно нестабильны , а это означает, что любое изменение в уравнениях может стабилизировать или дестабилизировать динамику. [145] [146] Например, одно из предположений состоит в том, что у хищников есть линейная функциональная реакция на добычу: скорость убийств увеличивается пропорционально частоте встреч. Если эта скорость ограничена временем, затрачиваемым на обработку каждого улова, то плотность популяции добычи может достичь такой плотности, при которой хищники не смогут ее контролировать. [144] Другое предположение состоит в том, что все жертвы идентичны. На самом деле хищники, как правило, выбирают молодых, слабых и больных особей, оставляя популяцию добычи способной вырасти заново. [147]
Многие факторы могут стабилизировать популяции хищников и жертв. [148] Одним из примеров является наличие нескольких хищников, особенно универсалов, которых привлекает определенный вид добычи, если его много, и которые ищут в другом месте, если его нет. [149] В результате популяционные циклы, как правило, наблюдаются в северных умеренных и субарктических экосистемах, поскольку пищевые сети проще. [150] Система зайцев-рысей на снегоступах является субарктической, но даже в ней участвуют и другие хищники, в том числе койоты, ястребы-тетеревятники и большие рогатые совы , а цикл подкрепляется вариациями в пище, доступной зайцам. [151]
Ряд математических моделей был разработан путем смягчения допущений, сделанных в модели Лотки – Вольтерра; они по-разному позволяют животным иметь географическое распространение или мигрировать ; иметь различия между особями, такие как пол и возрастная структура , так что только некоторые особи размножаются; жить в меняющейся среде, например, со сменой времен года ; [152] [153] и анализ взаимодействия более чем двух видов одновременно. Такие модели предсказывают сильно различающуюся и часто хаотичную динамику популяций хищник-жертва. [152] [154] Наличие убежищ , где добыча защищена от хищников, может позволить добыче поддерживать более крупные популяции, но также может дестабилизировать динамику. [155] [156] [157] [158]
Эволюционная история [ править ]
Хищничество возникло задолго до появления общепризнанных хищников на сотни миллионов (возможно, миллиарды) лет. Хищничество неоднократно развивалось у разных групп организмов. [5] [159] Возникновение эукариотических клеток около 2,7 Гя, появление многоклеточных организмов около 2 Гя и появление мобильных хищников (около 600 млн лет назад - 2 Гя, вероятно, около 1 Гя) - все это приписывают раннему хищническому поведению, и многие На очень ранних останках имеются следы скважин или других отметин, приписываемых мелким видам хищников. [5] Вероятно, это вызвало серьезные эволюционные переходы, включая появление клеток , эукариот , половое размножение , многоклеточность , увеличение размера, подвижность (включая полет насекомых) . [160] ) и бронеснаряды и экзоскелеты. [5]
Самыми ранними хищниками были микробные организмы, которые поглощали других или паслись на них. Поскольку летопись окаменелостей скудна, возраст этих первых хищников мог составлять от 1 до 2,7 млрд лет назад (миллиарда лет назад). [5] Хищничество, очевидно, приобрело важное значение незадолго до кембрийского периода — около 550 миллионов лет назад — о чем свидетельствует почти одновременное развитие кальцификации у животных и водорослей. [161] и зарывание нор, избегающее хищников . Однако хищники питались микроорганизмами по крайней мере 1000 миллионов лет назад . [5] [162] [163] со свидетельствами избирательного (а не случайного) хищничества в то же время. [164]
Auroralumina attenboroughii эдиакарской коронной группы — книдарий (557–562 млн лет назад, примерно за 20 миллионов лет до кембрийского взрыва) из Чарнвудского леса , Англия. Считается, что это одно из самых ранних хищных животных, ловящее мелкую добычу своими нематоцистами , как это делают современные книдарии. [165]
Летопись окаменелостей демонстрирует долгую историю взаимодействия между хищниками и их добычей, начиная с кембрийского периода, показывая, например, что некоторые хищники просверливали раковины двустворчатых и брюхоногих моллюсков, в то время как другие поедали эти организмы, разбивая их раковины. [166] Среди кембрийских хищников были такие беспозвоночные, как аномалокаридиды , с придатками, пригодными для хватания добычи, большими сложными глазами и челюстями из твердого материала, подобного экзоскелету насекомого . [167] первых рыб, у которых были челюсти, были бронированные и в основном хищные плакодермы силурийского Одними из и девонского периодов, одна из которых, 6-метровый (20 футов) Dunkleosteus , считается первым в мире позвоночным «сверххищником», охотящимся на других хищников. [168] [169] Насекомые развили способность летать в раннем карбоне или позднем девоне, что позволило им, среди прочего, спасаться от хищников. [160] Среди крупнейших хищников, которые когда-либо жили, были динозавры-тероподы, такие как тираннозавр из мелового периода. Они охотились на травоядных динозавров, таких как гадрозавры , цератопсы и анкилозавры . [170]
![Auroralumina attenboroughii, эдиакарский хищник (около 560 млн лет назад). Это был книдарий стволовой группы, ловивший добычу нематоцистами.[165]](//upload.wikimedia.org/wikipedia/commons/thumb/6/63/Auroralumina_attenboroughii_reconstruction.jpg/157px-Auroralumina_attenboroughii_reconstruction.jpg) Auroralumina attenboroughii , эдиакарский хищник (ок. 560 млн лет назад). стеблевой группы Это был книдарий , ловивший добычу своими нематоцистами . [165]
Auroralumina attenboroughii , эдиакарский хищник (ок. 560 млн лет назад). стеблевой группы Это был книдарий , ловивший добычу своими нематоцистами . [165] Кембрийская революция субстрата привела к тому, что жизнь на морском дне изменилась от минимального роющего (слева) к разнообразной роющей фауне (справа), вероятно, чтобы избежать новых кембрийских хищников.
Кембрийская революция субстрата привела к тому, что жизнь на морском дне изменилась от минимального роющего (слева) к разнообразной роющей фауне (справа), вероятно, чтобы избежать новых кембрийских хищников.

 Meganeura moNYi , хищное каменноугольного периода насекомое , родственное стрекозам , могло летать, спасаясь от наземных хищников. Его большой размер с размахом крыльев 65 см (30 дюймов) может отражать отсутствие в то время позвоночных воздушных хищников.
Meganeura moNYi , хищное каменноугольного периода насекомое , родственное стрекозам , могло летать, спасаясь от наземных хищников. Его большой размер с размахом крыльев 65 см (30 дюймов) может отражать отсутствие в то время позвоночных воздушных хищников.
![Auroralumina attenboroughii, эдиакарский хищник (около 560 млн лет назад). Это был книдарий стволовой группы, ловивший добычу нематоцистами.[165]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/63/Auroralumina_attenboroughii_reconstruction.jpg/157px-Auroralumina_attenboroughii_reconstruction.jpg)





В человеческом обществе [ править ]

Практическое использование
Человек, будучи всеядным , в некоторой степени хищен. [171] использование оружия и инструментов для ловли рыбы , [172] охотиться и ловить животных. [173] Они также используют другие хищные виды, такие как собаки , бакланы , [174] и соколы, чтобы ловить добычу для еды или для развлечения. [175] Два хищника среднего размера, собаки и кошки, — животные, которых чаще всего держат в качестве домашних животных в западном обществе. [176] [177] Человеческие охотники, в том числе сан из южной Африки, используют настойчивую охоту , форму преследования хищников, при которой преследователь может быть медленнее добычи, такой как антилопа куду, на коротких дистанциях, но преследует ее в полуденную жару, пока она не утомится, преследование. это может занять до пяти часов. [178] [179]
При биологической борьбе с вредителями хищники (и паразитоиды) из естественного ареала вредителя вводятся для контроля популяций, рискуя вызвать непредвиденные проблемы. Естественные хищники, при условии, что они не причиняют вреда видам, не являющимся вредителями, являются экологически чистым и устойчивым способом уменьшения ущерба сельскохозяйственным культурам и альтернативой использованию химических веществ, таких как пестициды . [180]
Символическое использование [ править ]

В кино идея хищника как опасного, хотя и гуманоидного врага используется в научно-фантастическом ужасов боевике 1987 года «Хищник» и трех его продолжениях . [181] [182] Ужасающий хищник, гигантская -людоед большая белая акула , также занимает центральное место в триллере Стивена Спилберга 1974 года «Челюсти» . [183]
Среди поэзии на тему хищничества можно было бы исследовать сознание хищника, как, например, в » Теда Хьюза «Пайке . [184] Фраза «Природа, красные зубы и когти» из стихотворения Альфреда Лорда Теннисона 1849 года « In Memoriam AHH » была истолкована как относящаяся к борьбе между хищниками и добычей. [185]
В мифологии и народных баснях хищники, такие как лиса и волк, имеют неоднозначную репутацию. [186] Лиса была символом плодородия в древней Греции, погодным демоном в Северной Европе и порождением дьявола в раннем христианстве; , лиса представлена хитрой, жадной и хитрой В баснях, начиная с Эзопа . [186] Большой злой волк известен детям в таких сказках, как «Красная Шапочка» , но является демонической фигурой в исландских сагах «Эдда» , где волк Фенрир появляется в апокалиптическом конце света . [186] В Средние века распространилась вера в оборотней , мужчин, превращавшихся в волков. [186] В Древнем Риме и Древнем Египте волку поклонялись, волчица фигурирует в мифе об основании Рима и кормит грудью Ромула и Рема . [186] Совсем недавно, в Редьярда Киплинга 1894 года «Книге джунглей» , Маугли воспитывается волчьей стаей. [186] Отношение к крупным хищникам Северной Америки, таким как волк, медведь гризли и пума, во второй половине XX века сместилось от враждебного или двойственного, сопровождавшегося активным преследованием, к позитивному и покровительственному. [187]
См. также [ править ]
Примечания [ править ]
- ^ Дальность 3000 километров означает дальность полета не менее 6000 километров туда и обратно.
Ссылки [ править ]
- ^ Гурр, Джефф М.; Раттен, Стивен Д.; Снайдер, Уильям Э. (2012). Биоразнообразие и насекомые-вредители: ключевые проблемы устойчивого управления . Джон Уайли и сыновья. п. 105. ИСБН 978-1-118-23185-2 .
- ^ Jump up to: Перейти обратно: а б с д Лафферти, К.Д.; Курис, AM (2002). «Трофические стратегии, разнообразие животных и размеры тела». Тенденции Экол. Эвол . 17 (11): 507–513. дои : 10.1016/s0169-5347(02)02615-0 .
- ^ Пулен, Робер ; Рандхава, Хасиб С. (февраль 2015 г.). «Эволюция паразитизма по конвергентным направлениям: от экологии к геномике» . Паразитология . 142 (Приложение 1): С6–С15. дои : 10.1017/S0031182013001674 . ПМЦ 4413784 . ПМИД 24229807 .
- ^ Пулен, Роберт (2011). Роллинсон, Д.; Хэй, С.И. (ред.). «Множество дорог к паразитизму: история конвергенции» . Достижения паразитологии . 74 . Академическая пресса: 27–28. дои : 10.1016/B978-0-12-385897-9.00001-X . ISBN 978-0-12-385897-9 . ПМИД 21295676 .
- ^ Jump up to: Перейти обратно: а б с д и ж г Бенгтсон, С. (2002). «Происхождение и ранняя эволюция хищничества». В Ковалевском, М.; Келли, PH (ред.). Ископаемая летопись хищничества. Документы Палеонтологического общества 8 (PDF) . Палеонтологическое общество. стр. 289–317.
- ^ Jump up to: Перейти обратно: а б Янзен, Д.Х. (1971). «Похищение семян животными». Ежегодный обзор экологии и систематики . 2 : 465–492. doi : 10.1146/annurev.es.02.110171.002341 .
- ^ Нильссон, Свен Г.; Бьоркман, Кристер; Форслунд, Пяр; Хёглунд, Джейкоб (1985). «Хищничество яиц в сообществах лесных птиц на островах и материке». Экология . 66 (4): 511–515. Бибкод : 1985Oecol..66..511N . дои : 10.1007/BF00379342 . ПМИД 28310791 . S2CID 2145031 .
- ^ Jump up to: Перейти обратно: а б Халм, ЧП; Бенкман, CW (2002). «Граниворы». В CM Herrera и O. Pellmyr (ред.). Взаимодействие растений и животных: эволюционный подход . Блэквелл. стр. 132–154. ISBN 978-0-632-05267-7 .
- ^ Кейн, Адам; Хили, Кевин; Гильерме, Томас; Ракстон, Грэм Д.; Джексон, Эндрю Л. (2017). «Рецепт уборки мусора у позвоночных - естественная история поведения». Экография . 40 (2): 324–334. Бибкод : 2017Экогр..40..324К . дои : 10.1111/ecog.02817 . hdl : 10468/3213 . S2CID 56280901 .
- ^ Круук, Ганс (1972). Пятнистая гиена: исследование хищничества и социального поведения . Издательство Калифорнийского университета. стр. 107–108. ISBN 978-0226455082 .
- ^ Шмидт, Джастин О. (2009). «Осы». Осы - ScienceDirect (Второе изд.). стр. 1049–1052. дои : 10.1016/B978-0-12-374144-8.00275-7 . ISBN 9780123741448 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Jump up to: Перейти обратно: а б с д и ж Стивенс, Элисон Н. П. (2010). «Хищничество, травоядность и паразитизм» . Знания о природном образовании . 3 (10): 36.
- ^ «Хищники, паразиты и паразитоиды» . Австралийский музей . Проверено 19 сентября 2018 г.
- ^ Ватанабэ, Джеймс М. (2007). «Беспозвоночные, обзор». В Денни, Марк В.; Гейнс, Стивен Дин (ред.). Энциклопедия приливов и скалистых берегов . Издательство Калифорнийского университета. ISBN 9780520251182 .
- ^ Фелан, Джей (2009). Что такое жизнь? : пособие по биологии (Студенческая ред.). WH Freeman & Co. с. 432. ИСБН 9781429223188 .
- ^ Вильянуэва, Роджер; Перриконе, Валентина; Фиорито, Грациано (17 августа 2017 г.). «Головоногие моллюски как хищники: короткое путешествие среди поведенческих гибкостей, адаптаций и пищевых привычек» . Границы в физиологии . 8 : 598. дои : 10.3389/fphys.2017.00598 . ПМЦ 5563153 . ПМИД 28861006 .
- ^ Ханссен, Свейнн Аре; Эрикстад, Челль Эйнар (2012). «Долгосрочные последствия хищничества яиц» . Поведенческая экология . 24 (2): 564–569. дои : 10.1093/beheco/ars198 .
- ^ Пайк, Дэвид А.; Кларк, Рулон В.; Маника, Андреа; Ценг, Хуэй-Юнь; Сюй, Юнг-Я; Хуан, Вэнь-Сан (26 февраля 2016 г.). «Прибой и газон: хищничество змей, питающихся яйцами, привело к эволюции родительской заботы у наземной ящерицы» . Научные отчеты . 6 (1): 22207. Бибкод : 2016NatSR...622207P . дои : 10.1038/srep22207 . ПМЦ 4768160 . ПМИД 26915464 .
- ^ Эйнсворт, Джилл; Калладайн, Джон; Мартей, Блез; Парк, Кирсти; Редпат, Стив; Вернхэм, Крис; Уилсон, Марк; Янг, Джульетта (2017). Понимание хищничества: обзор, объединяющий естественные науки и местные знания о недавних изменениях в популяциях диких птиц и их движущих силах в Шотландии . Шотландский форум Морленд. стр. 233–234. дои : 10.13140/RG.2.1.1014.6960 .
- ^ Хедрих, Райнер; Фукусима, Кендзи (20 мая 2021 г.). «О происхождении плотоядности: молекулярная физиология и эволюция растений на животной диете» . Ежегодный обзор биологии растений . 72 (1). аннурев–арплант–080620-010429. doi : 10.1146/annurev-arplant-080620-010429 . ISSN 1543-5008 . ПМИД 33434053 . S2CID 231595236 .
- ^ Прамер, Д. (1964). «Грибки, ловящие нематод». Наука . 144 (3617): 382–388. Бибкод : 1964Sci...144..382P . дои : 10.1126/science.144.3617.382 . JSTOR 1713426 . ПМИД 14169325 .
- ^ Jump up to: Перейти обратно: а б Велисер, Грегори Дж.; Мендес-Соареш, Хелена (2007). «Бактериальные хищники» (PDF) . Клетка . 19 (2): Р55–Р56. дои : 10.1016/j.cub.2008.10.043 . ПМИД 19174136 . S2CID 5432036 .
- ^ Jump up to: Перейти обратно: а б Юркевич, Эдуард; Давыдов, Яаков (2006). «Филогенетическое разнообразие и эволюция хищных прокариот». Хищные прокариоты . Спрингер. стр. 11–56 . дои : 10.1007/7171_052 . ISBN 978-3-540-38577-6 .
- ^ Хансен, Пер Жюэль; Бьёрнсен, Питер Кёфед; Хансен, Бенни Виндинг (1997). «Выпас и рост зоопланктона: масштабирование в диапазоне размеров тела 2–2 мкм» . Лимнология и океанография . 42 (4): 687–704. Бибкод : 1997LimOc..42..687H . дои : 10.4319/lo.1997.42.4.0687 . обобщает выводы многих авторов.
- ^ Jump up to: Перейти обратно: а б с д и ж Крамер, Дональд Л. (2001). «Собирательное поведение» (PDF) . В Фоксе, CW; Рофф, Д.А.; Фэйрберн, диджей (ред.). Эволюционная экология: концепции и тематические исследования . Издательство Оксфордского университета. стр. 232–238. ISBN 9780198030133 . Архивировано из оригинала (PDF) 12 июля 2018 года . Проверено 20 сентября 2018 г.
- ^ Jump up to: Перейти обратно: а б Гриффитс, Дэвид (ноябрь 1980 г.). «Затраты на добычу пищи и относительный размер добычи». Американский натуралист . 116 (5): 743–752. дои : 10.1086/283666 . JSTOR 2460632 . S2CID 85094710 .
- ^ Ветцель, Роберт Г.; Ликенс, Джин Э. (2000). «Взаимодействие хищника и жертвы» . Лимнологический анализ . Спрингер. стр. 257–262 . дои : 10.1007/978-1-4757-3250-4_17 . ISBN 978-1-4419-3186-3 .
- ^ Jump up to: Перейти обратно: а б с д Пианка, Эрик Р. (2011). Эволюционная экология (7-е (электронная книга) изд.). Эрик Р. Пианка. стр. 78–83.
- ^ Макартур, Роберт Х. (1984). «Экономика потребительского выбора». Географическая экология: закономерности распространения видов . Издательство Принстонского университета. стр. 59–76. ISBN 9780691023823 .
- ^ Jump up to: Перейти обратно: а б с д Белл 2012 , стр. 4–5.
- ^ Истман, Лукас Б.; Тиль, Мартин (2015). «Собирательное поведение ракообразных хищников и падальщиков». В Тиле, Мартин; Уотлинг, Лес (ред.). Образ жизни и биология питания . Издательство Оксфордского университета. стр. 535–556. ISBN 9780199797066 .
- ^ Перри, Гад (январь 1999 г.). «Эволюция режимов поиска: экологические и филогенетические перспективы». Американский натуралист . 153 (1): 98–109. дои : 10.1086/303145 . ПМИД 29578765 . S2CID 4334462 .
- ^ Jump up to: Перейти обратно: а б Белл 2012 , стр. 69–188.
- ^ Гремийе, Д.; Уилсон, Р.П.; Ванлесс, С.; Чейтер, Т. (2000). «Чернобровые альбатросы, международное рыболовство и Патагонский шельф» . Серия «Прогресс в области морской экологии» . 195 : 69–280. Бибкод : 2000MEPS..195..269G . дои : 10.3354/meps195269 .
- ^ Чарнов, Эрик Л. (1976). «Оптимальный поиск пищи, теорема о предельной ценности» (PDF) . Теоретическая популяционная биология . 9 (2): 129–136. дои : 10.1016/0040-5809(76)90040-x . ПМИД 1273796 .
- ^ Рейнольдс, Энди (сентябрь 2015 г.). «Освобождение исследований ходьбы Леви от оков оптимального поиска пищи». Обзоры физики жизни . 14 : 59–83. Бибкод : 2015PhLRv..14...59R . дои : 10.1016/j.plrev.2015.03.002 . ПМИД 25835600 .
- ^ Бьюкенен, Марк (5 июня 2008 г.). «Экологическое моделирование: математическое зеркало животной природы» . Природа . 453 (7196): 714–716. дои : 10.1038/453714a . ПМИД 18528368 .
- ^ Уильямс, Аманда С.; Флаксман, Сэмюэл М. (2012). «Могут ли хищники оценить качество ресурсов своей добычи?». Поведение животных . 83 (4): 883–890. дои : 10.1016/j.anbehav.2012.01.008 . S2CID 53172079 .
- ^ Шарф, Инон; Нульман, Эйнат; Овадия, Офер; Бускила, Амос (сентябрь 2006 г.). «Оценка эффективности двух конкурирующих способов добывания пищи в разных условиях». Американский натуралист . 168 (3): 350–357. дои : 10.1086/506921 . ПМИД 16947110 . S2CID 13809116 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к Мур, Талия Ю.; Бивенер, Эндрю А. (2015). «Обгонять или перехитрить: взаимодействие хищника и жертвы как модельная система для интеграции биомеханических исследований в более широкий экологический и эволюционный контекст» (PDF) . Интегративная и сравнительная биология . 55 (6): 1188–97. дои : 10.1093/icb/icv074 . ПМИД 26117833 .
- ^ Jump up to: Перейти обратно: а б де Врис, MS; Мерфи, EAK; Патек СН (2012 г.). «Механика удара засадного хищника: копьеносая креветка-богомол» . Журнал экспериментальной биологии . 215 (Часть 24): 4374–4384. дои : 10.1242/jeb.075317 . ПМИД 23175528 .
- ^ «Пума» . Внутренние районы Кто есть кто . Канадская служба дикой природы и Канадская федерация дикой природы . Архивировано из оригинала 18 мая 2007 года . Проверено 22 мая 2007 г.
- ^ «Щуки (Esocidae)» (PDF) . Индианский отдел рыбы и дикой природы . Проверено 3 сентября 2018 г.
- ^ Брей, Дайан. «Восточная рыба-лягушка, Batrachomoeus dubius» . Рыбы Австралии . Архивировано из оригинала 14 сентября 2014 года . Проверено 14 сентября 2014 г.
- ^ «Пауки-люки» . Би-би-си . Проверено 12 декабря 2014 г.
- ^ «Паук-люк» . Музей пустыни Аризона-Сонора. 2014 . Проверено 12 декабря 2014 г.
- ^ Газда, СК; Коннор, RC; Эдгар, РК; Кокс, Ф. (2005). «Разделение труда с ролевой специализацией на групповой охоте на афалин (Tursiops truncatus) у Сидар-Ки, Флорида» . Труды Королевского общества . 272 (1559): 135–140. дои : 10.1098/rspb.2004.2937 . ПМК 1634948 . ПМИД 15695203 .
- ^ Тайс, Гарольд М. (2011). Экология и охрана рыб . ЦРК Пресс. п. 233. ИСБН 978-1-4398-9759-1 .
- ^ Комбс, ЮАР; Сальседо, МК; Пандит, ММ; Ивасаки, Дж. М. (2013). «Определите успех и эффективность стрекоз, преследующих разные виды добычи» . Интегративная и сравнительная биология . 53 (5): 787–798. дои : 10.1093/icb/ict072 . ПМИД 23784698 .
- ^ Хубель, Татьяна Ю.; Мятт, Джулия П.; Джордан, Нил Р.; Дьюхерст, Оливер П.; МакНатт, Дж. Уэлдон; Уилсон, Алан М. (29 марта 2016 г.). «Затраты энергии и прибыль от охоты на африканских диких собак» . Природные коммуникации . 7 : 11034. дои : 10.1038/ncomms11034 . ПМЦ 4820543 . ПМИД 27023457 .
Стратегии беглой охоты варьируются от одной крайности — кратковременного ускорения, силы и скорости до другой крайности — настойчивости и выносливости, при этом добыча утомляется, чтобы облегчить поимку. Считается, что собаки и люди полагаются на выносливость, а не на прямую скорость и маневренность для достижения успеха при беглой охоте. .
- ^ Гольдбоген, Дж.А.; Каламбокидис, Дж.; Шедвик, RE; Олесон, EM; Макдональд, Массачусетс; Хильдебранд, JA (2006). «Кинематика кормового ныряния и кормления финвалов с выпада» . Журнал экспериментальной биологии . 209 (7): 1231–1244. дои : 10.1242/jeb.02135 . ПМИД 16547295 . S2CID 17923052 .
- ^ Сандерс, Джон Г.; Бейхман, Аннабель К.; Роман, Джо; Скотт, Джаррод Дж.; Эмерсон, Дэвид; Маккарти, Джеймс Дж.; Гиргис, Питер Р. (2015). «У усатых китов имеется уникальный кишечный микробиом, сходный как с плотоядными, так и с травоядными» . Природные коммуникации . 6 : 8285. Бибкод : 2015NatCo...6.8285S . дои : 10.1038/ncomms9285 . ПМЦ 4595633 . ПМИД 26393325 .
- ^ Форбс, Л. Скотт (1989). «Защита от добычи и поведение при обращении с хищниками: гипотеза об опасной добыче». Ойкос . 55 (2): 155–158. Бибкод : 1989Oikos..55..155F . дои : 10.2307/3565418 . JSTOR 3565418 .
- ^ Jump up to: Перейти обратно: а б Ланг, Стивен DJ; Фарин, Дэмиен Р. (2017). «Многомерная основа для изучения стратегий социального хищничества» . Экология и эволюция природы . 1 (9): 1230–1239. Бибкод : 2017NatEE...1.1230L . дои : 10.1038/s41559-017-0245-0 . ПМИД 29046557 . S2CID 4214982 .
- ^ МакНалти, Дэниел Р.; Таллиан, Эйми; Сталер, Дэниел Р.; Смит, Дуглас В. (12 ноября 2014 г.). Сюёр, Седрик (ред.). «Влияние размера группы на успех волков в охоте на бизонов» . ПЛОС ОДИН . 9 (11): e112884. Бибкод : 2014PLoSO...9k2884M . дои : 10.1371/journal.pone.0112884 . ПМЦ 4229308 . ПМИД 25389760 .
- ^ Пауэр, Р.Дж.; Компион, RX Shem (2009). «Похищение львами слонов в Савути, национальный парк Чобе, Ботсвана» . Африканская зоология . 44 (1): 36–44. дои : 10.3377/004.044.0104 . S2CID 86371484 .
- ^ Бошан 2012 , стр. 7–12.
- ^ Доусон, Джеймс В. (1988). Система кооперативного разведения ястреба Харриса в Аризоне (магистерская диссертация). Университет Аризоны . Проверено 17 ноября 2017 г.
- ^ Вейл, Александр Л.; Маника, Андреа; Бшары, Редуан (23 апреля 2013 г.). «Указательные жесты в совместной рыбной охоте» . Природные коммуникации . 4 (1): 1765. Бибкод : 2013NatCo...4.1765V . дои : 10.1038/ncomms2781 . ПМИД 23612306 .
- ^ Йонг, Эд (24 апреля 2013 г.). «Группировщики используют жесты, чтобы вербовать мурен в охотничьи отряды» . Нэшнл Географик . Архивировано из оригинала 17 сентября 2018 года . Проверено 17 сентября 2018 г.
- ^ Тофт, Клаус (продюсер) (2007). Убийцы в Эдеме (документальный DVD) . Австралийская радиовещательная корпорация . Архивировано из оригинала 12 августа 2009 года. ISBN R-105732-9.
- ^ Jump up to: Перейти обратно: а б Брайс, Калеб М.; Уилмерс, Кристофер С.; Уильямс, Терри М. (2017). «Энергетика и динамика уклонения крупных хищников и добычи: пумы против гончих» . ПерДж . 5 : е3701. дои : 10.7717/peerj.3701 . ПМЦ 5563439 . ПМИД 28828280 .
- ^ Майер, Мария; Холм, Кристина; Любин, Яэль; Бильде, Трина (2018). «Совместный поиск пищи расширяет пищевую нишу, но не компенсирует внутригрупповую конкуренцию за ресурсы у социальных пауков» . Научные отчеты . 8 (1): 11828. Бибкод : 2018НатСР...811828М . дои : 10.1038/s41598-018-30199-x . ПМК 6081395 . ПМИД 30087391 .
- ^ «Хищники из засады» . Природный центр Сибли . Архивировано из оригинала 2 августа 2021 года . Проверено 17 сентября 2018 г.
- ^ Эльброх, Л. Марк; Куигли, Ховард (10 июля 2016 г.). «Социальные взаимодействия одиночного хищника» . Современная зоология . 63 (4): 357–362. дои : 10.1093/cz/zow080 . ПМК 5804185 . ПМИД 29491995 .
- ^ Кенкуа, Дуглас (11 октября 2017 г.). «Одинокие пумы оказались горными львами, которые обедают» . Нью-Йорк Таймс . Проверено 17 сентября 2018 г.
- ^ Флорес, Дэн (2016). Койотская Америка: естественная и сверхъестественная история . Основные книги. ISBN 978-0465052998 .
- ^ Стоу, Адам; Нюквист, Марина Дж.; Гозлан, Родольф Э.; Кушеруссе, Жюльен; Бриттон, Дж. Роберт (2012). «Поведенческий синдром одиночного хищника не зависит от размера тела и скорости роста» . ПЛОС ОДИН . 7 (2): e31619. Бибкод : 2012PLoSO...731619N . дои : 10.1371/journal.pone.0031619 . ПМЦ 3282768 . ПМИД 22363687 .
- ^ «Как охотятся пауки?» . Американский музей естественной истории. 25 августа 2014 года . Проверено 5 сентября 2018 г.
- ^ Весело, Рональд М.; Заяц, Дж. Дэниел (2009). «Хищничество/Хищные насекомые». Энциклопедия насекомых (второе изд.). стр. 837–839. дои : 10.1016/B978-0-12-374144-8.00219-8 . ISBN 9780123741448 .
- ^ «Зоопланктон» . Общество охраны морской биологии. 17 июня 2018 г. Проверено 5 сентября 2018 г.
- ^ Бар-Ям. «Отношения хищник-жертва» . Институт сложных систем Новой Англии . Проверено 7 сентября 2018 г.
- ^ Jump up to: Перейти обратно: а б с «Хищник и жертва: Адаптации» (PDF) . Королевский музей Саскачевана. 2012. Архивировано из оригинала (PDF) 3 апреля 2018 года . Проверено 19 апреля 2018 г.
- ^ Вермей, Гират Дж. (1993). Эволюция и эскалация: экологическая история жизни . Издательство Принстонского университета. стр. 11 и пассим. ISBN 978-0-691-00080-0 .
- ^ Гетц, WM (2011). «Сети преобразования биомассы обеспечивают единый подход к моделированию потребительских ресурсов» . Экологические письма . 14 (2): 113–24. Бибкод : 2011EcolL..14..113G . дои : 10.1111/j.1461-0248.2010.01566.x . ПМК 3032891 . ПМИД 21199247 .
- ^ Сидорович, Вадим (2011). Анализ сообщества хищник-жертва позвоночных: исследования в пределах Европейской лесной зоны на территориях с переходным смешанным лесом в Беларуси . Тесей. п. 426. ИСБН 978-985-463-456-2 .
- ^ Анджеличи, Франческо М. (2015). Проблемная дикая природа: междисциплинарный подход . Спрингер. п. 160. ИСБН 978-3-319-22246-2 .
- ^ Хейворд, штат Вашингтон; Хеншель, П.; О'Брайен, Дж.; Хофмейр, М.; Бальме, Г.; Керли, GIH (2006). «Хищные предпочтения леопарда ( Panthera pardus )» (PDF) . Журнал зоологии . 270 (2): 298–313. дои : 10.1111/j.1469-7998.2006.00139.x . Архивировано из оригинала (PDF) 29 февраля 2020 года . Проверено 20 апреля 2018 г.
- ^ Пуллиам, Х. Рональд (1974). «К теории оптимальных диет». Американский натуралист . 108 (959): 59–74. дои : 10.1086/282885 . S2CID 8420787 .
- ^ Сих, Эндрю; Кристенсен, Бент (2001). «Теория оптимальной диеты: когда она работает, а когда и почему терпит неудачу?». Поведение животных . 61 (2): 379–390. дои : 10.1006/anbe.2000.1592 . S2CID 44045919 .
- ^ Спрулс, В. Гэри (1972). «Влияние хищничества и пищевой конкуренции в зависимости от размера на высокогорные сообщества зоопланктона». Экология . 53 (3): 375–386. Бибкод : 1972Экол...53..375С . дои : 10.2307/1934223 . JSTOR 1934223 .
- ^ Оуэн-Смит, Норман; Миллс, MGL (2008). «Отношения размеров хищника и добычи в пищевой сети крупных млекопитающих Африки» (PDF) . Журнал экологии животных . 77 (1): 173–183. Бибкод : 2008JAnEc..77..173O . дои : 10.1111/j.1365-2656.2007.01314.x . hdl : 2263/9023 . ПМИД 18177336 .
- ^ Котт 1940 , стр. 12–13.
- ^ Ллойд Дж. Э. (1965). «Агрессивная мимикрия в Photuris: Firefly Femmes Fatales». Наука . 149 (3684): 653–654. Бибкод : 1965Sci...149..653L . дои : 10.1126/science.149.3684.653 . ПМИД 17747574 . S2CID 39386614 .
- ^ Форбс, Питер (2009). Ослепленный и обманутый: мимикрия и камуфляж . Издательство Йельского университета. п. 134. ИСБН 978-0-300-17896-8 .
- ^ Бестер, Кэтлин (5 мая 2017 г.). «Антеннариус полосатый» . Музей Флориды . Университет Флориды . Проверено 31 января 2018 г.
- ^ Руперт, Эдвард Э.; Фокс, Ричард, С.; Барнс, Роберт Д. (2004). Зоология беспозвоночных, 7-е издание . Cengage Обучение. стр. 153–154. ISBN 978-81-315-0104-7 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Четарук, Эдвард В. (2005). «Гремучие змеи и другие кроталиды» . В Бренте, Джеффри (ред.). Токсикология интенсивной терапии: диагностика и лечение тяжело отравленных пациентов . Elsevier Науки о здоровье. п. 1075. ИСБН 978-0-8151-4387-1 .
- ^ Барселу, Дональд Г. (2008). Медицинская токсикология природных веществ: продукты питания, грибы, лекарственные травы, растения и ядовитые животные . Уайли. п. 1028. ИСБН 978-0-470-33557-4 .
- ^ Ли, Мин; Фрай, Б.Г.; Кини, Р. Манджунатха (2005). «Диета, состоящая только из яиц: ее влияние на изменения профиля токсинов и экологию мраморной морской змеи (Aipysurus eydouxii)». Журнал молекулярной эволюции . 60 (1): 81–89. Бибкод : 2005JMolE..60...81L . дои : 10.1007/s00239-004-0138-0 . ПМИД 15696370 . S2CID 17572816 .
- ^ Кастелло, Мэн; А. Родригес-Каттанео; П.А. Агилера; Л. Ирибарн; АК Перейра; А. А. Капути (2009). «Генерация сигналов у слабоэлектрических рыб Gymnotus coropinae (Hoedeman): электрический орган и разряд электрического органа» . Журнал экспериментальной биологии . 212 (9): 1351–1364. дои : 10.1242/jeb.022566 . ПМИД 19376956 .
- ^ Фёлнер, П.Г.; М. Плат; Дж. Энгельманн; Ф. Киршбаум; Р. Тидеманн (2009). «Электрифицирующая любовь: электрические рыбы используют видоспецифичный разряд для распознавания партнера» . Письма по биологии . 5 (2): 225–228. дои : 10.1098/rsbl.2008.0566 . ПМЦ 2665802 . ПМИД 19033131 .
- ^ Катания, Кеннет К. (2015). «Электрические угри используют высокое напряжение, чтобы отслеживать быстро движущуюся добычу» . Природные коммуникации . 6 (1): 8638. Бибкод : 2015NatCo...6.8638C . дои : 10.1038/ncomms9638 . ПМЦ 4667699 . ПМИД 26485580 .
- ^ Крамер, Бернд (1996). «Электрорецепция и коммуникация у рыб» (PDF) . Прогресс в зоологии . 42 .
- ^ Карасов, Уильям Х.; Даймонд, Джаред М. (1988). «Взаимодействие физиологии и экологии пищеварения». Бионаука . 38 (9): 602–611. дои : 10.2307/1310825 . JSTOR 1310825 .
- ^ Ruxton, Sherratt & Speed 2004 , стр. vii–xii.
- ^ Каро 2005 , стр. 67–114
- ^ Мерайлайта, Сами; Скотт-Сэмюэл, Николас Э.; Катхилл, Иннес К. (22 мая 2017 г.). «Как работает камуфляж» . Философские труды Королевского общества B: Биологические науки (Представленная рукопись). 372 (1724): 20160341. doi : 10.1098/rstb.2016.0341 . ПМК 5444062 . ПМИД 28533458 .
- ^ Каро 2005 , стр. 13–15
- ^ Бергстрем, Коннектикут; Лахманн, М. (2001). «Тревожные сигналы как дорогостоящие сигналы бдительности против хищников: игра бдительных болтунов». Поведение животных . 61 (3): 535–543. CiteSeerX 10.1.1.28.773 . дои : 10.1006/anbe.2000.1636 . S2CID 2295026 .
- ^ Гетти, Т. (2002). «Проницательный болтун встречает ястреба, придерживающегося оптимальной диеты». Поведение животных . 63 (2): 397–402. дои : 10.1006/anbe.2001.1890 . S2CID 53164940 .
- ^ Котт 1940 , стр. 241–307.
- ^ Бауэрс, доктор медицины; Браун, Ирен Л.; Уэй, Дэррил (1985). «Хищничество птиц как селективный агент в популяции бабочек» . Эволюция . 39 (1): 93–103. дои : 10.1111/j.1558-5646.1985.tb04082.x . ПМИД 28563638 . S2CID 12031679 .
- ^ Беренбаум, MR (3 января 1995 г.). «Химия защиты: теория и практика» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (1): 2–8. Бибкод : 1995ПНАС...92....2Б . дои : 10.1073/pnas.92.1.2 . ПМК 42807 . ПМИД 7816816 .
- ^ Ruxton, Sherratt & Speed 2004 , стр. 70–81.
- ^ Каро 2005 , стр. 663–684
- ^ Бошан 2012 , стр. 83–88.
- ^ Краузе, Йенс; Ракстон, Грэм Д. (10 октября 2002 г.). Проживание группами . Издательство Оксфордского университета. стр. 13–15. ISBN 9780198508182 .
- ^ Ракстон, Шерратт и Спид 2004 , стр. 54–55.
- ^ Домини, Уоллес Дж. (1983). «Моббинг среди колониально гнездящихся рыб, особенно синежабрников, Lepomis macrochirus ». Копейя . 1983 (4): 1086–1088. дои : 10.2307/1445113 . JSTOR 1445113 .
- ^ Броди, Эдмунд Д. (3 ноября 2009 г.). «Токсины и яды» (PDF) . Современная биология . 19 (20): Р931–Р935. Бибкод : 2009CBio...19.R931B . дои : 10.1016/j.cub.2009.08.011 . ПМИД 19889364 . S2CID 9744565 .
- ^ Котт 1940 , стр. 368–389.
- ^ Мерайлайта, Сами; Валлин, Адриан; Кодандарамайя, Улласа; и др. (26 июля 2011 г.). «Количество глазных пятен и их устрашающее воздействие на наивных хищников у бабочки-павлина» . Поведенческая экология . 22 (6): 1326–1331. дои : 10.1093/beheco/arr135 .
- ^ Эдмундс, Малькольм (2012). «Дейматическое поведение» . Спрингер . Проверено 31 декабря 2012 г.
- ^ Каро 2005 , стр. v – xi, 4–5.
- ^ Каро 2005 , с. 413–414
- ^ Джейкобс и Бастиан 2017 , с. 4
- ^ Барбоса, Педро; Кастелланос, Игнасио (2005). Экология взаимодействия хищник-жертва . Издательство Оксфордского университета. п. 78 . ISBN 9780199874545 .
- ^ Янис, СМ; Вильгельм, ПБ (1993). «Существовали ли в третичном периоде хищники, преследующие млекопитающих? Танцы с аватарами волков». Журнал эволюции млекопитающих . 1 (2): 103–125. дои : 10.1007/bf01041590 . S2CID 22739360 .
- ^ Jump up to: Перейти обратно: а б с Докинз, Ричард ; Кребс, младший (1979). «Гонка вооружений между видами и внутри них». Труды Королевского общества B: Биологические науки . 205 (1161): 489–511. Бибкод : 1979РСПСБ.205..489Д . дои : 10.1098/rspb.1979.0081 . ПМИД 42057 . S2CID 9695900 .
- ^ Jump up to: Перейти обратно: а б Абрамс, Питер А. (ноябрь 1986 г.). «Адаптивные реакции хищников на добычу и жертв на хищников: провал аналогии с гонкой вооружений» . Эволюция . 40 (6): 1229–1247. дои : 10.1111/j.1558-5646.1986.tb05747.x . ПМИД 28563514 . S2CID 27317468 .
- ^ Jump up to: Перейти обратно: а б с д и Броди, Эдмунд Д. (июль 1999 г.). «Гонка вооружений Хищник-жертва» . Бионаука . 49 (7): 557–568. дои : 10.2307/1313476 . JSTOR 1313476 .
- ^ Вермей, Г.Дж. (ноябрь 1994 г.). «Эволюционное взаимодействие видов: отбор, эскалация и коэволюция». Ежегодный обзор экологии и систематики . 25 (1): 219–236. doi : 10.1146/annurev.es.25.110194.001251 .
- ^ Джейкобс и Бастиан 2017 , с. 8
- ^ Джейкобс и Бастиан 2017 , с. 107
- ^ Шериф Майкл Дж.; Пикор, Скотт Д.; Хаулена, Дрор; Такер, Мария; Гайяр, Жан-Мишель (2020). «Влияние не чахоточных хищников на размер популяции жертв: недостаток доказательств» . Журнал экологии животных . 89 (6): 1302–1316. Бибкод : 2020JAnEc..89.1302S . дои : 10.1111/1365-2656.13213 . ПМИД 32215909 .
- ^ Прейссер, Эван Л.; Больник, Дэниел И.; Бенард, Майкл Ф. (2005). «Напуган до смерти? Эффекты запугивания и потребления во взаимодействии хищника и жертвы». Экология . 86 (2): 501–509. Бибкод : 2005Экол...86..501П . дои : 10.1890/04-0719 . ISSN 0012-9658 .
- ^ Jump up to: Перейти обратно: а б Пекарски, Барбара Л.; Абрамс, Питер А.; Больник, Дэниел И.; Дилл, Лоуренс М.; Грабовски, Джонатан Х.; Латтбег, Барни; Оррок, Джон Л.; Пикор, Скотт Д.; Прейссер, Эван Л.; Шмитц, Освальд Дж.; Трасселл, Джеффри К. (сентябрь 2008 г.). «Возвращаясь к классике: рассмотрение непотребительских эффектов в хрестоматийных примерах взаимодействия хищник-жертва» . Экология . 89 (9): 2416–2425. Бибкод : 2008Ecol...89.2416P . дои : 10.1890/07-1131.1 . ПМИД 18831163 .
- ^ Линдеман, Раймонд Л. (1942). «Трофо-динамический аспект экологии». Экология . 23 (4): 399–417. Бибкод : 1942Экол...23..399Л . дои : 10.2307/1930126 . JSTOR 1930126 .
- ^ Ордис, Андрес; Бишоф, Ричард; Свенсон, Джон Э. (2013). «Спасти крупных хищников, но потерять высшего хищника?» . Биологическая консервация . 168 : 128–133. Бибкод : 2013BCons.168..128O . дои : 10.1016/j.biocon.2013.09.024 . HDL : 11250/2492589 .
- ^ Пимм, СЛ; Лоутон, Дж. Х. (1978). «О питании более чем на одном трофическом уровне». Природа . 275 (5680): 542–544. Бибкод : 1978Natur.275..542P . дои : 10.1038/275542a0 . S2CID 4161183 .
- ^ Федриани, Дж. М.; Фуллер, ТК; Соважо, РМ; Йорк, ЕС (2000). «Конкуренция и внутригильдейское хищничество среди трех симпатрических хищников» (PDF) . Экология . 125 (2): 258–270. Бибкод : 2000Oecol.125..258F . дои : 10.1007/s004420000448 . hdl : 10261/54628 . ПМИД 24595837 . S2CID 24289407 .
- ^ Лалли, СМ; Парсонс, Т.Р. (1997). Биологическая океанография: Введение (2-е изд.). Открытый университет. п. 116.
- ^ «Перенос энергии в экосистемах» . Нэшнл Географик . 18 февраля 2023 г. Проверено 18 февраля 2023 г.
- ^ Бонд, WJ (2012). «11. Краеугольные виды». В Шульце, Эрнст-Детлеф; Муни, Гарольд А. (ред.). Биоразнообразие и функции экосистемы . Спрингер. п. 237. ИСБН 978-3642580017 .
- ^ Боткин Д.; Келлер, Э. (2003). Наука об окружающей среде: Земля как живая планета . Джон Уайли и сыновья. п. 2. ISBN 978-0-471-38914-9 .
- ^ Jump up to: Перейти обратно: а б Риппл, Уильям Дж.; Бешта, Роберт Л. (2004). «Волки и экология страха: может ли риск хищничества структурировать экосистемы?» . Бионаука . 54 (8): 755. doi : 10.1641/0006-3568(2004)054[0755:WATEOF]2.0.CO;2 .
- ^ Нил, Дик (2004). Введение в популяционную биологию . Издательство Кембриджского университета. стр. 68–69. ISBN 9780521532235 .
- ^ Нельсон, Эрик Х.; Мэтьюз, Кристофер Э.; Розенхайм, Джей А. (июль 2004 г.). «Хищники сокращают рост популяции жертв, вызывая изменения в их поведении» . Экология . 85 (7): 1853–1858. Бибкод : 2004Ecol...85.1853N . дои : 10.1890/03-3109 . JSTOR 3450359 .
- ^ Кребс, Чарльз Дж.; Бунстра, Руди; Бутин, Стэн; Синклер, ARE (2001). «Что движет 10-летним циклом зайцев на снегоступах?» . Бионаука . 51 (1): 25. doi : 10.1641/0006-3568(2001)051[0025:WDTYCO]2.0.CO;2 . HDL : 1807/359 .
- ^ Кребс, Чарли; Майерс, Джуди (12 июля 2014 г.). «10-летний цикл зайца на снегоступах - поучительная история» . Экологические тирады . Университет Британской Колумбии . Проверено 2 октября 2018 г.
- ^ «Хищники и их жертвы» . BBC Bitesize . Би-би-си . Проверено 7 октября 2015 г.
- ^ Гоэл, Нарендра С.; Майтра, Южная Каролина; Монтролл, EW (1971). О Вольтерре и других нелинейных моделях взаимодействующих популяций . Академическая пресса. ISBN 978-0122874505 .
- ^ Jump up to: Перейти обратно: а б Левин, Саймон А.; Карпентер, Стивен Р.; Годфрей, Х. Чарльз Дж.; Кинциг, Энн П.; Лоро, Мишель; Лосос, Джонатан Б.; Уокер, Брайан; Уилков, Дэвид С. (2009). Принстонский справочник по экологии . Издательство Принстонского университета. стр. 204–209 . ISBN 9781400833023 .
- ^ Мердок, Уильям В.; Бриггс, Шерил Дж.; Нисбет, Роджер М. (2013). Динамика потребительских ресурсов . Издательство Принстонского университета. п. 39. ИСБН 9781400847259 .
- ^ Новак, Мартин; Мэй, Роберт М. (2000). Динамика вирусов: математические основы иммунологии и вирусологии . Издательство Оксфордского университета. п. 8. ISBN 9780191588518 .
- ^ Дженоварт, М.; Негре, Н.; Тавеккья, Г.; Бистуэр, А.; Парпал, Л.; Оро, Д. (2010). «Молодые, слабые и больные: свидетельства естественного отбора посредством хищничества» . ПЛОС ОДИН . 5 (3): e9774. Бибкод : 2010PLoSO...5.9774G . дои : 10.1371/journal.pone.0009774 . ПМЦ 2841644 . ПМИД 20333305 .
- ^ Роквуд 2009 , с. 281
- ^ Роквуд 2009 , с. 246
- ^ Роквуд 2009 , стр. 271–272.
- ^ Роквуд 2009 , с. 272–273
- ^ Jump up to: Перейти обратно: а б Кушинг, Дж. М. (2005). «Рецензии на книги | Математика в популяционной биологии Хорста Р. Тиена» (PDF) . Бюллетень Американского математического общества . 42 (4): 501–505. дои : 10.1090/S0273-0979-05-01055-4 .
- ^ Тиме, Хорст Р. (2003). Математика в популяционной биологии . Издательство Принстонского университета. ISBN 978-0-691-09291-1 .
- ^ Козлов Владимир; Вакуленко, Сергей (3 июля 2013 г.). «О хаосе в системах Лотка – Вольтерра: аналитический подход». Нелинейность . 26 (8): 2299–2314. Бибкод : 2013Nonli..26.2299K . дои : 10.1088/0951-7715/26/8/2299 . S2CID 121559550 .
- ^ Сих, Эндрю (1987). «Убежища для добычи и стабильность хищник-жертва». Теоретическая популяционная биология . 31 : 1–12. дои : 10.1016/0040-5809(87)90019-0 .
- ^ Макнейр, Джеймс Н. (1986). «Влияние убежищ на взаимодействие хищник-жертва: пересмотр». Теоретическая популяционная биология . 29 (1): 38–63. дои : 10.1016/0040-5809(86)90004-3 . ПМИД 3961711 .
- ^ Берриман, Алан А.; Хокинс, Брэдфорд А.; Хокинс, Брэдфорд А. (2006). «Убежище как интегрирующая концепция экологии и эволюции». Ойкос . 115 (1): 192–196. Бибкод : 2006Oikos.115..192B . дои : 10.1111/j.0030-1299.2006.15188.x .
- ^ Крессман, Росс; Гарай, Йожеф (2009). «Система убежища хищник-жертва: эволюционная стабильность экологических систем». Теоретическая популяционная биология . 76 (4): 248–57. дои : 10.1016/j.tpb.2009.08.005 . ПМИД 19751753 .
- ^ Абрамс, Пенсильвания (2000). «Эволюция взаимодействия хищник-жертва: теория и доказательства». Ежегодный обзор экологии и систематики . 31 : 79–105. doi : 10.1146/annurev.ecolsys.31.1.79 .
- ^ Jump up to: Перейти обратно: а б Гримальди, Дэвид ; Энгель, Майкл С. (2005). Эволюция насекомых . Издательство Кембриджского университета. стр. 155–160 . ISBN 978-0-521-82149-0 .
- ^ Грант, SWF; Нолл, АХ; Микробы, GJB (1991). «Вероятные кальцинированные метафиты в новейшей протерозойской группе Нама, Намибия: происхождение, диагенез и последствия». Журнал палеонтологии . 65 (1): 1–18. Бибкод : 1991JPal...65....1G . дои : 10.1017/S002233600002014X . JSTOR 1305691 . ПМИД 11538648 . S2CID 26792772 .
- ^ Авраамик, С.М. (19 ноября 1971 г.). «Разнообразие столбчатых строматолитов докембрия: отражение внешнего вида многоклеточных животных». Наука . 174 (4011): 825–827. Бибкод : 1971Sci...174..825A . дои : 10.1126/science.174.4011.825 . ПМИД 17759393 . S2CID 2302113 .
- ^ Стэнли, Стивен М. (2008). «Хищничество побеждает конкуренцию на морском дне». Палеобиология . 34 (1): 1–21. Бибкод : 2008Pbio...34....1S . дои : 10.1666/07026.1 . S2CID 83713101 .
- ^ Лорон, Корентен К.; Рейнберд, Роберт Х.; Тернер, Элизабет К.; Уайлдер Гринман, Дж.; Жаво, Эммануэль Дж. (2018). «Последствия избирательного хищничества на макроэволюцию эукариот: данные из Арктической Канады». Новые темы в науках о жизни . 2 (2): 247–255. дои : 10.1042/ETLS20170153 . ПМИД 32412621 . S2CID 92505644 .
- ^ Jump up to: Перейти обратно: а б Данн, Ф.С.; Кенчингтон, штат Калифорния; Парри, Луизиана; Кларк, Дж.В.; Кендалл, РС; Уилби, PR (25 июля 2022 г.). «Книдария коронной группы из эдиакара Чарнвудского леса, Великобритания» . Экология и эволюция природы . 6 (8): 1095–1104. Бибкод : 2022NatEE...6.1095D . дои : 10.1038/s41559-022-01807-x . ПМЦ 9349040 . ПМИД 35879540 .
- ^ Келли, Патрисия (2003). Взаимодействие хищника и жертвы в летописи окаменелостей . Спрингер. стр. 113–139, 141–176 и пассим. ISBN 978-1-4615-0161-9 . OCLC 840283264 .
- ^ Дейли, Эллисон К. (2013). «Аномалокаридиды» . Современная биология . 23 (19): Р860–Р861. Бибкод : 2013CBio...23.R860D . дои : 10.1016/j.cub.2013.07.008 . ПМИД 24112975 .
- ^ Андерсон, PSL; Вестнит, М. (2009). «Биомеханическая модель кинематики питания Dunkleosteus terrelli (Arthrodira, Placodermi)». Палеобиология . 35 (2): 251–269. Бибкод : 2009Pbio...35..251A . дои : 10.1666/08011.1 . S2CID 86203770 .
- ^ Карр, Роберт К. (2010). «Палеоэкология Dunkleosteus terrelli (Placodermi: Arthrodira)» . Киртландия . 57 .
- ^ Свитек, Брайан (13 апреля 2012 г.). «Когда тираннозавр грыз зауроподов» . Журнал палеонтологии позвоночных . 25 (2): 469–472. doi : 10.1671/0272-4634(2005)025[0469:TRFTUC]2.0.CO;2 . S2CID 131583311 . Архивировано из оригинала 24 августа 2013 года . Проверено 24 августа 2013 г.
- ^ Даримонт, Коннектикут; Фокс, Швейцария; Брайан, HM; Раймхен, TE (20 августа 2015 г.). «Уникальная экология человека-хищника». Наука . 349 (6250): 858–860. Бибкод : 2015Sci...349..858D . дои : 10.1126/science.aac4249 . ПМИД 26293961 . S2CID 4985359 .
- ^ Габриэль, Отто; фон Брандт, Андрес (2005). Мировые способы ловли рыбы . Блэквелл. ISBN 978-0-85238-280-6 .
- ^ Гриффин, Эмма (2008). Кровавый спорт: охота в Британии с 1066 года . Издательство Йельского университета. ISBN 978-0300145458 .
- ^ Кинг, Ричард Дж. (1 октября 2013 г.). Дьявольский баклан: естественная история . Издательство Университета Нью-Гэмпшира. п. 9. ISBN 978-1-61168-225-0 .
- ^ Глейзер, Филипп (1998). Соколиная охота и Хокинг . Бэтсфорд. ISBN 978-0713484076 .
- ^ Эгертер, Джеймс; Фуракр, Дэвид; Смит, Грэм К. (2017). Олссон, я, Анна С. (ред.). «Первая оценка структуры и плотности популяций домашних кошек и собак по всей Великобритании» . ПЛОС ОДИН . 12 (4): e0174709. Бибкод : 2017PLoSO..1274709A . дои : 10.1371/journal.pone.0174709 . ПМК 5389805 . ПМИД 28403172 .
- ^ Гуманное общество США. «Статистика владения домашними животными в США» . Проверено 27 апреля 2012 г.
- ^ Либенберг, Луи (2008). «Значение настойчивой охоты для эволюции человека». Журнал эволюции человека . 55 (6): 1156–1159. Бибкод : 2008JHumE..55.1156L . дои : 10.1016/j.jhevol.2008.07.004 . ПМИД 18760825 .
- ^ «Пища для размышлений» (PDF) . Жизнь млекопитающих . Британская радиовещательная корпорация . 31 октября 2002 г.
- ^ Флинт, Мария Луиза; Драйштадт, Стив Х. (1998). Кларк, Джек К. (ред.). Справочник естественных врагов: Иллюстрированное руководство по биологической борьбе с вредителями . Издательство Калифорнийского университета. ISBN 978-0-520-21801-7 .
- ^ Джонстон, Кейт М. (2013). Научно-фантастический фильм: критическое введение . Издательство Берг . п. 98. ИСБН 9780857850560 .
- ^ Ньюби, Ричард (13 мая 2018 г.). «Получит ли «Хищник» наконец-то достойное продолжение?» . Голливудский репортер . Проверено 7 сентября 2018 г.
- ^ Шац, Томас. «Новый Голливуд». Киноблокбастеры . п. 25 . В: Стрингер, Джулиан (2003). Киноблокбастеры . Рутледж . стр. 15–44. ISBN 978-0-415-25608-7 .
- ^ Дэвисон, Питер (1 декабря 2002 г.). «Хищники и жертвы | Избранные стихи Теда Хьюза, 1957–1994» . Нью-Йорк Таймс . Проверено 5 октября 2018 г.
Самые ранние книги Хьюза содержали ошеломляющее обилие стихов между обложками: ... рыбы и птицы, полевые и лесные звери, энергичные воплощения хищников и добычи. Будучи студентом, Хьюз занимался антропологией, а не литературой, и решил медитировать, чтобы войти в трансоподобные состояния предсознания, прежде чем записать стихи на бумагу. Его стихи, ранние или поздние, вступают в отношения живых существ; они приближаются к животному сознанию: Мысль-Лис, Кот Эстер, Щука.
- ^ Гулд, Стивен Джей (1995). «Столетие Зуба и Когтя». Динозавр в стоге сена . Книги Гармонии. стр. 63–75. ISBN 978-0517703939 .
- ^ Jump up to: Перейти обратно: а б с д и ж Валлнер, Астрид (18 июля 2005 г.). «Роль хищников в мифологии» . Информация WaldWissen для управления лесным хозяйством. Архивировано из оригинала 5 октября 2018 года . Проверено 5 октября 2018 г. перевод из Валлнера А. (1998) Значение хищников в мифологии: результаты исследования литературы . – Информационный лист Направление исследований ландшафтной экологии 39:4–5.
- ^ Келлерт, Стивен Р.; Блэк, Мэтью; Раш, Коллин Рид; Бат, Алистер Дж. (1996). «Человеческая культура и охрана крупных хищников в Северной Америке». Биология сохранения . 10 (4): 977–990. Бибкод : 1996ConBi..10..977K . дои : 10.1046/j.1523-1739.1996.10040977.x .
Источники [ править ]
- Бошан, Гай (2012). Социальное хищничество: как групповая жизнь приносит пользу хищникам и добыче . Эльзевир. ISBN 9780124076549 .
- Белл, WJ (2012). Поисковое поведение: поведенческая экология поиска ресурсов . Спрингер Нидерланды. ISBN 9789401130981 .
- Каро, Тим (2005). Защита от хищников у птиц и млекопитающих . Издательство Чикагского университета. ISBN 978-0-226-09436-6 .
- Котт, Хью Б. (1940). Адаптивная окраска животных . Метуэн. OCLC 974070031 .
- Джейкобс, Дэвид Стив; Бастиан, Анна (2017). Взаимодействие хищник-жертва: коэволюция летучих мышей и их добычи . Спрингер. ISBN 9783319324920 .
- Роквуд, Ларри Л. (2009). Введение в популяционную экологию . Джон Уайли и сыновья. п. 281. ИСБН 9781444309102 .
- Ракстон, Грэм Д .; Шерратт, Том Н .; Скорость, Майкл П. (2004). Уклонение от нападения: эволюционная экология крипсиса, предупреждающие сигналы и мимикрия . Издательство Оксфордского университета. ISBN 9780198528593 .
Внешние ссылки [ править ]
 Цитаты, связанные с Хищничеством , в Wikiquote
Цитаты, связанные с Хищничеством , в Wikiquote  СМИ, связанные с Хищничеством , на Викискладе?
СМИ, связанные с Хищничеством , на Викискладе?

| Базы данных органов управления : Национальные |
|---|