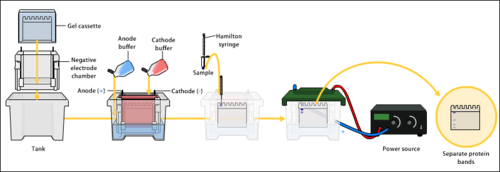
Гель-электрофорез
 Аппарат для гель-электрофореза - в этот заполненный буфером ящик помещается агарозный гель , и через источник питания сзади подается электрический ток. Отрицательная клемма находится на дальнем конце (черный провод), поэтому ДНК мигрирует к положительно заряженному аноду (красный провод). Это происходит потому, что фосфатные группы, обнаруженные во фрагментах ДНК, обладают отрицательным зарядом, который отталкивается отрицательно заряженным катодом и притягивается к положительно заряженному аноду. | |
| Классификация | Электрофорез |
|---|---|
| Другие методы | |
| Связанный | Капиллярный электрофорез SDS-СТРАНИЦА Двумерный гель-электрофорез Гель-электрофорез в градиенте температуры |

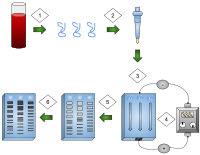
- ДНК выделяют.
- Выделение и амплификация ДНК.
- ДНК добавляли в лунки геля.
- Электрический ток подается на гель.
- Полосы ДНК разделены по размеру.
- Полосы ДНК окрашены.
Гель-электрофорез — метод разделения и анализа биомакромолекул ( ДНК , РНК , белков и т. д.) и их фрагментов на основе их размера и заряда. Он используется в клинической химии для разделения белков по заряду или размеру (агароза IEF, по существу независимая от размера), а также в биохимии и молекулярной биологии для разделения смешанной популяции фрагментов ДНК и РНК по длине, для оценки размера фрагментов ДНК и РНК или разделить белки по заряду. [1]
Молекулы нуклеиновой кислоты разделяются путем применения электрического поля для перемещения отрицательно заряженных молекул через матрицу из агарозы или других веществ. Более короткие молекулы движутся быстрее и мигрируют дальше, чем более длинные, потому что более короткие молекулы легче мигрируют через поры геля. Это явление называется просеиванием. [2] Белки разделены зарядом в агарозе, поскольку поры геля слишком велики, чтобы просеивать белки. Гель-электрофорез также можно использовать для разделения наночастиц .
Гель-электрофорез использует гель в качестве противоконвективной среды или просеивающей среды во время электрофореза, движения заряженной частицы в электрическом токе. Гели подавляют тепловую конвекцию, вызванную применением электрического поля, а также могут действовать как просеивающая среда, замедляя прохождение молекул; гели также могут просто служить для поддержания окончательного разделения, чтобы можно было нанести пятно после электрофореза. [3] Гель-электрофорез ДНК обычно проводится в аналитических целях, часто после амплификации ДНК с помощью полимеразной цепной реакции (ПЦР), но может использоваться в качестве препаративного метода перед использованием других методов, таких как масс-спектрометрия , ПДРФ , ПЦР, клонирование , секвенирование ДНК. или Саузерн-блоттинг для дальнейшей характеристики.
Физическая основа [ править ]
Электрофорез — это процесс, который позволяет сортировать молекулы по заряду, размеру или форме. Используя электрическое поле, можно заставить молекулы (например, ДНК) двигаться через гель, сделанный из агарозы или полиакриламида . Электрическое поле состоит из отрицательного заряда на одном конце, который толкает молекулы через гель, и положительного заряда на другом конце, который тянет молекулы через гель. Сортируемые молекулы распределяются в лунки в гелеобразном материале. Гель помещают в камеру для электрофореза, которую затем подключают к источнику питания. При приложении электрического поля более крупные молекулы движутся через гель медленнее, а более мелкие молекулы движутся быстрее. Молекулы разного размера образуют на геле отдельные полосы. [4]
Термин « гель » в данном случае относится к матрице, используемой для удержания и последующего разделения целевых молекул. В большинстве случаев гель представляет собой сшитый полимер , состав и пористость которого выбираются исходя из удельного веса и состава анализируемой мишени. При разделении белков или небольших нуклеиновых кислот ( ДНК , РНК или олигонуклеотидов ) гель обычно состоит из различных концентраций акриламида и сшивающего агента , образуя сетчатые сетки полиакриламида разного размера. При разделении более крупных нуклеиновых кислот (более нескольких сотен оснований ) предпочтительной матрицей является очищенная агароза. В обоих случаях гель образует твердую, но пористую матрицу. Акриламид, в отличие от полиакриламида, является нейротоксином , и с ним необходимо обращаться с соблюдением соответствующих мер предосторожности во избежание отравления. Агароза состоит из длинных неразветвленных цепей незаряженных углеводов без поперечных связей, в результате чего образуется гель с большими порами, позволяющий разделять макромолекулы и макромолекулярные комплексы. . [5]
Электрофорез относится к электродвижущей силе (ЭДС), которая используется для перемещения молекул через матрицу геля. Поместив молекулы в лунки геля и приложив электрическое поле, молекулы будут двигаться через матрицу с разной скоростью, определяемой в основном их массой, когда отношение заряда к массе (Z) всех видов одинаково. Однако, когда заряды не все однородны, электрическое поле, создаваемое процедурой электрофореза, заставит молекулы мигрировать дифференциально в зависимости от заряда. Виды, которые имеют чистый положительный заряд, будут мигрировать к катоду , который заряжен отрицательно (поскольку это электролитический, а не гальванический элемент ), тогда как виды, которые имеют чистый отрицательный заряд, будут мигрировать к положительно заряженному аноду. Масса остается фактором, определяющим скорость, с которой эти неоднородно заряженные молекулы мигрируют через матрицу к соответствующим электродам. [6]
Если несколько образцов были загружены в соседние лунки геля, они будут проходить параллельно по отдельным дорожкам. В зависимости от количества различных молекул каждая полоса показывает разделение компонентов из исходной смеси в виде одной или нескольких отдельных полос, по одной полосе на каждый компонент. Неполное разделение компонентов может привести к перекрытию полос или неразличимым размазкам, представляющим несколько неразрешенных компонентов. [ нужна ссылка ] Полосы на разных дорожках, которые оказываются на одинаковом расстоянии от вершины, содержат молекулы, прошедшие через гель с одинаковой скоростью, что обычно означает, что они примерно одинакового размера. Существуют маркеры размера молекулярной массы , которые содержат смесь молекул известных размеров. Если такой маркер был нанесен на одну дорожку в геле параллельно неизвестным образцам, наблюдаемые полосы можно сравнить с полосами неизвестных, чтобы определить их размер. Расстояние, которое проходит полоса, примерно обратно пропорционально логарифму размера молекулы (альтернативно это можно сказать, что пройденное расстояние обратно пропорционально логарифму молекулярной массы образцов). [7]
Существуют ограничения для электрофоретических методов. Поскольку прохождение тока через гель вызывает нагревание, гели могут плавиться во время электрофореза. Электрофорез проводится в буферных растворах, чтобы уменьшить изменения pH из-за электрического поля, что важно, поскольку заряд ДНК и РНК зависит от pH, но слишком долгая работа может истощить буферную емкость раствора. Существуют также ограничения при определении молекулярной массы с помощью SDS-PAGE, особенно при попытке определить молекулярную массу неизвестного белка. Некоторые биологические переменные трудно или невозможно минимизировать, и они могут влиять на электрофоретическую миграцию. К таким факторам относятся структура белка, посттрансляционные модификации и аминокислотный состав. Например, тропомиозин представляет собой кислый белок, который аномально мигрирует в гелях SDS-PAGE. Это связано с тем, что кислотные остатки отталкиваются отрицательно заряженным ДСН, что приводит к неточному соотношению массы к заряду и миграции. [8] Кроме того, разные препараты генетического материала могут не мигрировать последовательно друг с другом по морфологическим или другим причинам.
Виды геля [ править ]
Чаще всего используются агарозные и полиакриламидные гели. Каждый тип геля хорошо подходит для разных типов и размеров аналита. Полиакриламидные гели обычно используются для белков и обладают очень высокой разрешающей способностью для небольших фрагментов ДНК (5–500 п.н.). С другой стороны, агарозные гели имеют более низкую разрешающую способность для ДНК, но имеют больший диапазон разделения и поэтому используются для фрагментов ДНК размером обычно 50–20 000 п.н., но разрешение более 6 МБ возможно с помощью импульсного поля. гель-электрофорез (ПФГЭ). [9] Полиакриламидные гели работают в вертикальной конфигурации, тогда как агарозные гели обычно работают горизонтально в подводном режиме. Они также различаются методом отливки, поскольку агароза отверждается термически, а полиакриламид образуется в результате химической реакции полимеризации.
Агароза [ править ]

Агарозные гели изготавливаются из природных полисахаридов полимеров , извлеченных из морских водорослей .Агарозные гели легко отливать и обрабатывать по сравнению с другими матрицами, поскольку затвердевание геля представляет собой физическое, а не химическое изменение. Образцы также легко восстанавливаются. После окончания эксперимента полученный гель можно хранить в полиэтиленовом пакете в холодильнике.
Агарозные гели не имеют одинакового размера пор, но оптимальны для электрофореза белков размером более 200 кДа. [10] Электрофорез в агарозном геле также можно использовать для разделения фрагментов ДНК размером от 50 пар оснований до нескольких мегабаз (миллионов оснований). [11] самые крупные из которых требуют специального оборудования. Расстояние между полосами ДНК разной длины зависит от процентного содержания агарозы в геле, причем более высокие проценты требуют более длительного времени анализа, иногда нескольких дней. Вместо этого с высокопроцентными агарозными гелями следует работать с помощью импульсного полевого электрофореза (PFE) или полевого инверсионного электрофореза .
«Большинство агарозных гелей состоят из 0,7% (хорошее разделение или разрешение больших фрагментов ДНК размером 5–10 КБ) и 2% (хорошее разрешение для небольших фрагментов 0,2–1 КБ) агарозы, растворенной в буфере для электрофореза. Для электрофореза можно использовать до 3% разделяют очень мелкие фрагменты, но в этом случае более подходит вертикальный полиакриламидный гель. Гели с низким процентом очень слабы и могут сломаться при попытке их поднять. Гели с высоким процентом часто бывают хрупкими и не застывают равномерно. множество приложений». [12]
Полиакриламид [ править ]
Электрофорез в полиакриламидном геле (ПААГ) используется для разделения белков размером от 5 до 2000 кДа благодаря однородному размеру пор, обеспечиваемому полиакриламидным гелем. Размер пор контролируется путем регулирования концентрации порошка акриламида и бис-акриламида, используемого при создании геля. При создании этого типа геля необходимо соблюдать осторожность, поскольку акриламид в жидкой и порошкообразной форме является мощным нейротоксином.
Традиционные методы секвенирования ДНК, такие как методы Максама-Гилберта или Сэнгера, использовали полиакриламидные гели для разделения фрагментов ДНК, различающихся по длине одной парой оснований, чтобы последовательность можно было прочитать. В большинстве современных методов разделения ДНК сейчас используются агарозные гели, за исключением особо мелких фрагментов ДНК. В настоящее время он чаще всего используется в области иммунологии и анализа белков, часто используется для разделения разных белков или изоформ одного и того же белка на отдельные полосы. Их можно перенести на мембрану из нитроцеллюлозы или ПВДФ для исследования антителами и соответствующими маркерами, например, с помощью вестерн-блоттинга .
Обычно разрешающие гели изготавливаются 6%, 8%, 10%, 12% или 15%. Укладочный гель (5%) наливается поверх растворяющегося геля и вставляется гелевая гребенка (которая образует лунки и определяет дорожки, куда будут помещены белки, буфер для образцов и лестницы). Выбранный процент зависит от размера белка, который необходимо идентифицировать или исследовать в образце. Чем меньше известный вес, тем выше процент, который следует использовать. Изменения в буферной системе геля могут способствовать дальнейшему расщеплению белков очень малых размеров. [13]
Крахмал [ править ]
Частично гидролизованный картофельный крахмал является еще одной нетоксичной средой для электрофореза белков. Гели немного более непрозрачны, чем акриламид или агароза. Неденатурированные белки можно разделить по заряду и размеру. Их визуализируют с помощью окрашивания Napthal Black или Amido Black. Типичные концентрации крахмального геля составляют от 5% до 10%. [14] [15] [16]
Условия геля [ править ]
Денатурация [ править ]
Денатурирующие гели работают в условиях, которые нарушают естественную структуру аналита, заставляя его разворачиваться в линейную цепочку. Таким образом, подвижность каждой макромолекулы зависит только от ее линейной длины и отношения массы к заряду. Таким образом, вторичный, третичный и четверичный уровни биомолекулярной структуры разрушаются, и для анализа остается только первичная структура.
Нуклеиновые кислоты часто денатурируются путем включения мочевины в буфер, тогда как белки денатурируются с использованием додецилсульфата натрия , обычно как часть процесса SDS-PAGE . Для полной денатурации белков необходимо также восстановить ковалентные дисульфидные связи , стабилизирующие их третичную и четвертичную структуру , метод, называемый восстанавливающим ПААГ. Восстановительные условия обычно поддерживают добавлением бета-меркаптоэтанола или дитиотреитола . Для общего анализа образцов белка восстанавливающий PAGE является наиболее распространенной формой электрофореза белков .
Условия денатурации необходимы для правильной оценки молекулярной массы РНК. РНК способна образовывать больше внутримолекулярных взаимодействий, чем ДНК, что может приводить к изменению ее электрофоретической подвижности . Мочевина , ДМСО и глиоксаль являются наиболее часто используемыми денатурирующими агентами для разрушения структуры РНК. Первоначально высокотоксичный гидроксид метилртути часто использовался при электрофорезе денатурирующей РНК. [17] но для некоторых образцов этот метод может быть предпочтительным. [18]
Денатурирующий гель-электрофорез используется в методах, основанных на шаблоне полос ДНК и РНК, гель-электрофореза в температурном градиенте (TGGE). [19] и денатурирующий градиентный гель-электрофорез (DGGE). [20]
Родной [ править ]
Нативные гели работают в неденатурирующих условиях, чтобы сохранить природную структуру аналита. Это позволяет физическому размеру сложенного или собранного комплекса влиять на подвижность, позволяя анализировать все четыре уровня биомолекулярной структуры. Для биологических образцов детергенты используются только в той степени, в которой они необходимы для липидных мембран клетки лизирования . Комплексы остаются — по большей части — связанными и свернутыми, как если бы они были в клетке. Однако недостатком является то, что комплексы не могут разделяться четко и предсказуемо, поскольку трудно предсказать, как форма и размер молекулы повлияют на ее подвижность. Обращение и решение этой проблемы является основной целью препаративной нативной PAGE .
В отличие от методов денатурации, в нативном гель-электрофорезе не используется заряженный денатурирующий агент. Таким образом, разделяемые молекулы (обычно белки или нуклеиновые кислоты ) различаются не только по молекулярной массе и собственному заряду, но и по площади поперечного сечения и, таким образом, испытывают различные электрофоретические силы, зависящие от формы общей структуры. Что касается белков, поскольку они остаются в нативном состоянии, их можно визуализировать не только с помощью обычных реагентов для окрашивания белков, но также с помощью специфического ферментативного окрашивания.
Конкретным экспериментальным примером применения нативного гель-электрофореза является проверка ферментативной активности для проверки присутствия фермента в образце во время очистки белка. Например, для протеин-щелочной фосфатазы окрашивающий раствор представляет собой смесь соли 4-хлор-2-2-метилбензолдиазония с 3-фосфо-2-нафтойной кислотой-2'-4'-диметиланилином в Трис-буфере. Эта морилка продается в продаже в виде набора для окрашивания гелей. При наличии белка механизм реакции протекает в следующем порядке: начинается с дефосфорилирования 3-фосфо-2-нафтойной кислоты-2'-4'-диметиланилина щелочной фосфатазой (необходима вода для реакции). Фосфатная группа высвобождается и заменяется спиртовой группой из воды. Электрофил 4-хлор-2-2-метилбензолдиазоний (диазониевая соль Fast Red TR) замещает спиртовую группу, образуя конечный продукт - красный азокраситель. Как следует из названия, это конечный видимый красный продукт реакции. В студенческих академических экспериментах по очистке белка гель обычно используют рядом с коммерческими очищенными образцами, чтобы визуализировать результаты и сделать вывод, была ли очистка успешной. [22]
Нативный гель-электрофорез обычно используется в протеомике и металломике . Однако нативный PAGE также используется для сканирования генов (ДНК) на предмет неизвестных мутаций, как при одноцепочечном конформационном полиморфизме .
Буферы [ править ]
Буферы при гель-электрофорезе используются для получения ионов, переносящих ток, и для поддержания относительно постоянного значения pH.В этих буферах содержится много ионов, необходимых для прохождения через них электричества. Такие вещества, как дистиллированная вода или бензол, содержат мало ионов, что не идеально для использования в электрофорезе. [23] Для электрофореза используется ряд буферов. Наиболее распространенными являются нуклеиновые кислоты Трис/ацетат/ЭДТА (ТАЕ), Трис/борат/ЭДТА (ТБЕ). Было предложено множество других буферов, например, борат лития , который используется редко, на основании цитат в Pubmed (LB), изоэлектрический гистидин, буферы, соответствующие pK-товарам, и т. д.; в большинстве случаев предполагаемым объяснением является более низкая токовая (меньшая тепловая) подвижность ионов, согласованная с ним, что приводит к увеличению срока службы буфера. Борат проблематичен; Борат может полимеризоваться или взаимодействовать с цис-диолами, такими как те, которые содержатся в РНК. TAE имеет самую низкую буферную емкость, но обеспечивает лучшее разрешение для более крупных ДНК. Это означает более низкое напряжение и больше времени, но лучший продукт. LB является относительно новым и неэффективен при разрешении фрагментов размером более 5 кб; Однако из-за его низкой проводимости можно использовать гораздо более высокое напряжение (до 35 В/см), что означает более короткое время анализа для обычного электрофореза. Разница в размерах всего в одну пару оснований может быть устранена в 3% агарозном геле со средой с чрезвычайно низкой проводимостью (1 мМ борат лития). [24]
Большинство разделений белков с помощью SDS-PAGE выполняется с использованием «прерывистой» (или DISC) буферной системы , которая значительно повышает четкость полос внутри геля. Во время электрофореза в прерывистой гелевой системе на ранней стадии электрофореза формируется ионный градиент, который заставляет все белки фокусироваться на одной резкой полосе в процессе, называемом изотахофорезом . Разделение белков по размерам достигается в нижней, «рассасывающейся» области геля. Рассасывающийся гель обычно имеет гораздо меньший размер пор, что приводит к эффекту просеивания, который теперь определяет электрофоретическую подвижность белков.
Визуализация [ править ]
После завершения электрофореза молекулы геля можно окрасить , чтобы сделать их видимыми. ДНК можно визуализировать с помощью бромистого этидия , который при интеркаляции в ДНК флуоресцирует в ультрафиолетовом свете, а белок можно визуализировать с помощью окраски серебром или красителя Кумасси бриллиантовый синий . Для визуализации разделения компонентов смеси на геле можно использовать и другие методы. Если молекулы, которые необходимо разделить, содержат радиоактивность , например, в геле для секвенирования ДНК , можно записать авторадиограмму геля. систему Гели можно фотографировать, часто используя Gel Doc . Затем гели обычно маркируются для презентаций и научных записей на популярном веб-сайте по созданию фигурок SciUGo .
Последующая обработка [ править ]
После разделения можно использовать дополнительный метод разделения, такой как изоэлектрофокусирование или SDS-PAGE . Затем гель физически разрезают и белковые комплексы экстрагируют из каждой порции отдельно. Затем каждый экстракт можно проанализировать, например, с помощью дактилоскопии пептидной массы или секвенирования пептидов de novo после расщепления в геле . Это может предоставить много информации об идентичности белков в комплексе.
Приложения [ править ]
- Оценка размера молекул ДНК после расщепления рестриктазами, например, при рестрикционном картировании клонированной ДНК.
- Анализ продуктов ПЦР , например, при молекулярно -генетической диагностике или генетическом дактилоскопировании.
- Разделение ограниченной геномной ДНК перед переносом по Саузерну или РНК перед переносом по Нозерну .
Гель-электрофорез применяется в криминалистике , молекулярной биологии , генетике , микробиологии и биохимии . Результаты можно проанализировать количественно, визуализируя гель с помощью УФ-света и устройства для визуализации геля. Изображение записывается камерой с компьютерным управлением, затем измеряется интенсивность полосы или интересующего пятна и сравнивается со стандартом или маркерами, нанесенными на тот же гель. Измерения и анализ в основном выполняются с помощью специализированного программного обеспечения.
В зависимости от типа выполняемого анализа в сочетании с результатами гель-электрофореза часто применяются другие методы, что обеспечивает широкий спектр приложений в конкретной области.
Нуклеиновые кислоты [ править ]
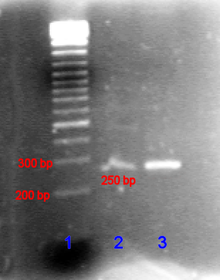
В случае нуклеиновых кислот направление миграции от отрицательных электродов к положительным обусловлено естественным отрицательным зарядом, переносимым остовом сахарофосфатным их . [25]
Фрагменты двухцепочечной ДНК естественным образом ведут себя как длинные палочки, поэтому их миграция через гель зависит от их размера или, для циклических фрагментов, от их радиуса вращения . Однако кольцевая ДНК, такая как плазмиды , может иметь несколько полос, и скорость миграции может зависеть от того, расслаблена ли она или сверхскручена. Одноцепочечная ДНК или РНК имеет тенденцию сворачиваться в молекулы сложной формы и мигрировать через гель сложным образом в зависимости от их третичной структуры. Поэтому агенты, разрушающие водородные связи , такие как гидроксид натрия или формамид , используются для денатурации нуклеиновых кислот и заставляют их снова вести себя как длинные палочки. [26]
Гель-электрофорез крупных ДНК или РНК обычно проводят с помощью электрофореза в агарозном геле. См. страницу « Метод обрыва цепи », где приведен пример полиакриламидного геля для секвенирования ДНК. Характеристика посредством лигандного взаимодействия нуклеиновых кислот или фрагментов может быть выполнена с помощью аффинного электрофореза со сдвигом подвижности .
Электрофорез образцов РНК можно использовать для проверки загрязнения геномной ДНК, а также деградации РНК. РНК эукариотических организмов демонстрирует отчетливые полосы 28-й и 18-й рРНК, причем полоса 28-х примерно в два раза интенсивнее полосы 18-х. Деградированная РНК имеет менее четко выраженные полосы, имеет размытый вид, соотношение интенсивностей менее 2:1.
Белки [ править ]
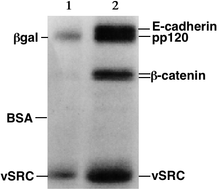
Белки , в отличие от нуклеиновых кислот, могут иметь разные заряды и сложную форму, поэтому они не могут мигрировать в полиакриламидный гель с одинаковой скоростью или со всеми одинаковыми скоростями при воздействии на образец ЭДС от отрицательной до положительной. Поэтому белки обычно денатурируются в присутствии детергента, такого как додецилсульфат натрия (SDS), который покрывает белки отрицательным зарядом. [3] Как правило, количество связанного ДСН зависит от размера белка (обычно 1,4 г ДСН на грамм белка), так что полученные денатурированные белки имеют общий отрицательный заряд, и все белки имеют одинаковое соотношение заряда к массе. соотношение. Поскольку денатурированные белки действуют как длинные палочки, а не имеют сложную третичную форму, скорость, с которой полученные белки, покрытые SDS, мигрируют в геле, зависит только от их размера, а не от их заряда или формы. [3]
Белки обычно анализируются с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия ( SDS-PAGE ), с помощью нативного гель-электрофореза , с помощью препаративного нативного гель-электрофореза ( QPNC-PAGE ) или с помощью 2-D электрофореза .
Характеристика посредством взаимодействия лигандов может быть выполнена с помощью электроблоттинга или с помощью аффинного электрофореза в агарозе или с помощью капиллярного электрофореза, что касается оценки констант связывания и определения структурных особенностей, таких как гликанов содержание посредством связывания лектина .
Наночастицы [ править ]
Новым применением гель-электрофореза является разделение или определение характеристик наночастиц металлов или оксидов металлов (например, Au, Ag, ZnO, SiO2) в отношении размера, формы или химического состава поверхности наночастиц. [27] Задача состоит в том, чтобы получить более однородный образец (например, с более узким гранулометрическим составом), который затем можно будет использовать в дальнейших продуктах/процессах (например, в процессах самосборки). Для разделения наночастиц внутри геля ключевым параметром является отношение размера частиц к размеру сетки, в результате чего были идентифицированы два механизма миграции: неограниченный механизм, где размер частиц << размер сетки, и ограниченный механизм, где Размер частиц аналогичен размеру сетки. [28]
История [ править ]
- 1930-е годы – первые сообщения об использовании сахарозы для гель-электрофореза; электрофорез с подвижной границей ( Тизелиус )
- 1950 г. – внедрение «зонального электрофореза» (Тизелиус); бумажный электрофорез
- 1955 — внедрение крахмальных гелей, посредственное разделение ( Кузницы ). [15]
- 1959 г. – внедрение акриламидных гелей; прерывистый электрофорез ( Орнштейн и Дэвис ); точный контроль таких параметров, как размер пор и стабильность; и ( Раймонд и Вайнтрауб )
- 1965 — внедрение электрофореза в свободном потоке ( Ханниг ).
- 1966 г. – первое использование агаровых гелей. [29]
- 1969 – внедрение денатурирующих агентов, особенно SDS для разделения белковых субъединиц ( Вебер и Осборн ). [30]
- 1970 – Лэммли разделил 28 компонентов фага Т4 с помощью накопительного геля и SDS.
- 1972 г. – агарозные гели с окраской бромистым этидием. [31]
- 1975 — 2-мерные гели ( О’Фаррелл ); изоэлектрофокусирование , затем гель-электрофорез в ДСН.
- 1977 — для секвенирования гели ( Sanger )
- 1981 г. – внедрение капиллярного электрофореза (Йоргенсон и Лукач).
- 1983 — гель-электрофорез в импульсном поле позволяет разделять большие молекулы ДНК ( Суили ).
- 2004 г. – введение стандартизированного времени полимеризации акриламидного геля растворов для оптимизации свойств геля, в частности стабильности геля, во время электрофореза ( Кастенхольц ). [32]
В книге Милана Бира по электрофорезу 1959 года цитируются ссылки из 1800-х годов. [33] Однако Оливер Смитис внес значительный вклад. Бир утверждает: «Метод Кузнеца... находит широкое применение благодаря своей уникальной разделительной способности». В контексте Бир явно подразумевает, что метод Смитиса является улучшением.
См. также [ править ]
Ссылки [ править ]
- ^ Крындушкин Д.С.; Александров И.М.; Тер-Аванесян; Кушниров В.В. (2003). «Агрегаты прионов дрожжей [PSI+] образованы небольшими полимерами Sup35, фрагментированными Hsp104» . J Биол Хим . 278 (49): 49636–43. дои : 10.1074/jbc.M307996200 . ПМИД 14507919 .
- ^ Сэмбрук, Джозеф (2001). Молекулярное клонирование: лабораторное руководство (на испанском языке). Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. ISBN 978-0-87969-576-7 . OCLC 45015638 .
- ^ Jump up to: Перейти обратно: а б с Берг, Джереми (2002). Биохимия (на эстонском языке). Нью-Йорк: WH Freeman. ISBN 978-0-7167-4955-4 . OCLC 48055706 .
- ^ Уилсон, Кейт (2018). Принципы и методы биохимии и молекулярной биологии Уилсона и Уокера . Кембридж, Великобритания Нью-Йорк, Нью-Йорк: Издательство Кембриджского университета. ISBN 978-1-316-61476-1 . OCLC 998750377 .
- ^ Бойер, Родни (2000). Современная экспериментальная биохимия (на эстонском языке). Сан-Франциско: Бенджамин Каммингс. ISBN 978-0-8053-3111-0 . OCLC 44493241 .
- ^ Робит, Джон (1990). Биохимические методы: теория и практика . Проспект-Хайтс, Иллинойс: Waveland Press. ISBN 978-0-88133-556-9 . ОСЛК 22549624 .
- ^ Ли П.И.; Костумбрадо Дж; Сюй CY; Ким Ю.Х. (2012). «Электрофорез в агарозном геле для разделения фрагментов ДНК» . Джей Вис Эксп (62). дои : 10.3791/3923 . ПМЦ 4846332 . ПМИД 22546956 .
- ^ «Определение молекулярной массы с помощью SDS-PAGE, версия B» (PDF) . www.bio-rad.com . Архивировано (PDF) из оригинала 17 ноября 2021 года . Проверено 23 марта 2022 г.
- ^ Том Маниатис; Э. Ф. Фрич; Джозеф Сэмбрук (1982). «Глава 5, протокол 1». Молекулярное клонирование. Лабораторное пособие . Том. 1 (3-е изд.). п. 5.2–5.3. ISBN 978-0879691363 .
- ^ Смисек, Дэвид Л.; Хоугланд, Дэвид А. (1989). «Электрофорез в агарозном геле высокомолекулярных синтетических полиэлектролитов». Макромолекулы . 22 (5). Американское химическое общество (ACS): 2270–2277. Бибкод : 1989МаМол..22.2270С . дои : 10.1021/ma00195a048 . ISSN 0024-9297 .
- ^ Войтас, Дэниел (май 2001 г.). «Электрофорез в агарозном геле» . Современные протоколы в иммунологии . Глава 10: 10.4.1–10.4.8. дои : 10.1002/0471142735.im1004s02 . ISSN 1934-368X . ПМИД 18432695 . S2CID 39623776 . Архивировано из оригинала 2 февраля 2022 года . Проверено 1 марта 2023 г.
- ^ «Электрофорез в агарозном геле (основной метод)» . Биологические протоколы . Архивировано из оригинала 11 октября 2018 года . Проверено 23 марта 2022 г.
- ^ Шеггер Х (2006). «Трицин-СДС-ПАЖ» . Нат Проток . 1 (1): 16–22. дои : 10.1038/нпрот.2006.4 . ПМИД 17406207 . S2CID 209529082 . Архивировано из оригинала 11 июня 2022 года . Проверено 23 марта 2022 г.
- ^ Гордон, AH (1969). Электрофорез белков в полиакриламидных и крахмальных гелях: лабораторные методы биохимии и молекулярной биологии . Амстердам: Паб Северной Голландии. компании ISBN 978-0-7204-4202-1 . ОСЛК 21766 .
- ^ Jump up to: Перейти обратно: а б Кузницы О (1955). «Зональный электрофорез в крахмальных гелях: групповые вариации белков сыворотки нормальных взрослых людей» . Биохим Дж . 61 (4): 629–41. дои : 10.1042/bj0610629 . ПМЦ 1215845 . ПМИД 13276348 .
- ^ Рэксалл Б.Г.; Каллифорд Би Джей (1968). «Тонкослойный крахмальный гель-метод ферментативного типирования пятен крови» . J Forensic Sci Soc . 8 (2): 81–2. дои : 10.1016/s0015-7368(68)70449-7 . ПМИД 5738223 . Архивировано из оригинала 11 июня 2022 года . Проверено 23 марта 2022 г.
- ^ Бьюэлл Дж.Н.; Член парламента Викенса; Пайвар Ф; Шимке РТ (1978). «Синтез полноразмерных кДНК из четырех частично очищенных мРНК яйцевода» . J Биол Хим . 253 (7): 2471–82. дои : 10.1016/S0021-9258(17)38097-3 . ПМИД 632280 .
- ^ Шелп С., Кааден О.Р. (1989). «Усиление полноразмерной транскрипции РНК вируса Синдбис за счет эффективной денатурации гидроксидом метилртути» . Акта Вирол . 33 (3): 297–302. ПМИД 2570517 . Архивировано из оригинала 11 июня 2022 года . Проверено 23 марта 2022 г.
- ^ Фромин Н; Хамелин Дж; Тарнавский С; Роести Д; Журден-Мизер К.; Форестье Н; и др. (2002). «Статистический анализ паттернов дактилоскопии денатурирующего гель-электрофореза (ДГЭ)» . Энвайрон Микробиол . 4 (11): 634–43. дои : 10.1046/j.1462-2920.2002.00358.x . ПМИД 12460271 . Архивировано из оригинала 11 июня 2022 года . Проверено 23 марта 2022 г.
- ^ Фишер С.Г.; Лерман Л.С. (1979). «Независимое от длины разделение фрагментов рестрикции ДНК при двумерном гель-электрофорезе» . Клетка . 16 (1): 191–200. дои : 10.1016/0092-8674(79)90200-9 . ПМИД 369706 . S2CID 9369012 . Архивировано (PDF) из оригинала 11 июня 2022 года . Проверено 23 марта 2022 г.
- ^ Хемпельманн Э; Уилсон Р.Дж. (1981). «Обнаружение глюкозо-6-фосфатдегидрогеназы у малярийных паразитов» . Мол Биохим Паразитол . 2 (3–4): 197–204. дои : 10.1016/0166-6851(81)90100-6 . ПМИД 7012616 . Архивировано из оригинала 6 июля 2023 года . Проверено 23 марта 2022 г.
- ^ Нинфа Эй Джей, Баллоу ДП (1998). Фундаментальные подходы к биохимии и биотехнологии . Бетесда, Мэриленд: Fitzgerald Science Press. ISBN 9781891786006 .
- ^ Нинфа, Александр Дж.; Баллоу, Дэвид П.; Бенор, Мэрили (2009). фундаментальные лабораторные подходы к биохимии и биотехнологии . Хобокен, Нью-Джерси: Уайли. п. 161. ИСБН 978-0470087664 .
- ^ Броуди-младший; Керн С.Е. (2004). «История и принципы проводящих сред для стандартного электрофореза ДНК» . Анальная биохимия . 333 (1): 1–13. дои : 10.1016/j.ab.2004.05.054 . ПМИД 15351274 . Архивировано из оригинала 11 июня 2022 года . Проверено 23 марта 2022 г.
- ^ Лодиш Х; Берк А; Мацудайра П. (2004). Молекулярно-клеточная биология (5-е изд.). WH Фриман: Нью-Йорк, штат Нью-Йорк. ISBN 978-0-7167-4366-8 .
- ^ Устранение неполадок электрофореза ДНК в агарозном геле. Фокус 19:3 с.66 (1997).
- ^ Ханауэр, Матиас; Пьерра, Себастьян; Зинс, Инга; Лотц, Александр; Зоннихсен, Карстен (2007). «Разделение наночастиц с помощью гель-электрофореза по размеру и форме» . Нано-буквы . 7 (9): 2881–2885. Бибкод : 2007NanoL...7.2881H . дои : 10.1021/nl071615y . ПМИД 17718532 .
- ^ Барасински, Маттеус; Гарнвейтнер, Георг (12 февраля 2020 г.). «Механизмы ограниченной и неограниченной миграции наночастиц кремнезема в агарозных гелях и их использование для разделения бинарных смесей». Журнал физической химии C. 124 (9). Американское химическое общество (ACS): 5157–5166. дои : 10.1021/acs.jpcc.9b10644 . ISSN 1932-7447 . S2CID 213566317 .
- ^ Торн Х.В. (1966). «Электрофоретическое отделение ДНК вируса полиомы от ДНК клетки-хозяина» . Вирусология . 29 (2): 234–9. дои : 10.1016/0042-6822(66)90029-8 . ПМИД 4287545 . Архивировано из оригинала 11 июня 2022 года . Проверено 23 марта 2022 г.
- ^ Вебер К; Осборн М (1969). «Надежность определения молекулярной массы методом электрофореза в додецилсульфат-полиакриламидном геле» . J Биол Хим . 244 (16): 4406–12. дои : 10.1016/S0021-9258(18)94333-4 . ПМИД 5806584 .
- ^ Аай С; Борст П. (1972). «Гель-электрофорез ДНК» . Биохим Биофиз Акта . 269 (2): 192–200. дои : 10.1016/0005-2787(72)90426-1 . ПМИД 5063906 . Архивировано из оригинала 11 июня 2022 года . Проверено 23 марта 2022 г.
- ^ Михов, Б. (2022). Основы электрофореза: основная теория и практика . Де Грюйтер, ISBN 9783110761627. doi : 10.1515/9783110761641 . ISBN 9783110761641 .
- ^ Бир, Милан (1959). Электрофорез: теория, методы и приложения . Академическая пресса. п. 225. ОСЛК 1175404 .
- ^ Минд, Дэвид П.; Морис, Мэделон М.; Рюдигер, Стефан Г.Д. (3 октября 2012 г.). Уверский, Владимир Н. (ред.). «Определение биофизической стабильности белков в лизатах с помощью анализа быстрого протеолиза, FASTpp» . ПЛОС ОДИН . 7 (10). Публичная научная библиотека (PLoS): e46147. Бибкод : 2012PLoSO...746147M . дои : 10.1371/journal.pone.0046147 . ISSN 1932-6203 . ПМЦ 3463568 . ПМИД 23056252 .
Внешние ссылки [ править ]
- Демонстрация электрофореза в лаборатории биотехнологий из Учебного центра генетики Университета Юты.
- Прерывистый гель-электрофорез нативных белков
- Электрофорез через питьевую соломинку
- Как запустить гель ДНК или РНК
- Анимация гелевого анализа рестрикции ДНК
- Пошаговые фотографии запуска геля и извлечения ДНК
- Типичный метод из викиверситета