Агароза

используется для гель-электрофореза
Агароза представляет собой гетерополисахарид , обычно извлекаемый из некоторых красных водорослей . [1] Это линейный полимер, состоящий из повторяющихся звеньев агаробиозы, которая представляет собой дисахарид, состоящий из D -галактозы и 3,6-ангидро- L -галактопиранозы. [2] [3] Агароза является одним из двух основных компонентов агара и очищается от агара путем удаления другого компонента агара, агаропектина . [4]
Агароза часто используется в молекулярной биологии для разделения крупных молекул, особенно ДНК , с помощью электрофореза . Пластинки агарозного геля (обычно 0,7–2%) для электрофореза легко готовят, заливая теплый жидкий раствор в форму. широкий спектр различных агароз различной молекулярной массы Для этой цели коммерчески доступен и свойств. Агарозу также можно формовать в виде шариков и использовать в ряде хроматографических методов очистки белков .
Структура
[ редактировать ]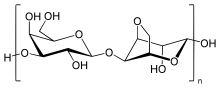
Агароза представляет собой линейный полимер с молекулярной массой около 120 000, состоящий из чередующихся D - галактозы и 3,6-ангидро- L -галактопиранозы, связанных α-(1→3) и β-(1→4) гликозидными связями. 3,6-ангидро- L -галактопираноза представляет собой L -галактозу с ангидромостиком между положениями 3 и 6, хотя некоторые звенья L -галактозы в полимере могут не содержать мостика. Некоторые звенья D -галактозы и L -галактозы могут быть метилированы , а пируват и сульфат . также в небольших количествах обнаруживаются [5]
Каждая агарозная цепь содержит ~800 молекул галактозы, а полимерные цепи агарозы образуют спиральные волокна, которые агрегируются в сверхспиральную структуру радиусом 20-30 нанометров (нм). [6] Волокна квазижесткие и имеют широкий диапазон длины в зависимости от концентрации агарозы. [7] При затвердевании волокна образуют трехмерную сетку каналов диаметром от 50 нм до >200 нм в зависимости от концентрации используемой агарозы - более высокие концентрации приводят к более низким средним диаметрам пор. Трехмерная структура удерживается водородными связями и поэтому может быть разрушена при нагревании до жидкого состояния.
Характеристики
[ редактировать ]Агароза доступна в виде белого порошка, который растворяется в кипящей воде и образует гель при охлаждении. Агароза проявляет явление термического гистерезиса при переходе из жидкости в гель, т.е. она гелеобразует и плавится при разных температурах. Температуры гелеобразования и плавления варьируются в зависимости от типа агарозы. Стандартные агарозы, полученные из Gelidium, имеют температуру гелеобразования 34–38 °C (93–100 °F) и температуру плавления 90–95 °C (194–203 °F), тогда как агарозы, полученные из Gracilaria , из-за более высокой метокси- заместители, имеет температуру гелеобразования 40–52 ° C (104–126 ° F) и температуру плавления 85–90 ° C (185–194 ° F). [8] Температуры плавления и гелеобразования могут зависеть от концентрации геля, особенно при низкой концентрации геля менее 1%. Поэтому температуры гелеобразования и плавления даны при определенной концентрации агарозы.
Природная агароза содержит незаряженные метильные группы, и степень метилирования прямо пропорциональна температуре гелеобразования. Однако синтетическое метилирование имеет обратный эффект: повышенное метилирование снижает температуру гелеобразования. [9] Благодаря химическим модификациям доступны различные химически модифицированные агарозы с различными температурами плавления и гелеобразования.
Агароза в геле образует сетку, содержащую поры, размер пор зависит от концентрации добавленной агарозы. При стоянии агарозные гели склонны к синерезису (экструзии воды через поверхность геля), но этот процесс достаточно медленный, чтобы не мешать использованию геля. [10] [11]
Агарозный гель может иметь высокую прочность геля при низкой концентрации, что делает его подходящим в качестве антиконвекционной среды для гель-электрофореза . Агарозные гели, разбавленные до 0,15%, могут образовывать пластинки для гель-электрофореза. [12] Полимер агарозы содержит заряженные группы, в частности пируват и сульфат . [9] Эти отрицательно заряженные группы могут замедлять движение молекул ДНК в процессе, называемом электроэндосмосом (EEO), и поэтому агароза с низким EEO обычно предпочтительна для использования в электрофорезе нуклеиновых кислот в агарозном геле . Также доступны агарозы с нулевым EEO, но они могут быть нежелательны для некоторых применений, поскольку их можно получить путем добавления положительно заряженных групп, которые могут повлиять на последующие ферментативные реакции. [13] Электроэндосмос является причиной того, что агарозу предпочитают использовать вместо агара, поскольку агаропектин в агаре содержит значительное количество отрицательно заряженных сульфатных и карбоксильных групп. Удаление агаропектина в агарозе существенно снижает ЭЭО, а также снижает неспецифическую адсорбцию биомолекул на матрице геля. Однако для некоторых применений, таких как электрофорез сывороточных белков, может быть желательным высокий уровень ЭЭО, и в используемый гель можно добавлять агаропектин. [14]
Агарозы с низкой температурой плавления и гелеобразования
[ редактировать ]Температуру плавления и гелеобразования агарозы можно изменить путем химической модификации, чаще всего путем гидроксиэтилирования, которое уменьшает количество внутрицепочечных водородных связей, что приводит к более низким температурам плавления и застывания по сравнению со стандартными агарозами. [15] Точная температура определяется степенью замещения, и многие доступные агарозы с низкой температурой плавления (LMP) могут оставаться жидкими в диапазоне 30–35 ° C (86–95 ° F). Это свойство позволяет ферментативные проводить манипуляции непосредственно после гель-электрофореза ДНК путем добавления к реакционной смеси кусочков расплавленного геля, содержащего интересующий фрагмент ДНК. Агароза LMP содержит меньше сульфатов, которые могут влиять на некоторые ферментативные реакции, и поэтому ее предпочтительно использовать для некоторых применений.
Гидроксиэтилированная агароза также имеет меньший размер пор (~ 90 нм), чем стандартные агарозы. [16] Гидроксиэтилирование может уменьшить размер пор за счет уменьшения плотности упаковки агарозных пучков, поэтому гель LMP также может влиять на время и разделение во время электрофореза. [17] Агарозы со сверхнизкой температурой плавления или гелеобразования могут образовывать гель только при 8–15 ° C (46–59 ° F).
Приложения
[ редактировать ]
Агароза является предпочтительной матрицей для работы с белками и нуклеиновыми кислотами , поскольку она обладает широким диапазоном физической, химической и термической стабильности, а ее более низкая степень химической сложности также снижает вероятность ее взаимодействия с биомолекулами . Агароза чаще всего используется в качестве среды для электрофоретического разделения в аналитическом масштабе при электрофорезе в агарозном геле . Гели, изготовленные из очищенной агарозы, имеют относительно большой размер пор, что делает их полезными для разделения крупных молекул, таких как белки и белковые комплексы >200 килодальтон, а также фрагментов ДНК >100 пар оснований. Агароза также широко используется для ряда других применений, например, иммунодиффузии и иммуноэлектрофореза , поскольку волокна агарозы могут выступать в качестве якоря для иммунокомплексов .
Электрофорез в агарозном геле
[ редактировать ]Электрофорез в агарозном геле — стандартный метод разделения ДНК в лаборатории. Агарозные гели имеют более низкую разрешающую способность для ДНК, чем акриламидные гели, но они имеют больший диапазон разделения и поэтому обычно используются для фрагментов ДНК длиной 50–20 000 п.н. ( пар оснований возможно разрешение более 6 Мб. ), хотя при импульсном режиме полевой гель-электрофорез (PFGE). [18] Его также можно использовать для разделения крупных белковых молекул, и он является предпочтительной матрицей для гель-электрофореза частиц с эффективным радиусом более 5-10 нм. [12]
Размер пор геля влияет на размер ДНК, которую можно просеять. Чем ниже концентрация геля, тем больше размер пор и тем больше ДНК можно просеять. Однако гели низкой концентрации (0,1–0,2%) хрупкие, и поэтому с ними трудно обращаться, а электрофорез крупных молекул ДНК может занять несколько дней. Предел разрешения стандартного электрофореза в агарозном геле составляет около 750 кб. [18] Этот предел можно преодолеть с помощью PFGE, когда к гелю прикладывают переменные ортогональные электрические поля. Фрагменты ДНК переориентируются, когда приложенное поле меняет направление, но более крупным молекулам ДНК требуется больше времени, чтобы перестроиться при изменении электрического поля, а более мелким — быстрее, и поэтому ДНК можно фракционировать по размеру.
Агарозные гели отливают в форму и после затвердевания обычно перемещаются горизонтально, погруженные в буферный раствор. трис-ацетат-ЭДТА и трис-борат-ЭДТА Обычно используются буферы другие буферы, такие как трис-фосфат, барбитуровая кислота-барбитурат натрия или трис- барбитуратные буферы. , но в других целях можно использовать и [1] ДНК обычно визуализируется путем окрашивания бромистым этидием , а затем просматривается в УФ-свете , но доступны и другие методы окрашивания, такие как SYBR Green , GelRed , метиленовый синий и кристаллический фиолетовый . Если разделенные фрагменты ДНК необходимы для дальнейшего эксперимента, их можно вырезать из геля срезами для дальнейших манипуляций.
Очистка белка
[ редактировать ]
Матрица агарозного геля часто используется для очистки белков , например, при препаративном масштабном разделении на колонках, например, в гель-фильтрационной хроматографии , аффинной хроматографии и ионообменной хроматографии . Однако он не используется в виде сплошного геля, а формируется в виде пористых шариков или смол различной крупности. [19] Гранулы имеют высокую пористость, поэтому белок может свободно течь через гранулы. Эти шарики на основе агарозы, как правило, мягкие и легко измельчаются, поэтому их следует использовать в процедурах гравитационного потока, низкоскоростного центрифугирования или низкого давления. [20] Прочность смол можно улучшить за счет увеличения поперечных связей и химического отверждения агарозных смол, однако такие изменения могут также привести к снижению связывающей способности белка в некоторых процедурах разделения, таких как аффинная хроматография .
Агароза является полезным материалом для хроматографии, поскольку она не поглощает биомолекулы в сколько-нибудь значительной степени, имеет хорошие свойства текучести и может переносить экстремальные значения pH и ионной силы , а также высокие концентрации денатурантов , таких как 8M мочевина или 6M гуанидин HCl . [21] Примерами матрицы на основе агарозы для гель-фильтрационной хроматографии являются Sepharose и WorkBeads 40 SEC (сшитая гранулированная агароза), Praesto и Superose (гранулированные агарозы с высокой степенью поперечных связей) и Superdex ( декстран, ковалентно связанный с агарозой).
Для аффинной хроматографии гранулированная агароза является наиболее часто используемой матричной смолой для прикрепления лигандов, связывающих белок. [22] Лиганды ковалентно связаны через спейсер с активированными гидроксильными группами агарозного полимера в виде шариков. Затем интересующие белки можно избирательно связать с лигандами, чтобы отделить их от других белков, после чего их можно элюировать. Используемые агарозные гранулы обычно имеют плотность 4% и 6% и обладают высокой способностью связывания белка.
Твердые питательные среды
[ редактировать ]Иногда для культивирования организмов вместо агара можно использовать пластинку с агарозой, поскольку агар может содержать примеси, которые могут повлиять на рост организма или на некоторые последующие процедуры, такие как полимеразная цепная реакция (ПЦР). Агароза также тверже, чем агар, и поэтому может быть предпочтительнее там, где необходима более высокая прочность геля, а ее более низкая температура гелеобразования может предотвратить тепловой шок для организма, когда клетки перед гелеобразованием суспендируются в жидкости. Может быть использован для культуры строгих автотрофных бактерий, протопластов растений , [23] Ценорхабдитис элегантный , [24] другие организмы и различные клеточные линии.
Анализы подвижности
[ редактировать ]Иногда вместо агара для измерения подвижности и подвижности микроорганизмов используют агарозу. Подвижные виды смогут мигрировать, хотя и медленно, по пористому гелю, и тогда можно будет визуализировать скорость проникновения. клеток можно использовать гели различной концентрации Пористость геля напрямую связана с концентрацией агара или агарозы в среде, поэтому для оценки плавающей , роящейся , скользящей и подергивающейся подвижности . Анализ миграции клеток под агарозой можно использовать для измерения хемотаксиса и хемокинеза. Слой агарозного геля помещают между популяцией клеток и хемоаттрактантом . Поскольку градиент концентрации развивается в результате диффузии хемоаттрактанта в гель, различные популяции клеток, требующие разных уровней стимуляции для миграции, затем можно визуализировать с течением времени с помощью микрофотографии, когда они туннелируют вверх через гель против силы тяжести вдоль градиента.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Джеппссон Дж.О., Лорелл С.Б., Франзен Б. (апрель 1979 г.). «Электрофорез в агарозном геле» . Клиническая химия . 25 (4): 629–38. дои : 10.1093/клинчем/25.4.629 . ПМИД 313856 .
- ^ Акари С (1956). «Структура агарозного компонента агар-агара» . Бюллетень Химического общества Японии . 29 (4): 543–544. дои : 10.1246/bcsj.29.543 .
- ↑ Агар . Архивировано 16 октября 2007 г., в Wayback Machine на lsbu.ac.uk. Структура воды и наука.
- ^ «Агар» . Продовольственная и сельскохозяйственная организация Объединенных Наций .
- ^ Армисен Р., Галатас Ф. «Глава 1 – Производство, свойства и использование агара» . Фао.орг.
- ^ Маниатис Т., Фрич Э.Ф., Сэмбрук Дж. (1982). «Глава 5, протокол 1». Молекулярное клонирование. Лабораторное пособие . Том. 1. п. 5.4. ISBN 978-0879691363 .
- ^ Стивен А.М., Phillips GO, ред. (2006). Пищевые полисахариды и их применение . ЦРК Пресс. п. 226. ИСБН 978-0824759223 .
- ^ Семинар по биотехнологии морских водорослей: краткий отчет . Национальная Академия Пресс. 1986. с. 25.
- ^ Перейти обратно: а б «Приложение B: Физическая химия агарозы» (PDF) . Группа компаний «Лонза» . Архивировано (PDF) из оригинала 9 октября 2022 г.
- ^ Хилл С.Э., Ледвард Д.А., Митчелл-младший, ред. (1998). Функциональные свойства пищевых макромолекул . Спрингер. п. 149. ИСБН 978-0-7514-0421-0 .
- ^ Парк Х, Парк К, Шалаби В.С. (1993). Биоразлагаемые гидрогели для доставки лекарств . ЦРК Пресс. п. 102. ИСБН 978-1566760041 .
- ^ Перейти обратно: а б Сервер П (1983). «Агарозные гели: свойства и применение для электрофореза». Электрофорез . 4 (6): 375–382. дои : 10.1002/elps.1150040602 . S2CID 97819634 .
- ^ Сэмбрук Дж., Рассел Д. «Глава 5, протокол 1». Молекулярное клонирование. Лабораторное пособие . Том. 1 (3-е изд.). п. 5.7. ISBN 978-0-87969-577-4 .
- ^ Керен Д. (26 сентября 2003 г.). Белковый электрофорез в клинической диагностике . ЦРК Пресс. стр. 7–8. ISBN 978-0340812136 .
- ^ Маниатис Т., Фрич Э.Ф., Сэмбрук Дж. «Глава 5, протокол 6». Молекулярное клонирование. Лабораторное пособие . Том. 1. п. 5.29. ISBN 978-0879695774 .
- ^ Грисс, Гэри А.; Морено, Елена Т.; Исом, Ричард А.; Сервер, Филип (1989). «Просеивание сфер во время электрофореза в агарозном геле: количественный анализ и моделирование». Биополимеры . 28 (8). Уайли: 1475–1484 гг. дои : 10.1002/bip.360280811 . ISSN 0006-3525 .
- ^ Ли П.Ю., Костумбрадо Дж., Сюй С.И., Ким Ю.Х. (апрель 2012 г.). «Электрофорез в агарозном геле для разделения фрагментов ДНК» . Журнал визуализированных экспериментов . 62 (62): 3923. дои : 10.3791/3923 . ПМЦ 4846332 . ПМИД 22546956 .
- ^ Перейти обратно: а б Маниатис Т., Фрич Э.Ф., Сэмбрук Дж. (1982). «Глава 5, протокол 1». Молекулярное клонирование. Лабораторное пособие . Том. 1. п. 5.2–5.3. ISBN 978-0879691363 .
- ^ Фрайфельдер Д. (1982). Физическая биохимия: приложения к биохимии и молекулярной биологии (2-е изд.). У. Х. Фриман. п. 240. ИСБН 978-0716714446 .
- ^ «Обзор аффинной очистки» . Термо Сайентифик .
- ^ Фрайфельдер Д. (1982). Физическая биохимия: приложения к биохимии и молекулярной биологии (2-е изд.). У. Х. Фриман. п. 258. ИСБН 978-0716714446 .
- ^ Куатрекасас П., Вильчек М. (2004). Леннарц В.Дж., Лейн, доктор медицины (ред.). Энциклопедия биологической химии . Том. 1. Академическая пресса. п. 52. ИСБН 9780124437104 .
- ^ Бонга Дж. М., фон Адеркас П. (1992). Культура деревьев in vitro . Спрингер. п. 16. ISBN 978-0792315407 .
- ^ Колдуэлл Г.А., Уильямс С.Н., Колдуэлл К.А. (2006). Интегрированная геномика: лабораторный курс, основанный на открытиях . Уайли. стр. 94–95. ISBN 978-0470095027 .
| Базы данных органов управления : Национальные |
|---|