дисплей мРНК
Дисплей мРНК — это метод отображения, используемый для in vitro эволюции белков и/или пептидов для создания молекул, которые могут связываться с желаемой мишенью. В результате этого процесса образуются транслируемые пептиды или белки , которые связаны со своим предшественником мРНК посредством пуромициновой связи. Затем комплекс связывается с иммобилизованной мишенью на этапе отбора ( аффинная хроматография ). Слитые мРНК-белки, которые хорошо связываются, затем подвергаются обратной транскрипции в кДНК, и их последовательность амплифицируется посредством полимеразной цепной реакции . В результате получается нуклеотидная последовательность, кодирующая пептид с высоким сродством к интересующей молекуле.
Пуромицин — аналог 3'-конца тирозил-тРНК, часть структуры которого имитирует молекулу аденозина , а другая часть — молекулу тирозина . По сравнению с расщепляемой эфирной связью в тирозил-тРНК, пуромицин имеет негидролизуемую амидную связь. В результате пуромицин препятствует трансляции и вызывает преждевременное высвобождение продуктов трансляции.

Все матрицы мРНК, используемые для технологии отображения мРНК, содержат пуромицин на 3'-конце. По мере трансляции рибосома движется вдоль матрицы мРНК, и как только она достигнет 3'-конца матрицы, слитый пуромицин войдет в сайт А рибосомы и включится в формирующийся пептид. Затем слитая мРНК-полипептид высвобождается из рибосомы (рис. 1).
Для синтеза слияния мРНК-полипептида слитый пуромицин не является единственной модификацией матрицы мРНК. [1] Олигонуклеотиды и другие спейсеры должны быть задействованы вместе с пуромицином, чтобы обеспечить гибкость и достаточную длину для проникновения пуромицина в сайт А. В идеале линкер между 3'-концом мРНК и пуромицином должен быть гибким и достаточно длинным, чтобы позволить пуромицину проникнуть в сайт А при трансляции последнего кодона. Это позволяет эффективно производить высококачественный полноразмерный слитый мРНК-полипептид. Жихэ Лю и др. оптимизировали спейсер 3'-пуромицинового олигонуклеотида. Они сообщили, что dA25 в сочетании со Spacer 9 (Glen Research) и dAdCdCP на 5'-конце лучше всего подходят для реакции синтеза. Они обнаружили, что линкеры длиной более 40 нуклеотидов и короче 16 нуклеотидов демонстрируют значительно сниженную эффективность образования слияния. Кроме того, когда последовательность rUrUP была представлена рядом с пуромицином, слияние не образовывалось эффективно. [2]
Помимо обеспечения гибкости и длины, поли-дА-часть линкера также позволяет осуществлять дальнейшую очистку слитой мРНК-полипептида благодаря ее высокому сродству к dT-целлюлозной смоле. [3] Слитые мРНК-полипептиды можно отбирать по иммобилизованным мишеням селекции в течение нескольких раундов с возрастающей строгостью. После каждого раунда отбора те члены библиотеки, которые остаются связанными с иммобилизованной мишенью, амплифицируются с помощью ПЦР , а несвязывающие вещества смываются.
Метод
[ редактировать ]
Синтез библиотеки дисплея мРНК начинается с синтеза библиотеки ДНК. Библиотеку ДНК для любого интересующего белка или небольшого пептида можно синтезировать методом твердофазного синтеза с последующей ПЦР-амплификацией. Обычно каждый член этой библиотеки ДНК имеет сайт транскрипции РНК-полимеразы Т7 и сайт связывания рибосомы на 5'-конце. Область промотора T7 позволяет осуществлять крупномасштабную транскрипцию T7 in vitro для транскрипции библиотеки ДНК в библиотеку мРНК, которая позже обеспечивает матрицы для реакции трансляции in vitro . Сайт связывания рибосомы в 5'-нетранслируемой области (5'-UTR) конструируют в соответствии с in vitro используемой системой трансляции . Есть две популярные коммерчески доступные in vitro системы перевода . Одним из них является система экстракта E. coli S30 (Promega), для которой требуется последовательность Шайна-Дальгарно в 5'-UTR в качестве сайта связывания рибосомы; [4] другой - Red Nova Lysate (Novagen), которому необходим сайт связывания рибосомы ΔTMV.
После создания библиотеки мРНК ее очищают с помощью электрофореза с мочевиной и лигируют с помощью ДНК-лигазы Т4 к линкеру-спейсеру ДНК, содержащему пуромицин на 3'-конце. На этом этапе лигирования часть мРНК лигируется с одноцепочечной ДНК с помощью ДНК-лигазы Т4. Это не стандартная реакция лигирования ДНК-лигазы Т4, при которой два фрагмента двухцепочечной ДНК связываются вместе. Чтобы увеличить выход этого специального лигирования, можно использовать шину одноцепочечной ДНК для облегчения реакции лигирования. 5'-конец шины комплементарен 3'-концу мРНК, а 3'-конец шины комплементарен 5'-концу линкера-спейсера ДНК, который обычно состоит из поли нуклеотиды dA (рис. 2).
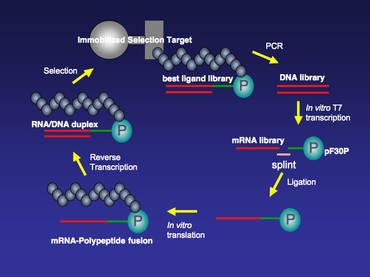
Лигированная библиотека мРНК-ДНК-пуромицин транслируется в лизате Red Nova (Novagen) или E. coli системе экстракта S30 (Promega), в результате чего образуются полипептиды, ковалентно связанные в цис-конце с кодирующей мРНК. Трансляцию in vitro также можно осуществить в системе PURE (синтез белка с использованием рекомбинантных элементов). Система PURE — это E. coli бесклеточная система трансляции , в которой присутствуют только необходимые компоненты трансляции. Некоторые компоненты, такие как аминокислоты и аминоацил-тРНК-синтазы (AARS), могут быть исключены из системы. Вместо этого в систему PURE можно добавить химически ацилированную тРНК. Было показано, что некоторые неприродные аминокислоты, такие как тРНК, акцилированная N-метиламинокислотой, могут быть включены в пептиды или слитые мРНК-полипептиды в системе PURE. [5]
После трансляции одноцепочечные части мРНК слитых материалов будут преобразованы в гетеродуплекс РНК/ДНК с помощью обратной транскриптазы, чтобы устранить любые нежелательные вторичные структуры РНК и сделать часть слитой нуклеиновой кислоты более стабильной. Этот шаг представляет собой стандартную реакцию обратной транскрипции. Например, это можно сделать с помощью Superscript II (GIBCO-BRL), следуя протоколу производителя.
Слитые мРНК/ДНК-полипептиды можно отбирать по иммобилизованным мишеням селекции в течение нескольких раундов (рис. 3). Для первых нескольких раундов отбора может быть относительно высокий фон, и его можно свести к минимуму за счет повышения строгости отбора, например, регулирования концентрации соли, количества детергента и/или температуры в течение периода связывания мишени/слияния. После выбора связывания те члены библиотеки, которые остаются связанными с иммобилизованной мишенью, амплифицируются с помощью ПЦР. Этап ПЦР-амплификации обогатит популяцию из библиотеки мРНК-дисплея, которая имеет более высокое сродство к иммобилизованной мишени. ПЦР, подверженная ошибкам, также может проводиться между каждым раундом отбора, чтобы еще больше увеличить разнообразие библиотеки отображения мРНК и уменьшить фон при отборе. [6]
Недавно был опубликован менее трудоемкий протокол отображения мРНК. [7]
Преимущества
[ редактировать ]Хотя существует множество других технологий молекулярного дисплея, таких как фаговый дисплей , бактериальный дисплей , дрожжевой дисплей и рибосомный дисплей , технология дисплея мРНК имеет много преимуществ по сравнению с другими. [8] Первые три перечисленные библиотеки биологических дисплеев содержат полипептиды или белки, экспрессируемые на поверхности соответствующего микроорганизма, и сопровождающая кодирующая информация для каждого полипептида или белка может быть получена из генома микроорганизма. Однако размер библиотеки для этих трех систем отображения in vivo ограничен эффективностью трансформации каждого организма. Например, размер библиотеки для фагового и бактериального дисплея ограничен 1–10 × 10^9 различными элементами. Размер библиотеки для отображения дрожжей еще меньше. Более того, эти системы клеточного дисплея позволяют проводить скрининг и обогащение только пептидов/белков, содержащих природные аминокислоты. Напротив, дисплей мРНК и дисплей рибосом являются in vitro методами селекции . Они допускают размер библиотеки до 10^15 различных членов. Большой размер библиотеки увеличивает вероятность выбора очень редких последовательностей, а также улучшает разнообразие выбранных последовательностей. Кроме того, методы отбора in vitro устраняют нежелательное давление отбора, такое как плохая экспрессия белка и быстрая деградация белка, что может уменьшить разнообразие выбранных последовательностей. Окончательно, методы селекции in vitro позволяют применять in vitro . мутагенез [9] и методы рекомбинации на протяжении всего процесса отбора.
Хотя и рибосомный дисплей, и дисплей мРНК являются методами селекции in vitro , дисплей мРНК имеет некоторые преимущества перед технологией отображения рибосом. [10] Для дисплея мРНК используются ковалентные комплексы мРНК-полипептид, связанные через пуромицин; тогда как для рибосомного дисплея используются остановленные нековалентные комплексы рибосома-мРНК-полипептид. [11] Для рибосомного дисплея строгость отбора ограничена сохранением комплекса рибосома-мРНК-полипептид из-за нековалентных комплексов рибосома-мРНК-полипептид. Это может вызвать трудности с уменьшением фоновой привязки во время цикла выбора. Кроме того, полипептиды, отбираемые в системе рибосомного дисплея, присоединяются к огромному комплексу рРНК-белок — рибосоме, молекулярная масса которого превышает 2 000 000 Да. Между мишенью селекции и рибосомой может иметь место непредсказуемое взаимодействие, что может привести к потере потенциальных связующих во время цикла селекции. Напротив, спейсерный линкер ДНК пуромицина, используемый в технологии отображения мРНК, намного меньше по сравнению с рибосомой. У этого линкера может быть меньше шансов взаимодействовать с иммобилизованной мишенью селекции. Таким образом, технология отображения мРНК с большей вероятностью даст менее предвзятые результаты.
Приложение
[ редактировать ]В 1997 году Робертс и Шостак показали, что слияния между синтетической мРНК и ее кодируемым эпитопом myc могут быть обогащены из пула слияний мРНК со случайной последовательностью и полипептидом путем иммунопреципитации. [6]
Девять лет спустя Фукуда и его коллеги выбрали метод отображения мРНК для эволюции in vitro фрагментов одноцепочечных Fv (scFv) антител. [12] Они выбрали шесть различных мутантов scFv с пятью консенсусными мутациями. Однако кинетический анализ этих мутантов показал, что их антигенспецифичность остается такой же, как у дикого типа. Однако они продемонстрировали, что две из пяти консенсусных мутаций находились в пределах областей, определяющих комплементарность (CDR). И они пришли к выводу, что дисплей мРНК имеет потенциал для быстрой искусственной эволюции высокоаффинных диагностических и терапевтических антител за счет оптимизации их CDR.
Робертс и его коллеги продемонстрировали, что неприродные пептидные олигомеры, состоящие из N-замещенной аминокислоты, могут быть синтезированы в виде слияний мРНК и полипептида. [13] Пептиды, содержащие N-замещенные аминокислоты, обладают хорошей протеолитической стабильностью и улучшенными фармакокинетическими свойствами. Эта работа показывает, что технология отображения мРНК имеет потенциал для выбора лекарствоподобных пептидов для терапевтического использования, устойчивых к протеолизу. [14]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Робертс Р.В., Шостак Дж.В. (1997). «Слияния РНК-пептидов для отбора пептидов и белков in vitro» . ПНАС . 94 (23): 12297–12302. Бибкод : 1997PNAS...9412297R . дои : 10.1073/pnas.94.23.12297 . ПМК 24913 . ПМИД 9356443 .
- ^ Лю Р., Баррик Дж.Э., Шостак Дж.В., Робертс Р.В. (2000). «Оптимизированный синтез слияний РНК-белок для селекции белков in vitro». Часть B. Взаимодействия РНК-лиганда . Методы энзимологии . Том. 318. стр. 268–93. дои : 10.1016/S0076-6879(00)18058-9 . ISBN 9780121822194 . ПМИД 10889994 .
- ^ Курц М., Гу К., Лозе П.А. (2000). «Конъюгаты мРНК с пуромицином, фотосшитые псораленом: новая матрица для быстрого и простого приготовления слияний мРНК и белка» . Исследования нуклеиновых кислот . 28 (18): 83д–83. дои : 10.1093/нар/28.18.e83 . ПМК 110755 . ПМИД 10982894 .
- ^ Маттеакис Л.К., Бхатт Р.Р., Дауэр В.Дж. (1994). «Система отображения полисом in vitro для идентификации лигандов из очень больших пептидных библиотек» . Proc Natl Acad Sci США . 91 (19): 9022–6. Бибкод : 1994PNAS...91.9022M . дои : 10.1073/pnas.91.19.9022 . ПМЦ 44739 . ПМИД 7522328 .
- ^ Каваками Т., Мураками Х., Шуга Х. (январь 2008 г.). «Запрограммированное РНК-мессенджером включение множества N-метиламинокислот в линейные и циклические пептиды». Химия и биология . 15 (1): 32–42. doi : 10.1016/j.chembiol.2007.12.008 . ПМИД 18215771 .
- ^ Jump up to: а б Робертс Р.В., Шостак Дж.В. (1997). «Слияния РНК-пептидов для отбора пептидов и белков in vitro» . Proc Natl Acad Sci США . 94 (23): 12297–302. Бибкод : 1997PNAS...9412297R . дои : 10.1073/pnas.94.23.12297 . ПМК 24913 . ПМИД 9356443 .
- ^ Барендт П.А., Нг Д.Т., Маккуэйд С.Н., Саркар Калифорния (2013). «Упрощенный протокол отображения мРНК» . АСУ Комбинаторная наука . 15 (2): 77–81. дои : 10.1021/co300135r . ПМК 3666848 . ПМИД 23305392 .
- ^ Робертс Р.В. (июнь 1999 г.). «Полностью селекция белков in vitro с использованием слияний мРНК-белок и рибосомного дисплея». Современное мнение в области химической биологии . 3 (3): 268–73. дои : 10.1016/S1367-5931(99)80042-8 . ПМИД 10359713 .
- ^ Цзин Д., Ли Ф, Цзян М., Цай Дж., Ву Ю, Се К., Ву Икс, Тан С., Лю Дж., Го В., Шэнь Г., Луо Е (ноябрь 2013 г.). «Импульсные электромагнитные поля улучшают микроструктуру и прочность костей у крыс с удаленными яичниками» . ПЛОС ОДИН . 8 (11): e79377. Бибкод : 2013PLoSO...879377J . дои : 10.1371/journal.pone.0079377 . ПМЦ 3828367 . ПМИД 24244491 .
- ^ Золото L (апрель 2001 г.). «Дисплей мРНК: разнообразие имеет значение при селекции in vitro» . Proc Natl Acad Sci США . 98 (9): 4825–6. Бибкод : 2001PNAS...98.4825G . дои : 10.1073/pnas.091101698 . ПМЦ 33119 . ПМИД 11320229 .
- ^ Эндрю Бьюкенен; Лутц Джермутус. «Рибосомный дисплей или метод MRNA-дисплея с селекцией для повышения стабильности белка» . Гугл Патенты . Проверено 9 июня 2014 г.
- ^ Фукуда И., Коджо К., Табата Н. и др. (2006). «Эволюция одноцепочечных антител in vitro с использованием дисплея мРНК» . Исследования нуклеиновых кислот . 34 (19): е127. дои : 10.1093/нар/gkl618 . ПМЦ 1636464 . ПМИД 17012279 .
- ^ Франкель А., Миллуорд С.В., Робертс Р.В. (ноябрь 2003 г.). «Энкодамеры: неприродные пептидные олигомеры, кодируемые РНК» (PDF) . Химия и биология . 10 (11): 1043–50. doi : 10.1016/j.chembiol.2003.11.004 . ПМИД 14652071 .
- ^ Уайт, Э. Рейли; Сан, Люксин; Ма, Чжун; Бекта, Джейсон М.; Данциг, Бретань А.; Хакер, Дэвид Э.; Хьюи, Мелисса; Уильямс, Дэвид К.; Эдвардс, Росс А. (15 мая 2015 г.). «Подход пептидной библиотеки для обнаружения фосфомиметических ингибиторов C-концевого домена BRCA1» . АКС Химическая биология . 10 (5): 1198–1208. дои : 10.1021/cb500757u . ПМЦ 4433557 . ПМИД 25654734 .