Сообщество (экология)

В экологии сообщество — это группа или объединение популяций занимающих двух или более разных видов, одну и ту же географическую территорию одновременно, известная также как биоценоз , биотическое сообщество , биологическое сообщество , экологическое сообщество или жизненная совокупность . Термин «сообщество» имеет множество значений. В простейшей форме это относится к группам организмов в определенном месте или времени, например, «рыбное сообщество озера Онтарио до индустриализации».
Экология сообществ или синэкология — это изучение взаимодействий между видами в сообществах во многих пространственных и временных масштабах, включая распределение, структуру, численность, демографию и взаимодействие сосуществующих популяций. [1] Основное внимание экологии сообществ уделяется взаимодействиям между популяциями, определяемым конкретными генотипическими и фенотипическими характеристиками. При оценке экологии сообщества важно понимать происхождение, сохранение и последствия видового разнообразия. [2]
Экология сообщества также учитывает абиотические факторы , которые влияют на распределение или взаимодействие видов (например, годовая температура или pH почвы ). [3] Например, растительные сообщества, населяющие пустыни, сильно отличаются от растительных сообществ влажных тропических лесов из-за различий в годовом количестве осадков. Люди также могут влиять на структуру сообщества посредством нарушения среды обитания , например, интродукции инвазивных видов .
На более глубоком уровне значение и ценность концепции сообщества в экологии являются предметом дискуссий. Сообщества традиционно понимались в более широком смысле как локальные процессы, создающие (или разрушающие) совокупность видов, например, то, как изменение климата может повлиять на состав травяных сообществ. [4] Недавно такая ориентация на местное сообщество подверглась критике. Роберт Риклефс , профессор биологии в Университете Миссури и автор книги « Дезинтеграция экологического сообщества» , утверждал, что полезнее думать о сообществах в региональном масштабе, опираясь на эволюционную таксономию и биогеографию . [1] где одни виды или клады развиваются, а другие вымирают. [5] Сегодня экология сообществ фокусируется на экспериментах и математических моделях, однако раньше она фокусировалась в первую очередь на моделях организмов. Например, таксономические подразделения сообществ называются популяциями , а функциональные подразделения — гильдиями .
Организация [ править ]
Ниша [ править ]
Внутри сообщества каждый вид занимает свою нишу . Ниша вида определяет, как он взаимодействует с окружающей средой и его роль в сообществе. Имея разные ниши, виды могут сосуществовать. [6] Это известно как разделение ниши. Например, время суток, на которое охотится тот или иной вид, или на какую добычу он охотится.
Разделение ниш снижает конкуренцию между видами. [7] так что виды могут сосуществовать, поскольку они подавляют собственный рост больше, чем ограничивают рост других видов (т. е. конкуренция внутри вида больше, чем конкуренция между видами, или внутривидовая конкуренция больше, чем межвидовая).
Количество ниш, присутствующих в сообществе, определяет количество присутствующих видов. Если два вида занимают одну и ту же нишу (например, одинаковые потребности в пище), то один вид превосходит другой. Чем больше ниш заполнено, тем выше биоразнообразие сообщества.
Трофический уровень [ править ]

вида Трофический уровень — это его положение в пищевой цепи или сети. В нижней части пищевой цепи находятся автотрофы , также известные как первичные продуценты . Производители обеспечивают свою собственную энергию посредством фотосинтеза или хемосинтеза , растения являются первичными продуцентами. Следующий уровень — травоядные (основные потребители), эти виды питаются растительностью в качестве источника энергии. Травоядные животные поедаются всеядными или плотоядными животными . Эти виды являются вторичными и третичными консументами. Дополнительные уровни трофической шкалы возникают, когда более мелкие всеядные или плотоядные животные поедаются более крупными. На вершине пищевой цепи находится высший хищник , этот вид животных не потребляется другими членами сообщества. Травоядные, всеядные и плотоядные животные являются гетеротрофами . [8]
Основной пример пищевой цепи: трава → кролик → лиса. Пищевые цепи становятся более сложными, когда присутствует больше видов, часто представляющих собой пищевые сети. Энергия передается вверх по трофическим уровням. Энергия теряется на каждом уровне из-за экологической неэффективности . [9]
Трофический уровень организма может меняться в зависимости от присутствия других видов. Например, тунец может быть высшим хищником, питающимся более мелкой рыбой, такой как скумбрия. Однако в сообществе, где присутствует вид акул, акула становится высшим хищником, питающимся тунцом. [10]
Редуценты играют важную роль в трофической пирамиде. Они обеспечивают источник энергии и питательных веществ для видов растений в сообществе. Разрушители, такие как грибы и бактерии, перерабатывают энергию обратно в основу пищевой цепи, питаясь мертвыми организмами на всех трофических уровнях. [11]
Гильдия [ править ]
Гильдия — это группа видов в сообществе, которые используют одни и те же ресурсы одинаковым образом. Организмы в одной гильдии испытывают конкуренцию из-за общего ресурса. [12] Близкородственные виды часто находятся в одной гильдии из-за черт, унаследованных по общему происхождению от общего предка . Однако гильдии состоят не только из близкородственных видов. [13]
Хищники, всеядные и травоядные — все это основные примеры гильдий. Более точная гильдия - это позвоночные, которые кормятся наземными членистоногими , сюда входят определенные птицы и млекопитающие. [14] Цветковые растения, имеющие одного и того же опылителя, также образуют гильдию. [15]
виды Влиятельные
Определенные виды оказывают большее влияние на сообщество посредством прямого и косвенного взаимодействия с другими видами. На популяцию влиятельных видов влияют абиотические и биотические нарушения. Эти виды важны для определения экологических сообществ. Утрата этих видов приводит к большим изменениям в сообществе, часто снижая его стабильность. Изменение климата и интродукция инвазивных видов могут повлиять на функционирование ключевых видов и, таким образом, оказать косвенное воздействие на общественные процессы. Индустриализация и попадание химических загрязнителей в окружающую среду навсегда изменили сообщества и даже целые экосистемы. [16]
Виды фундамента [ править ]
Основные виды во многом влияют на популяцию, динамику и процессы сообщества, создавая физические изменения в самой окружающей среде. [17] Эти виды могут занимать любой трофический уровень, но, как правило, являются продуцентами. [18] Красный мангровый лес является основным видом морских сообществ. Корни мангровых зарослей служат местом нагула молоди рыб, например, окуней . [19]
Сосна белокорая ( Pinus albicaulis ) — основной вид. После пожара дерево обеспечивает тень (благодаря своему густому росту), что позволяет возобновить рост других видов растений в сообществе. Этот рост способствует возвращению беспозвоночных и микробов, необходимых для разложения. Семена белокорой сосны служат пищей медведям гризли. [20]

Краеугольные виды [ править ]
Ключевые виды оказывают непропорционально большее влияние на сообщество, чем большинство видов. Ключевые виды, как правило, находятся на более высоких трофических уровнях, часто являясь высшими хищниками. Удаление ключевых видов вызывает нисходящие трофические каскады . Волки — ключевой вид, являющийся высшим хищником.
В Йеллоустонском национальном парке потеря популяции волков из-за чрезмерной охоты привела к потере биоразнообразия в сообществе. Волки контролировали численность лосей в парке с помощью хищников. Без волков популяция лосей резко увеличилась, что привело к чрезмерному выпасу скота. Это отрицательно повлияло на другие организмы в парке; увеличение выпаса лосей лишило других присутствующих животных источников пищи. С тех пор волки были вновь введены, чтобы вернуть парковое сообщество к оптимальному функционированию. см. в разделах «Реинтродукция волков» и «История волков в Йеллоустоне» Более подробную информацию об этом тематическом исследовании .
Морским примером краеугольного вида является Pisaster ochraceus . Эта морская звезда контролирует численность Mytilus Californianus , обеспечивая достаточно ресурсов для других видов в сообществе. [21]
Инженеры-экологи [ править ]
Экосистемный инженер — это вид, который поддерживает, модифицирует и создает аспекты сообщества. Они вызывают физические изменения в среде обитания и изменяют ресурсы, доступные другим присутствующим организмам. [22]
Бобры, строящие плотины, — инженеры-экологи. Вырубая деревья для образования плотин, они изменяют поток воды в сообществе. Эти изменения влияют на растительность прибрежной зоны , исследования показывают, что биоразнообразие увеличивается. [23] Роя бобры, они создают каналы, увеличивая связи между местами обитания. Это помогает передвижению других организмов в сообществе, таких как лягушки. [24]
Теории структуры сообщества
Структура сообщества – это состав сообщества. Его часто измеряют с помощью биологических сетей , таких как пищевые сети . [25] Пищевые сети — это карта, показывающая сети видов и энергию, которая связывает виды вместе посредством трофических взаимодействий. [26]
Холистическая теория [ править ]
Холистическая теория относится к идее, что сообщество определяется взаимодействием между входящим в него организмами. Все виды взаимозависимы, каждый из них играет жизненно важную роль в работе сообщества. Благодаря этому сообщества повторяются и их легко идентифицировать, а повсюду контролируют одни и те же абиотические факторы.
Фредерик Клементс разработал целостную (или организмистическую) концепцию сообщества, как если бы оно было суперорганизмом или дискретной единицей с четкими границами. [27] Клементс предложил эту теорию, заметив, что определенные виды растений регулярно встречаются вместе в местах обитания, и пришел к выводу, что эти виды зависят друг от друга. Формирование сообществ неслучайно и предполагает коэволюцию . [28]
Холистическая теория вытекает из более широкого подхода к холизму , который относится к системе, состоящей из многих частей, и все они необходимы для функционирования системы.
Индивидуалистическая теория [ править ]
Генри Глисон разработал индивидуалистическую (также известную как открытая или континуальная) концепция сообщества, в которой численность популяции вида постепенно меняется в соответствии со сложными экологическими градиентами. [29] Каждый вид изменяется независимо по отношению к другим видам, присутствующим вдоль градиента. [30] Объединение видов случайно и обусловлено совпадением. На состав сообщества влияют различные условия окружающей среды и вероятность прибытия и закрепления каждого вида вдоль градиента. [31]
Индивидуалистическая теория предполагает, что сообщества могут существовать как непрерывные образования в дополнение к дискретным группам, упомянутым в целостной теории.
Нейтральная теория [ править ]
Стивен П. Хаббелл представил нейтральную теорию экологии (не путать с нейтральной теорией молекулярной эволюции ). Внутри сообщества (или метасообщества ) виды функционально эквивалентны, а численность популяции вида изменяется в результате случайных демографических процессов (т. е. случайных рождений и смертей). [32] Эквивалентность видов в сообществе приводит к экологическому дрейфу. Экологический дрейф приводит к случайным колебаниям популяций видов, в то время как общая численность особей в сообществе остается постоянной. Когда человек умирает, существует равная вероятность того, что каждый вид колонизирует этот участок. Стохастические изменения могут привести к вымиранию видов внутри сообщества, однако это может занять много времени, если особей этого вида много.
Виды могут сосуществовать, поскольку они схожи, а ресурсы и условия применяют фильтр к типу видов, присутствующих в сообществе. Каждая популяция имеет одинаковую адаптивную ценность (конкурентоспособность и способность к расселению) и потребность в ресурсах. Локальный и региональный состав представляет собой баланс между видообразованием или расселением (которые увеличивают разнообразие) и случайным вымиранием (которые уменьшают разнообразие). [33]
Межвидовые взаимодействия [ править ]
Виды взаимодействуют различными способами: конкуренция, хищничество , паразитизм , мутуализм , комменсализм и т. д. Организация биологического сообщества с учетом экологических взаимодействий называется структурой сообщества.
| Взаимодействия | Вид 1 | |||
|---|---|---|---|---|
| Отрицательный | Нейтральный | Позитивный | ||
| Вид 2 | Отрицательный | Соревнование | Аменсализм | Хищничество/паразитизм |
| Нейтральный | Аменсализм | Нейтрализм | Комменсализм | |
| Позитивный | Хищничество/паразитизм | Комменсализм | мутуализм | |
Конкурс [ править ]
Виды могут конкурировать друг с другом за ограниченные ресурсы . Это считается важным ограничивающим фактором размера популяции , биомассы и видового богатства . Описано множество типов конкуренции, но доказательство существования этих взаимодействий является предметом дискуссий. Прямая конкуренция наблюдалась между особями, популяциями и видами, но мало свидетельств того, что конкуренция была движущей силой эволюции больших групп. [34]
- Интерференционная конкуренция : возникает, когда особь одного вида напрямую мешает особи другого вида. Это может быть за еду или за территорию. Примеры включают в себя льва, преследующего убитую гиену, или растение, выделяющее аллелопатические химические вещества, препятствующие росту конкурирующих видов.
- Явная конкуренция : возникает, когда два вида имеют общего хищника. Например, пума охотится на лесных карибу и оленей. Популяции обоих видов могут быть подавлены хищничеством без прямой эксплуататорской конкуренции. [35]
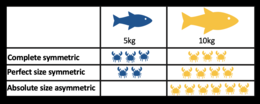
- Эксплуататорская конкуренция : происходит за счет потребления ресурсов. Когда особь одного вида потребляет ресурс (например, еду, жилье, солнечный свет и т. д.), этот ресурс больше не доступен для потребления представителем второго вида. Считается, что эксплуататорская конкуренция более распространена в природе, но необходимо проявлять осторожность, чтобы отличать ее от явной конкуренции. Примером эксплуататорской конкуренции может быть между травоядными животными, потребляющими растительность; кролик и олень едят луговую траву. Эксплуататорская конкуренция бывает разной:
- полная симметричность – все особи получают одинаковое количество ресурсов, независимо от их размера
- идеальная симметричность размеров - все особи используют одинаковое количество ресурсов на единицу биомассы.
- абсолютная асимметричность размеров — самые крупные особи используют весь доступный ресурс. [36]
- Степень асимметрии размеров оказывает большое влияние на структуру и разнообразие экологических сообществ.
Хищничество [ править ]
Хищничество охотится на другие виды ради еды. Это позитивно-негативное взаимодействие: виды-хищники получают выгоду, а виды-жертвы наносят вред. Некоторые хищники убивают свою добычу, прежде чем съесть ее, что также известно как «убить и съесть». Например, ястреб ловит и убивает мышь.Другие хищники — паразиты, питающиеся добычей при жизни, например летучая мышь-вампир, питающаяся коровой. Однако паразитизм может со временем привести к гибели организма-хозяина. Другой пример — питание растениями травоядных животных , например, пасущейся коровы. Травоядность — это тип хищничества, при котором растение (добыча в данном примере) пытается отговорить хищника от поедания растения, нагнетая токсин в листья растения. Это может привести к тому, что хищник съест другие части растения или вообще не съест растение. [37] Хищничество может повлиять на размер популяции хищников и жертв, а также на количество видов, сосуществующих в сообществе.
Хищничество может быть специализированным, например, ласка охотится исключительно на полевок. Или универсальный вариант, например, белый медведь в основном питается тюленями, но может переключиться на питание птицами, когда популяция тюленей низкая. [38] [39]
Виды могут быть одиночными или групповыми хищниками. Преимущество охоты в группе означает, что можно поймать более крупную добычу, однако источник пищи должен быть общим. Волки — групповые хищники, а тигры — одиночки.

Хищничество зависит от плотности , что часто приводит к популяционным циклам. Когда добычи много, количество видов хищников увеличивается, что приводит к поеданию большего количества видов добычи и к сокращению популяции добычи. Из-за нехватки пищи популяция хищника сокращается. Из-за отсутствия хищников популяция жертв увеличивается. см . в уравнениях Лотки – Вольтерра Более подробную информацию об этом . Хорошо известным примером этого являются циклы популяций рыси и зайца , наблюдаемые на севере. [40]
Хищничество может привести к коэволюции – эволюционной гонке вооружений , жертва приспосабливается, чтобы избегать хищника, хищник развивается. Например, у вида-жертвы вырабатывается токсин, который убивает хищника, а у хищника развивается устойчивость к токсину, что делает его более не смертельным.
Мутуализм [ править ]
Мутуализм – это взаимодействие между видами, при котором оба вида получают выгоду.
Примером могут служить бактерии Rhizobium , растущие в клубеньках на корнях бобовых растений. Эти отношения между растением и бактериями являются эндосимбиотическими : бактерии живут на корнях бобовых. Растение обеспечивает бактерии соединениями, образующимися в процессе фотосинтеза, которые можно использовать в качестве источника энергии. В то время как Rhizobium являются азотфиксирующими бактериями, обеспечивающими растение аминокислотами или аммонием. [41]
Еще одним примером являются насекомые, опыляющие цветки покрытосеменных растений . Многие растения зависят от опыления опылителями. женского Опылитель переносит пыльцу с мужского цветка на рыльце . Это оплодотворяет цветок и позволяет растению размножаться. Пчелы, такие как медоносные пчелы , являются наиболее известными опылителями. Пчелы получают нектар из растений, который они используют в качестве источника энергии. Неперенесенная пыльца обеспечивает пчелу белком. Растение получает пользу от оплодотворения, а пчела получает питание. [42]
Комменсализм [ править ]
Комменсализм – это тип отношений между организмами, при которых один организм получает пользу, а другой не получает ни пользы, ни вреда. Организм, получивший пользу, называется комменсалом , а другой организм, которому не было ни пользы, ни вреда, называется хозяином .
Например, эпифитная орхидея, прикрепленная к дереву для поддержки, приносит орхидее пользу, но не вредит и не приносит пользы дереву. Этот вид комменсализма называется инквилинизмом , орхидея постоянно живет на дереве.
Форезия - еще один тип комменсализма, комменсал использует хозяина исключительно для транспорта. Многие виды клещей для распространения полагаются на другие организмы, например, на птиц или млекопитающих. [43]
Метабиоз – конечная форма комменсализма. Комменсал полагается на хозяина, чтобы подготовить среду, пригодную для жизни. Например, у водорослей есть корневая система, называемая опорой , которая прикрепляет их к морскому дну. После укоренения он обеспечивает моллюскам , таким как морские улитки, дом, защищающий их от хищников. [44]
Аменсализм [ править ]
Противоположностью комменсализма является аменсализм , межвидовые отношения, при которых продукт одного организма оказывает негативное влияние на другой организм, но исходный организм не затрагивается. [45]
Примером может служить взаимодействие головастиков обыкновенной лягушки и пресноводной улитки . Головастики потребляют большое количество микроводорослей. Из-за того, что водорослей становится меньше для улитки, водоросли, доступные для улитки, также становятся более низкого качества. Таким образом, головастик оказывает негативное воздействие на улитку, не получая от нее заметного преимущества. Головастики получали одинаковое количество пищи независимо от присутствия улитки или без нее. [46]
Более старое и высокое дерево может препятствовать росту деревьев меньшего размера. Новый саженец, растущий в тени взрослого дерева, изо всех сил пытается получить свет для фотосинтеза. Взрослое дерево также имеет хорошо развитую корневую систему, что помогает ему конкурировать с саженцем за питательные вещества. Таким образом, рост саженца замедляется, что часто приводит к его гибели. Отношения между двумя деревьями - аменсализм: на взрослое дерево не влияет присутствие меньшего. [47]
Паразитизм [ править ]
Паразитизм – это взаимодействие, при котором один организм, хозяин, получает вред, а другой, паразит, получает пользу.
Паразитизм — это симбиоз , долговременная связь, при которой паразит питается хозяином или отбирает у него ресурсы. Паразиты могут жить в организме, например, ленточный червь . Или на поверхности тела, например, от вшей .

Малярия является результатом паразитарной связи между самкой комара Anopheles и плазмодием . Комары заражаются паразитом, питаясь зараженным позвоночным. Внутри комара плазмодий развивается в стенке средней кишки. После развития зиготы паразит перемещается в слюнные железы, где может передаваться позвоночным видам, например человеку. [48] Комар действует как переносчик малярии. Паразит имеет тенденцию сокращать продолжительность жизни комара и подавлять производство потомства. [49]
Второй пример паразитизма — выводковый паразитизм . Кукушки регулярно занимаются этим видом паразитизма. Кукушки откладывают яйца в гнезда других видов птиц. Таким образом, хозяин заботится о птенце кукушки, как о своем собственном, не замечая разницы. [50] Птенцы кукушки выбрасывают детенышей хозяина из гнезда, а это означает, что они получают более высокий уровень заботы и ресурсов от родителей. Выращивание молодняка обходится дорого и может снизить успех будущего потомства, поэтому кукушка пытается избежать этих затрат за счет выводкового паразитизма. [51]
Подобно хищничеству, паразитизм может привести к эволюционной гонке вооружений . Хозяин эволюционирует, чтобы защитить себя от паразита, а паразит развивается, чтобы преодолеть это ограничение. [52]
Нейтрализм [ править ]
Нейтрализм – это когда виды взаимодействуют, но это взаимодействие не оказывает заметного влияния ни на один из участвующих видов. Из-за взаимосвязанности сообществ настоящий нейтрализм встречается редко. Примеры нейтрализма в экологических системах трудно доказать из-за косвенного воздействия, которое виды могут оказывать друг на друга.
См. также [ править ]
- Биоценоз - взаимодействующие организмы, живущие вместе в среде обитания.
- Коэволюция – два или более вида влияют на эволюцию друг друга.
- Структура сообщества - концепция теории графов
- Конвергентная эволюция - независимая эволюция схожих функций.
- Теория сосуществования - концепция, объясняющая, как черты конкурентов могут поддерживать видовое разнообразие.
- Глубоководное сообщество — группы организмов, живущих глубоко под поверхностью моря и имеющих общую среду обитания.
- Экологические последствия биоразнообразия
- Эволюционная радиация - увеличение таксономического разнообразия или морфологического различия.
- Предельное сходство - концепция теоретической экологии и экологии сообществ.
- Метасообщество - Группа сообществ по экологии.
- Популяционная экология - Подобласть экологии
- Экология фагового сообщества - Взаимодействие бактериофагов с окружающей средой.
- Филогеография - раздел генеалогии.
- Фитоценоз - эмпирическое исследование роста растений в сообществах.
- Растительное сообщество - Коллекция местных фотосинтезирующих организмов.
- Научная классификация – Наука классификации.
- асимметричная конкуренция по размеру
- Правило R* — гипотеза о конкуренции за использование растительных ресурсов в экологии сообщества.
- Теория КСО – Теоретическая экология
- Гильдия (экология) - группа симпатрических видов со схожей экологической функцией.
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б Сахни, С.; Бентон, MJ (2008). «Восстановление после самого глубокого массового вымирания всех времен» . Труды Королевского общества B: Биологические науки . 275 (1636): 759–65. дои : 10.1098/rspb.2007.1370 . ПМЦ 2596898 . ПМИД 18198148 .
- ^ Морин, Питер Дж. (13 апреля 2009 г.). Общественная экология . Джон Уайли и сыновья. ISBN 978-1-4443-1231-7 .
- ^ Дансон, Уильям А.; Трэвис, Джозеф (ноябрь 1991 г.). «Роль абиотических факторов в общественной организации». Американский натуралист . 138 (5): 1067–1091. дои : 10.1086/285270 . S2CID 84867707 .
- ^ Грайм Дж. П.; и др. (2008). «Долгосрочное сопротивление моделируемому изменению климата на бесплодных пастбищах» . ПНАС . 105 (29): 10028–10032. Бибкод : 2008PNAS..10510028G . дои : 10.1073/pnas.0711567105 . ПМЦ 2481365 . ПМИД 18606995 .
- ^ Риклефс Р.Э. (2008). «Распад экологического сообщества». Американский натуралист . 172 (6): 741–750. дои : 10.1086/593002 . ПМИД 18954264 . S2CID 17464820 .
- ^ Альбрехт, М.; Готелли, Нью-Джерси (2001). «Пространственное и временное разделение ниш у луговых муравьев». Экология . 126 (1): 134–141. Бибкод : 2001Oecol.126..134A . дои : 10.1007/s004420000494 . ПМИД 28547432 . S2CID 5236696 .
- ^ Утомленный, Карл С.; Исон, Перри К. (2017). «Разделение ниш и роль внутривидовых вариаций ниш в структурировании гильдии бесхвостых универсалов» . Королевское общество открытой науки . 4 (3): 170060. Бибкод : 2017RSOS....470060C . дои : 10.1098/rsos.170060 . ПМЦ 5383860 . ПМИД 28405403 .
- ^ «Трофический уровень – определение, примеры и факты» . Британская энциклопедия .
- ^ Козловский, Дэниел Георгиевич (1968). «Критическая оценка концепции трофического уровня. I. Экологическая эффективность». Экология . 49 (1): 48–60. Бибкод : 1968Экол...49...48К . дои : 10.2307/1933560 . JSTOR 1933560 .
- ^ КОРТЕС, Э (1999). «Стандартизированный состав рациона и трофический уровень акул» . Журнал морских наук ICES . 56 (5): 707–717. Бибкод : 1999ICJMS..56..707C . дои : 10.1006/jmsc.1999.0489 .
- ^ Наим, Шахид; Хан, Дэниел Р.; Шурман, Грегор (2000). «Взаимозависимость производителя и разрушителя влияет на биоразнообразие». Природа . 403 (6771): 762–764. Бибкод : 2000Natur.403..762N . дои : 10.1038/35001568 . ПМИД 10693803 . S2CID 998380 .
- ^ «Цеховая экология» . Британская энциклопедия .
- ^ Корнян, Мартин; Кропиль, Рудольф (2014). «Что такое экологические гильдии? Дилемма концепций гильдий». Российский экологический журнал . 45 (5): 445–447. Бибкод : 2014RuJEc..45..445K . дои : 10.1134/S1067413614050178 . S2CID 7727306 .
- ^ Крунквист, Мэри Джо; Брукс, Роберт П. (1991). «Использование гильдий птиц и млекопитающих в качестве индикаторов кумулятивного воздействия на прибрежно-болотные территории». Экологический менеджмент . 15 (5): 701–714. Бибкод : 1991EnMan..15..701C . дои : 10.1007/BF02589628 . S2CID 55353111 .
- ^ Пеллмир, Олле; Томпсон, Джон Н. (1996). «Источники различий в вкладе опылителей внутри гильдии: влияние факторов растений и опылителей». Экология . 107 (4): 595–604. Бибкод : 1996Oecol.107..595P . дои : 10.1007/BF00333953 . ПМИД 28307405 . S2CID 26210118 .
- ^ Рор, Джейсон Р.; Керби, Джейкоб Л.; Сих, Эндрю (ноябрь 2006 г.). «Экология сообщества как основа прогнозирования воздействия загрязнений» . Тенденции в экологии и эволюции . 21 (11): 606–613. дои : 10.1016/j.tree.2006.07.002 . PMID 16843566 – через Cell Press.
- ^ «Виды, оказывающие большое влияние на структуру сообщества | Изучайте науку в Scitable» . www.nature.com . Проверено 16 февраля 2023 г.
- ^ Эллисон, Аарон М.; Банк, Майкл С.; и др. (ноябрь 2005 г.). «Потеря основных видов: последствия для структуры и динамики лесных экосистем» . Границы в экологии и окружающей среде . 3 (9): 479–486. doi : 10.1890/1540-9295(2005)003[0479:LOFSCF]2.0.CO;2 . hdl : 11603/29165 .
- ^ Анджелини, Кристина; Альтьери, Эндрю Х.; и др. (октябрь 2011 г.). «Взаимодействие между основными видами и их последствия для общественной организации, биоразнообразия и сохранения» . Бионаука . 61 (10): 782–789. дои : 10.1525/bio.2011.61.10.8 .
- ^ Эллисон, Аарон М.; Банк, Майкл С.; и др. (2005). «Потеря основных видов: последствия для структуры и динамики лесных экосистем» . Границы в экологии и окружающей среде . 3 (9): 479–486. doi : 10.1890/1540-9295(2005)003[0479:LOFSCF]2.0.CO;2 . hdl : 11603/29165 .
- ^ Менге, Брюс А.; Берлоу, Эрик Л.; и др. (1994). «Концепция ключевых видов: изменение силы взаимодействия в каменистой приливной среде обитания». Экологические монографии . 64 (3): 249–286. Бибкод : 1994ЭкоМ...64..249М . дои : 10.2307/2937163 . JSTOR 2937163 .
- ^ Джонс, Клайв Г.; Лоутон, Джон Х.; Шачак, Моше (1994). «Организмы как инженеры экосистем». Ойкос . 69 (3): 373. Бибкод : 1994Oikos..69..373J . дои : 10.2307/3545850 . JSTOR 3545850 .
- ^ Райт, Джастин П.; Джонс, Клайв Г.; Флекер, Александр С. (2002). «Инженер экосистемы, бобр, увеличивает видовое богатство в ландшафтном масштабе». Экология . 132 (1): 96–101. Бибкод : 2002Oecol.132...96W . дои : 10.1007/s00442-002-0929-1 . ПМИД 28547281 . S2CID 5940275 .
- ^ Худ, Глиннис А.; Ларсон, Дэвид Г. (2015). «Экологическая инженерия и водная связь: новый взгляд на водно-болотные угодья, модифицированные бобрами». Пресноводная биология . 60 (1): 198–208. Бибкод : 2015FrBio..60..198H . дои : 10.1111/fwb.12487 .
- ^ Эйди, Уолтер Х.; Лавленд, Карен (2007). «Структура сообщества: биоразнообразие в модельных экосистемах». Динамический Аквариум (Третье издание) . Академическая пресса: 173–189. дои : 10.1016/B978-0-12-370641-6.50021-2 . ISBN 978-0-12-370641-6 .
- ^ Томпсон, Росс М.; Брозе, Ульрих; Данн, Дженнифер А.; Холл младший, Роберт О.; Гладиз, Салли; Китчинг, Роджер Л.; Мартинес, Нео Д.; Рантала, Хайди; Романюк Тамара Н.; Стоуффер, Дэниел Б.; Тилианакис, Джейсон М. (декабрь 2012 г.). «Пищевые сети: согласование структуры и функции биоразнообразия» . Тенденции в экологии и эволюции . 27 (12): 689–697. дои : 10.1016/j.tree.2012.08.005 . hdl : 10261/67305 . PMID 22959162 – через Cell Press.
- ^ Ханспах, Ян; Хартель, Тибор; и др. (2014). «Целостный подход к изучению социально-экологических систем и его применение к южной Трансильвании» . Экология и общество . 19 (4). дои : 10.5751/ES-06915-190432 .
- ^ Шипли, Билл; Кедди, Пол А. (апрель 1987 г.). «Индивидуалистические концепции и концепции общинной единицы как фальсифицируемые гипотезы». Растительность . 69 (1–3): 47–55. дои : 10.1007/BF00038686 . S2CID 25395638 .
- ^ Верховф, Герман А. (23 мая 2012 г.). «Экология сообщества». Оксфордские библиографии . дои : 10.1093/обо/9780199830060-0042 . ISBN 978-0-19-983006-0 .
- ^ «Что такое классификация растительности?» . Международная ассоциация наук о растительности (IAVS) . Проверено 8 марта 2015 г.
- ^ Макинтош, Роберт П. (1995). «Индивидуалистическая концепция Х.А. Глисона» и теория сообществ животных: продолжающаяся полемика». Биологические обзоры . 70 (2): 317–357. дои : 10.1111/j.1469-185X.1995.tb01069.x . ПМИД 7605849 . S2CID 6328280 .
- ^ Хаббелл, Стивен П. (2001). Единая нейтральная теория биоразнообразия и биогеографии (Печать по запросу. Под ред.). Принстон [ua]: Princeton Univ. Нажимать. ISBN 978-0-691-02128-7 .
- ^ Велленд, Марк (июнь 2010 г.). «Концептуальный синтез в экологии сообществ». Ежеквартальный обзор биологии . 85 (2): 183–206. дои : 10.1086/652373 . ПМИД 20565040 . S2CID 10026873 .
- ^ Сахни, С.; Бентон, MJ; Ферри, Пенсильвания (2010). «Связь между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–547. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
- ^ Холт Р.Д. (1977). «Хищничество, очевидная конкуренция и структура сообществ добычи». Теоретическая популяционная биология . 12 (2): 197–229. дои : 10.1016/0040-5809(77)90042-9 . ПМИД 929457 .
- ^ дель Рио, Мирен; Кондес, Соня; Преч, Ганс (2014). «Анализ симметричной и асимметричной по размерам и внутри- и межвидовой конкуренции в смешанных насаждениях бука (Fagus sylvatica L.)» . Лесная экология и управление . 325 : 90–98. дои : 10.1016/j.foreco.2014.03.047 .
- ^ Леман, Кларенс; Лоберг, Шелби; Кларк, Адам Т; Шмиттер, Дэниел (22 апреля 2020 г.). «Объединение основных моделей экологии, чтобы сделать их более полными и простыми в обучении» . Бионаука . 70 (5): 415–426. doi : 10.1093/biosci/biaa013 . ISSN 0006-3568 .
- ^ Грэм, Исла М.; Ламбин, Ксавье (2002). «Влияние хищничества ласки на циклическое выживание полевок: гипотеза специализированного хищника противоречит» . Журнал экологии животных . 71 (6): 946–956. Бибкод : 2002JAnEc..71..946G . дои : 10.1046/j.1365-2656.2002.00657.x .
- ^ Рассел, Ричард Х. (1975). «Пищевые привычки белых медведей залива Джеймс и юго-западной части Гудзонова залива летом и осенью» . Арктика . 28 (2). дои : 10.14430/arctic2823 .
- ^ Кейт, Ллойд Б. (1983). «Роль еды в циклах популяции зайцев». Ойкос . 40 (3): 385–395. Бибкод : 1983Ойкос..40..385К . дои : 10.2307/3544311 . JSTOR 3544311 .
- ^ Мароти, Гергеи; Кондороси, Ева (2014). «Азотфиксирующий симбиоз ризобий-бобовых растений: являются ли полиплоидия и дифференцировка симбионтов, управляемая пептидами хозяина, общими принципами эндосимбиоза?» . Границы микробиологии . 5 : 326. дои : 10.3389/fmicb.2014.00326 . ПМК 4074912 . ПМИД 25071739 .
- ^ Хунг, Кенг-Лу Джеймс; Кингстон, Дженнифер М.; и др. (2018). «Всемирное значение медоносных пчел как опылителей в естественной среде обитания» . Труды Королевского общества B: Биологические науки . 285 (1870): 20172140. doi : 10.1098/rspb.2017.2140 . ПМК 5784195 . ПМИД 29321298 .
- ^ Хоук, Массачусетс; ОКоннор, Б.М. (январь 1991 г.). «Экологическое и эволюционное значение форезии астигматов». Ежегодный обзор энтомологии . 36 (1): 611–636. дои : 10.1146/annurev.en.36.010191.003143 .
- ^ Андерсон, Марти Дж .; Дибель, Кэрол Э.; и др. (2005). «Согласованность и изменчивость скоплений водорослей: пространственные закономерности биоразнообразия основных типов при различных таксономических разрешениях». Журнал экспериментальной морской биологии и экологии . 320 : 35–56. дои : 10.1016/j.jembe.2004.12.023 .
- ^ Уилли, Джоан М.; Шервуд, Линда М.; Вулвертон Кристофер Дж. (2011). Микробиология . Прескотта. стр. 713–738.
- ^ Доддс, Уолтер К.; Пока, Мэтт Р. (2020). «Мирвидовые межвидовые взаимодействия растений и животных в пресноводных сообществах». Экология пресноводных вод (3-е изд.). Эльзевир. стр. 653–670. дои : 10.1016/b978-0-12-813255-5.00021-1 . ISBN 978-0-12-813255-5 . S2CID 243070121 .
- ^ Мейер Элиан С., Элиан С.; Киенаст, Феликс; Пирман, Питер Б; Свеннинг, Йенс-Кристиан; Тюллер, Вильфрид; Араужо, Мигель Б.; Антуан, Гизан; Циммерманн, Никлаус Э. (2010). «Биотические и абиотические переменные не имеют большого дублирования в объяснении распределения видов деревьев». Экография . 33 (6): 1038–1048. Бибкод : 2010Экогр..33.1038М . дои : 10.1111/j.1600-0587.2010.06229.x .
- ^ Бейер, Джон К. (1998). «Развитие малярийных паразитов у комаров». Ежегодный обзор энтомологии . 43 : 519–543. дои : 10.1146/annurev.ento.43.1.519 . ПМИД 9444756 .
- ^ ХОГГ, ДЖОН К.; ХЕРД, ХИЛАРИ (1995). «Вызванное малярией снижение плодовитости во время первого гонотрофного цикла комаров Anopheles Stephensi». Медицинская и ветеринарная энтомология . 9 (2): 176–180. дои : 10.1111/j.1365-2915.1995.tb00175.x . ПМИД 7787226 . S2CID 30277109 .
- ^ Дэвис, Северная Каролина; Бурк, Эндрю Ф.Г.; де Л. Брук, М. (1989). «Кукушки и муравьи-паразиты: межвидовой паразитизм выводков как эволюционная гонка вооружений». Тенденции в экологии и эволюции . 4 (9): 274–278. дои : 10.1016/0169-5347(89)90202-4 . ПМИД 21227369 .
- ^ Петри, М.; Мёллер, AP (1991). «Откладка яиц в чужие гнезда: Внутривидовой выводковый паразитизм у птиц». Тенденции в экологии и эволюции . 6 (10): 315–320. дои : 10.1016/0169-5347(91)90038-Y . ПМИД 21232496 .
- ^ Шит, Дэнни Дж.; Дик, Джейми Т.А.; и др. (2018). «Победа в гонке вооружений: общая эволюционная история хозяина и паразита снижает риск заражения у рыб-последних хозяев» . Письма по биологии . 14 (7): 20180363. doi : 10.1098/rsbl.2018.0363 . ПМК 6083226 . ПМИД 30045905 .
Дальнейшее чтение [ править ]
- Акин, Уоллес Э. (1991). Глобальные закономерности: климат, растительность и почвы . Университет Оклахомы Пресс. ISBN 0-8061-2309-5 .
- Барбур, Берк и Питтс, 1987. Экология наземных растений , 2-е изд. Каммингс, Менло-Парк, Калифорния.
- Морин, Питер Дж. (1999). Общественная экология . Уайли-Блэквелл Пресс. ISBN 978-0-86542-350-3 .
- Одум Е.П. (1959) Основы экологии . WB Saunders Co., Филадельфия и Лондон.
- Риклефс, Р.Э. (2005) Экономика природы , 6-е изд. У.Х. Фриман, США.
- Рикеттс, Тейлор Х., Эрик Динерштейн, Дэвид М. Олсон, Колби Дж. Лукс и др. ( ВВФ ) (1999). Наземные экорегионы Северной Америки: оценка сохранения . Остров Пресс. ISBN 1-55963-722-6 .
- Конспект лекций курса Стефано Аллесины по экологии сообщества: https://stefanoallesina.github.io/Theoretical_Community_Ecology/
Внешние ссылки [ править ]
- Сообщество, BioMineWiki. Архивировано 27 июня 2021 г. в Wayback Machine.
- Определите виды микробов в сообществе, BioMineWiki. Архивировано 30 июня 2021 года в Wayback Machine.
- Глоссарий , состояние и тенденции национальных биологических ресурсов, Геологическая служба США.
- Глоссарий , Консультанты по охране окружающей среды ENTRIX.
