Гель-электрофорез нуклеиновых кислот
Этот раздел нуждается в дополнительных цитатах для проверки . ( Ноябрь 2023 г. ) |


Гель-электрофорез нуклеиновых кислот — это аналитический метод разделения фрагментов ДНК или РНК по размеру и реакционной способности. Молекулы нуклеиновой кислоты помещаются в гель , где электрическое поле побуждает нуклеиновые кислоты (которые заряжены отрицательно из-за их сахаро- фосфатного остова) мигрировать к положительно заряженному аноду . Молекулы разделяются по мере прохождения через гель в зависимости от размера и формы каждой молекулы. Более длинные молекулы движутся медленнее, потому что гель сопротивляется их движению сильнее, чем более коротким молекулам. Через некоторое время электричество отключают и анализируют положения различных молекул.
Нуклеиновую кислоту, которую необходимо разделить, можно получить несколькими способами перед разделением электрофорезом. В случае больших молекул ДНК ДНК часто разрезается на более мелкие фрагменты с помощью эндонуклеазы рестрикции ДНК (или фермента рестрикции). В других случаях, например, в образцах, амплифицированных с помощью ПЦР , ферменты, присутствующие в образце и которые могут повлиять на разделение молекул, удаляются различными способами перед анализом. После того как нуклеиновая кислота приготовлена должным образом, образцы раствора нуклеиновой кислоты помещают в лунки геля и на гель подают напряжение в течение определенного периода времени.
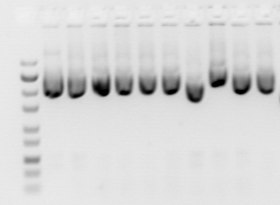
Фрагменты ДНК разной длины визуализируются с помощью специфичного для ДНК флуоресцентного красителя , например бромистого этидия . На геле видны полосы, соответствующие различным популяциям молекул нуклеиновых кислот с разной молекулярной массой. Размер фрагмента обычно указывается в «нуклеотидах», «парах оснований» или «т.п.н.» (для тысяч пар оснований) в зависимости от того, была ли разделена одноцепочечная или двухцепочечная нуклеиновая кислота. Определение размера фрагмента обычно проводится путем сравнения с коммерчески доступными ДНК-маркерами, содержащими линейные фрагменты ДНК известной длины.
Типы гелей, наиболее часто используемые для электрофореза нуклеиновых кислот, - это агароза (для относительно длинных молекул ДНК) и полиакриламид (для высокого разрешения коротких молекул ДНК, например, при секвенировании ДНК ). Гели обычно обрабатываются в формате «пластин», например, показанном на рисунке, но капиллярный электрофорез стал важным для таких приложений, как высокопроизводительное секвенирование ДНК. Методы электрофореза, используемые при оценке повреждения ДНК, включают электрофорез в щелочном геле и гель-электрофорез в импульсном поле .
Для коротких сегментов ДНК, таких как двухцепочечная ДНК длиной от 20 до 60 пар оснований, обработка их в полиакриламидном геле (PAGE) даст лучшее разрешение (нативное состояние). [1] Аналогичным образом, РНК и одноцепочечную ДНК можно анализировать и визуализировать с помощью гелей PAGE, содержащих денатурирующие агенты, такие как мочевина. Гели PAGE широко используются в таких методах, как печать ДНК, EMSA и других методах взаимодействия ДНК-белок.
Измерения и анализ в основном выполняются с помощью специализированного программного обеспечения для анализа гелей. Результаты капиллярного электрофореза обычно отображаются в виде кривой, называемой электрофореграммой .
Факторы, влияющие на миграцию нуклеиновых кислот
[ редактировать ]На миграцию нуклеиновых кислот может влиять ряд факторов: размер пор геля, используемое напряжение, ионная сила буфера и концентрация интеркалирующего красителя, такого как бромид этидия, если он используется во время электрофореза. [2]
Размер ДНК
[ редактировать ]Гель фильтрует ДНК по размеру молекулы ДНК, в результате чего более мелкие молекулы движутся быстрее. Двухцепочечная ДНК движется со скоростью, примерно обратно пропорциональной логарифму числа пар оснований. Однако эта связь нарушается при использовании очень больших фрагментов ДНК, и их невозможно разделить с помощью стандартного электрофореза в агарозном геле . Предел разрешения зависит от состава геля и напряженности поля. [3] а на подвижность более крупной кольцевой ДНК может сильнее влиять размер пор геля, чем на линейную ДНК. [4] Для разделения очень больших фрагментов ДНК требуется импульсный гель-электрофорез (PFGE). При гель-электрофорезе с инверсией поля (FIGE, разновидность PFGE) можно получить «инверсию полос», когда большие молекулы могут двигаться быстрее, чем маленькие молекулы.
Конформация ДНК
[ редактировать ]Конформация обычно движется быстрее, чем расслабленная ДНК , молекулы ДНК может существенно влиять на движение ДНК, например, сверхспиральная ДНК поскольку она плотно скручена и, следовательно, более компактна. В нормальном препарате плазмидной ДНК могут присутствовать несколько форм ДНК. [5] и гель при электрофорезе плазмид обычно показывает основную полосу, которая представляет собой отрицательно сверхспиральную форму, в то время как другие формы ДНК могут проявляться как второстепенные, более слабые полосы. Эти второстепенные полосы могут представлять собой ДНК с надрезом (открытая кольцевая форма) и расслабленная закрытая кольцевая форма, которые обычно работают медленнее, чем суперспиральная ДНК , а одноцепочечная форма (которая иногда может появляться в зависимости от методов получения) может двигаться впереди сверхспиральной ДНК. . Однако скорость, с которой движутся различные формы, может меняться в зависимости от условий электрофореза, например, линейная ДНК может работать быстрее или медленнее, чем сверхспиральная ДНК, в зависимости от условий. [6] а на подвижность более крупной кольцевой ДНК может сильнее влиять размер пор геля, чем на линейную ДНК. [4] сверхспиральной Если не используются маркеры ДНК , размер кольцевой ДНК, подобной плазмиде, может быть более точно измерен после ее линеаризации с помощью рестрикционного расщепления .
Повреждение ДНК из-за увеличения перекрестных связей также будет уменьшать электрофоретическую миграцию ДНК дозозависимым образом. [7] [8]
Концентрация бромистого этидия
[ редактировать ]Концентрация бромида этидия сильнее влияет на кольцевую ДНК, чем на линейную ДНК, если бромид этидия присутствует в геле во время электрофореза. Все встречающиеся в природе кольца ДНК закручены, но бромид этидия, который интеркалируется в кольцевую ДНК, может изменить заряд, длину, а также сверхспиральность молекулы ДНК, поэтому его присутствие во время электрофореза может повлиять на ее движение в геле. Увеличение количества интеркалированного в ДНК бромида этидия может превратить ее из отрицательно свернутой молекулы в полностью релаксированную форму, а затем в положительно закрученную суперспираль при максимальной интеркаляции. [9] Электрофорез в агарозном геле можно использовать для разделения кольцевой ДНК с различной топологией суперспирализации.
Концентрация геля
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Ноябрь 2023 г. ) |
Концентрация геля определяет размер пор геля, который влияет на миграцию ДНК. Разрешение ДНК меняется в зависимости от процентной концентрации геля. Увеличение концентрации агарозы в геле снижает скорость миграции и улучшает разделение более мелких молекул ДНК, тогда как снижение концентрации геля позволяет разделить большие молекулы ДНК. Для стандартного электрофореза в агарозном геле концентрация геля 0,7% дает хорошее разделение или разрешение больших фрагментов ДНК размером 5–10 т.п.н., тогда как концентрация геля 2% дает хорошее разрешение для небольших фрагментов размером 0,2–1 т.п.н. Для разделения очень мелких фрагментов можно использовать гель с концентрацией до 3%, но вертикальный полиакриламидный гель для разделения небольших фрагментов более подходящим будет . Однако гель с высокими концентрациями требует более длительного времени обработки (иногда несколько дней), а гели с высоким процентом часто бывают хрупкими и могут не схватываться равномерно. Агарозные гели с высоким процентом содержания следует использовать с PFGE или FigE. Гели с низким содержанием (0,1–0,2%) хрупкие и могут сломаться. 1%-ные гели распространены для многих применений. [10]
Прикладная область
[ редактировать ]При низком напряжении скорость миграции ДНК пропорциональна приложенному напряжению, т.е. чем выше напряжение, тем быстрее движется ДНК. Однако с увеличением напряженности электрического поля подвижность высокомолекулярных фрагментов ДНК увеличивается дифференциально, а эффективный диапазон разделения уменьшается, и поэтому разрешение при высоком напряжении снижается. Для оптимального разрешения ДНК размером более 2 КБ при стандартном гель-электрофорезе рекомендуется от 5 до 8 В/см. [6] Напряжение также ограничено тем фактом, что оно нагревает гель и может привести к его плавлению, если гель работает при высоком напряжении в течение длительного периода времени, особенно для агарозного геля с низкой температурой плавления.
Однако подвижность ДНК может измениться в нестационарном поле. В поле, которое периодически меняется на противоположное, подвижность ДНК определенного размера может значительно снизиться при определенной частоте циклирования. [11] Это явление может привести к инверсии полос, при которой более крупные фрагменты ДНК движутся в PFGE быстрее, чем более мелкие.
Механизм миграции и разделения
[ редактировать ]Отрицательный заряд его фосфатного остова перемещает ДНК к положительно заряженному аноду во время электрофореза. Однако миграция молекул ДНК в растворе в отсутствие гелевой матрицы не зависит от молекулярной массы во время электрофореза, т.е. без гелевой матрицы не происходит разделения по размерам. [12] Гидродинамическое взаимодействие между различными частями ДНК прерывается потоком противоионов, движущихся в противоположном направлении, поэтому не существует механизма, позволяющего генерировать зависимость скорости от длины в масштабе, превышающем длину экранирования около 10 нм. [11] Это отличает его от других процессов, таких как седиментация или диффузия, где важно долгосрочное гидродинамическое взаимодействие.
Таким образом, гель-матрица отвечает за разделение ДНК по размеру во время электрофореза, однако точный механизм разделения не совсем ясен. Существует ряд моделей механизма разделения биомолекул в гелевой матрице, широко распространенной является модель Огстона, которая рассматривает полимерную матрицу как сито, состоящее из случайно распределенной сети взаимосвязанных пор. [13] Глобулярный белок или случайная спираль ДНК движется через соединенные поры, достаточно большие, чтобы обеспечить его прохождение, а движение более крупных молекул с большей вероятностью будет затруднено и замедлено из-за столкновений с матрицей геля, и поэтому молекулы разных размеров могут быть отделены в этом процессе просеивания. [11]
Однако модель Огстона не работает для больших молекул, когда поры значительно меньше размера молекулы. Для молекул ДНК размером более 1 т.п.н. рептации чаще всего используется модель (или ее варианты). Эта модель предполагает, что ДНК может ползать «змееподобно» (отсюда и «рептация») через поры в виде удлиненной молекулы. При более высокой напряженности электрического поля это превратилось в модель смещенной рептации, в которой передний конец молекулы сильно смещается в прямом направлении, и этот передний край тянет за собой остальную часть молекулы. В полностью смещенном режиме подвижность достигла точки насыщения, и ДНК, превышающая определенный размер, не может быть разделена. [13] Однако на практике идеальное параллельное выравнивание цепи с полем не наблюдается, поскольку это означало бы одинаковую подвижность для длинных и коротких молекул. [11] Дальнейшее уточнение модели смещенной рептации учитывает внутренние колебания цепи. [14]
Модель смещенной рептации также использовалась для объяснения подвижности ДНК в PFGE. Ориентация ДНК постепенно выстраивается в результате рептации после появления поля, и время достижения стационарной скорости зависит от размера молекулы. Когда поле изменяется, более крупным молекулам требуется больше времени для переориентации, поэтому можно отличить длинные цепочки, которые не могут достичь установившейся скорости, от коротких, которые большую часть времени движутся с постоянной скоростью. [14] Однако существуют и другие модели.
Флуоресцентная микроскопия окрашенных молекул в реальном времени показала более тонкую динамику во время электрофореза: ДНК проявляла значительную эластичность, поскольку она поочередно растягивалась в направлении приложенного поля, а затем сжималась в шар или зацеплялась в U-образную форму, когда она зацепляется за полимерные волокна. [15] [16] Это наблюдение можно назвать моделью «гусеницы». [17] Другая модель предполагает, что ДНК запутывается в полимерной матрице, и чем больше молекула, тем больше вероятность того, что она запутается и ее движение затруднится. [18]
Визуализация
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Ноябрь 2023 г. ) |

Наиболее распространенным красителем, используемым для того, чтобы сделать полосы ДНК или РНК видимыми при электрофорезе в агарозном геле, является бромид этидия , обычно обозначаемый сокращенно EtBr. Он флуоресцирует в УФ-свете при внедрении в большую бороздку ДНК (или РНК). Пропуская ДНК через гель, обработанный EtBr, и визуализируя ее в УФ-свете, любая полоса, содержащая более ~20 нг ДНК, становится отчетливо видимой. EtBr — известный мутаген . [19] доступны и более безопасные альтернативы, такие как GelRed производства Biotium , который связывается с малой бороздкой. [20]
SYBR Green I — еще один краситель дцДНК, производимый Invitrogen . Он дороже, но в 25 раз чувствительнее и, возможно, безопаснее, чем EtBr, хотя данных о его мутагенности или токсичности для человека нет. [21]
SYBR Safe — это вариант SYBR Green, который, как было показано, имеет достаточно низкий уровень мутагенности и токсичности, чтобы считаться неопасными отходами в соответствии с федеральными правилами США. [22] Он имеет такой же уровень чувствительности, как и EtBr. [22] но, как и SYBR Green, значительно дороже. Однако в странах, где безопасная утилизация опасных отходов является обязательной, затраты на утилизацию EtBr могут легко превысить первоначальную разницу в цене.
Поскольку ДНК, окрашенная EtBr, не видна при естественном свете, ученые смешивают ДНК с отрицательно заряженными загрузочными буферами, прежде чем добавлять смесь в гель. Загрузочные буферы полезны, поскольку они видны при естественном свете (в отличие от УФ-света для ДНК, окрашенной EtBr), и они осаждаются совместно с ДНК (то есть они движутся с той же скоростью, что и ДНК определенной длины). Ксилолцианол и бромфеноловый синий — распространенные красители, встречающиеся в загрузочных буферах; они работают примерно с той же скоростью, что и фрагменты ДНК длиной 5000 и 300 пар оснований соответственно, но точное положение зависит от процентного содержания геля. Другими менее часто используемыми маркерами прогресса являются Cresol Red и Orange G, которые имеют длину около 125 п.н. и 50 п.н. соответственно.
Визуализация также может быть достигнута путем переноса ДНК после SDS-PAGE на нитроцеллюлозную мембрану с последующим воздействием гибридизационного зонда . Этот процесс называется Саузерн-блоттингом .
Для флуоресцентных красителей после электрофореза гель освещают ультрафиолетовой лампой (обычно помещая ее на световой короб с использованием защитного снаряжения для ограничения воздействия ультрафиолетового излучения). Устройство осветителя в большинстве случаев также содержит устройство формирования изображения, которое снимает изображение геля после освещения УФ-излучением. флуоресцирует Бромид этидия красновато -оранжевым в присутствии ДНК, поскольку он интеркалирован в ДНК. Полосу ДНК также можно вырезать из геля, а затем растворить для получения очищенной ДНК.Затем гель можно сфотографировать, как правило, цифровой камерой или камерой Polaroid. Хотя окрашенная нуклеиновая кислота флуоресцирует красновато-оранжевым цветом, изображения обычно отображаются в черно-белом режиме (см. рисунки). УФ-повреждение образца ДНК может снизить эффективность последующих манипуляций с образцом, таких как лигирование и клонирование. УФ-излучение с более короткой длиной волны (302 или 312 нм) вызывает больший ущерб, например, воздействие в течение всего лишь 45 секунд может значительно уменьшить эффективность трансформации . Поэтому, если ДНК будет использоваться для последующих процедур, следует ограничить воздействие более коротковолнового УФ-излучения, вместо этого следует использовать УФ-излучение с более высокой длиной волны (365 нм), которое вызывает меньший ущерб. Однако излучение с более высокой длиной волны приводит к более слабой флуоресценции, поэтому, если необходимо запечатлеть изображение геля, на короткое время можно использовать ультрафиолетовый свет с более короткой длиной волны. Добавление цитидина или гуанозина в буфер для электрофореза в концентрации 1 мМ может защитить ДНК от повреждения. [23] источник возбуждения синим светом с красителем, возбуждаемым синим светом, таким как SYBR Green или GelGreen В качестве альтернативы можно использовать .
В исследованиях гель-электрофореза часто используются программные инструменты анализа изображений, такие как ImageJ .
| 1 | 2 | 3 |
|---|---|---|
 |  |  |
Ссылки
[ редактировать ]- ^ Джагува Васудеван, Ананда Айяппан; Марио Перкович; Янник Буллиард; Клаус Цихутек; Дидье Трон; Дитер Хаус; Карстен Мунк (август 2013 г.). «Прототип пенистого вируса ухудшает димеризацию и цитозольную растворимость человеческого APOBEC3G» . Журнал вирусологии . 87 (16): 9030–9040. дои : 10.1128/JVI.03385-12 . ПМЦ 3754047 . ПМИД 23760237 .
- ^ Г. Люкотт; Ф. Бэнейкс (1993). Введение в методы молекулярного клонирования . Уайли-Блэквелл. п. 32. ISBN 978-0471188490 .
- ^ Джозеф Сэмбрук; Дэвид Рассел. «Глава 5, протокол 1». Молекулярное клонирование. Лабораторное пособие . Том. 1 (3-е изд.). п. 5.2. ISBN 978-0-87969-577-4 .
- ^ Jump up to: а б Аай С., Борст П. (1972). «Гель-электрофорез ДНК». Биохим Биофиз Акта . 269 (2): 192–200. дои : 10.1016/0005-2787(72)90426-1 . ПМИД 5063906 .
- ^ Ричард Р. Синден (24 ноября 1994 г.). Структура и функции ДНК . Academic Press Inc. с. 97. ИСБН 978-0126457506 .
- ^ Jump up to: а б Джозеф Сэмбрук; Дэвид Рассел. «Глава 5, протокол 1». Молекулярное клонирование. Лабораторное пособие . Том. 1 (3-е изд.). стр. 5.5–5.6. ISBN 978-0-87969-577-4 .
- ^ Бласиак Дж, Тшечак А, Малецка-Панас Е, Джевоски Дж, Воеводска М (2000). «Генотоксичность этанола и ацетальдегида in vitro в лимфоцитах человека и клетках слизистой оболочки желудочно-кишечного тракта». Токсикология in vitro . 14 (4): 287–295. дои : 10.1016/S0887-2333(00)00022-9 . ПМИД 10906435 .
- ^ Лу Й, Моримото К. (2009). «Связано ли регулярное употребление алкоголя со снижением электрофоретической миграции ДНК в лейкоцитах периферической крови у японцев-мужчин с дефицитом ALDH2?» . Мутагенез . 24 (4): 303–308. дои : 10.1093/mutage/gep008 . ПМИД 19286920 .
- ^ Дональд Воэт; Джудит Г. Воэт (1995). Биохимия (2-е изд.). Джон Уайли и сыновья. стр. 877–878 . ISBN 978-0471586517 .
- ^ «Электрофорез в агарозном геле (основной метод)» . Биологические протоколы . Проверено 23 августа 2011 г.
- ^ Jump up to: а б с д Зимм Б.Х., Левен С.Д. (1992). «Проблемы и перспективы теории гель-электрофореза ДНК» (PDF) . Ежеквартальные обзоры биофизики . 25 (2): 171–204. дои : 10.1017/s0033583500004662 . ПМИД 1518924 . S2CID 27976751 .
- ^ Роберт В. Олд; Сэнди Б. Примроуз (27 сентября 1994 г.). Принцип манипуляции генами - Введение в генную инженерию (5-е изд.). Блэквелл Сайентифик. п. 9 . ISBN 9780632037124 .
- ^ Jump up to: а б Ли Чжу; Хун Ван (2 марта 2009 г.). «Глава 4. Генетический анализ в миниатюрных системах электрофореза» . В Тянь, Вэй-Чэн; Файнхаут, Эрин (ред.). Микрофлюидика для биологических приложений . Спрингер. п. 125. ИСБН 978-0-387-09480-9 .
- ^ Jump up to: а б Жан-Луи Виви (2000). «Электрофорез ДНК и других полиэлектролитов: физические механизмы». Обзоры современной физики . 72 (3): 813–872. Бибкод : 2000РвМП...72..813В . дои : 10.1103/RevModPhys.72.813 .
- ^ Смит С.Б., Олдридж П.К., Каллис Дж.Б. (1989). «Наблюдение за отдельными молекулами ДНК, подвергающимися гель-электрофореза». Наука . 243 (4888): 203–206. Бибкод : 1989Sci...243..203S . дои : 10.1126/science.2911733 . ПМИД 2911733 .
- ^ Шварц, округ Колумбия, Коваль М (1989). «Конформационная динамика отдельных молекул ДНК при гель-электрофорезе». Природа . 338 (6215): 520–2. Бибкод : 1989Natur.338..520S . дои : 10.1038/338520a0 . ПМИД 2927511 . S2CID 4249063 .
- ^ Дэвид Шиэн (2009), Физическая биохимия: принципы и приложения (2-е изд.), Wiley-Blackwell, стр. 181, ISBN 978-0470856031
- ^ Форстер Р.Э., Херт Д.Г., Чизл Т.Н., Фредлейк С.П., Бэррон А.Е. (2009). «Анализ механизма миграции ДНК для применения в капиллярном и микрочиповом электрофорезе» . Электрофорез . 30 (12): 2014–24. дои : 10.1002/elps.200900264 . ПМК 2762034 . ПМИД 19582705 .
- ^ Бегусова, М; и др. (2000). «Влияние интеркаляции бромида этидия на радиочувствительность ДНК». Int J Радиат Биол . 76 (1): 1–9. дои : 10.1080/095530000138952 . ПМИД 10665952 . S2CID 218867604 .
- ^ «Самые безопасные и чувствительные гелевые красители нуклеиновых кислот» (PDF) .
- ^ «SYBR Green I Окраска геля нуклеиновых кислот» (PDF) . Архивировано из оригинала (PDF) 22 мая 2012 г. Проверено 23 июня 2013 г.
- ^ Jump up to: а б «SYBR Safe DNA Gel Stain» (PDF) . Архивировано из оригинала (PDF) 7 сентября 2012 г. Проверено 23 июня 2013 г.
- ^ Грюндеманн Д., Шёмиг Э. (1996). «Защита ДНК при препаративном электрофорезе в агарозном геле от повреждений, вызванных ультрафиолетовым светом» . БиоТехники . 21 (5): 898–903. дои : 10.2144/96215rr02 . ПМИД 8922632 .