QPNC-СТРАНИЦА
QPNC-PAGE , или количественный препаративный непрерывный биохимии в полиакриламидном электрофорез геле , представляет собой биоаналитический , одномерный , высокоточный и высокоточный метод электрофореза , применяемый в и элюирования бионеорганической химии для разделения белков количественного по изоэлектрической точке и путем непрерывного . из геля столбец. [1]
Этот гибридный вариант нативного аналитического гель-электрофореза и препаративного полиакриламидного гель-электрофореза используется биологами для разделения макромолекул в растворе с высоким выходом, например, в активные или нативные металлопротеины в биологических образцах или в правильно и неправильно свернутые белки, содержащие металл -кофактор , или в изоформы белка в сложных белковых смесях . [2]
Введение
[ редактировать ]несколько функций Белки выполняют в живых организмах , включая каталитические реакции и транспорт молекул или ионов внутри клеток , органов или всего тела . Понимание процессов в организме человека, которые в основном обусловлены биохимическими реакциями и белок-белковыми взаимодействиями , в значительной степени зависит от способности выделять активные белки в биологические образцы для более детального изучения химического строения и физиологических функций . Эта важная информация может служить важным показателем состояния здоровья пациента. [3]
Поскольку около 30–40% всех известных белков содержат один или несколько кофакторов ионов металлов (например, церулоплазмин , ферритин , белок-предшественник бета-амилоида , матриксную металлопротеиназу или металлошапероны ), особенно нативные и денатурированные металлопротеины необходимо выделить, идентифицировать и количественно оценить. после жидкой биопсии . Многие из этих кофакторов (например, железо , медь или цинк ) играют ключевую роль в жизненно важных ферментативно- каталитических процессах или стабилизируют молекулы глобулярных белков . [4] Таким образом, высокоточный гель-электрофорез и сопоставимые методы разделения очень актуальны в качестве начального этапа белков и микроэлементов анализа состава , за которым впоследствии следуют современные масс-спектрометрические и магнитно-резонансные методы для количественного определения и идентификации представляющих интерес растворимых белков. [5]
Метод
[ редактировать ]Механизмы разделения и буферизации
[ редактировать ]
В гель-электрофорезе белки обычно разделяются по заряду , размеру или форме . [7] целью изоэлектрического фокусирования Например, pH . (ИЭФ) является разделение белков в соответствии с их изоэлектрической точкой (pI), то есть в соответствии с их зарядом при различных значениях [8] Здесь аналогичный механизм реализуется в имеющейся в продаже камере электрофореза (см. рис. Оборудование ) для разделения заряженных биомолекул , например, супероксиддисмутазы (СОД). [9] или аллергены , [10] в условиях постоянного pH и различных скоростей миграции цвиттер в зависимости от разных изоэлектрических точек -ионов . Отделенные (металлические) белки элюируются последовательно, начиная с самого низкого (pI > 2–4) и заканчивая самым высоким pI (pI < 10,0) растворенных белковых молекул, подлежащих анализу. [11]
Благодаря специфическим свойствам приготовленного геля и буферного раствора для электрофореза , который является основным и содержит трис - HCl и NaN 3 , [6] большинство белков биологической системы (например, Helicobacter pylori [12] ) заряжены отрицательно в растворе и будут мигрировать от катода к аноду под действием электрического поля . В целом уравнение реакции (1) показывает, что карбоксильная боковая группа протеиногенной аминокислоты заряжена отрицательно, уравнение (2) — что боковые аминогруппы электрически нейтральны в этих условиях:
(1) R-СООН + ОН − → R-COO − + Н 2 О
(2) Р-НХ 3 + + ОН − → R-NH 2 + H 2 O
На аноде электрохимически генерируемые . ионы водорода реагируют с молекулами Трис с образованием одновалентных ионов Трис (3) Положительно заряженные ионы Трис мигрируют через гель к катоду, где нейтрализуют ионы гидроксида с образованием молекул Трис и воды (4):
(3) (НОСН 2 ) 3 CNH 2 + Н + → [(НОСН 2 ) 3 CNH 3 ] +
(4) [(НОСН 2 ) 3 CNH 3 ] + + ОН − → (HOCH 2 ) 3 CNH 2 + H 2 O
Таким образом, механизм буферизации на основе Триса обеспечивает постоянный уровень pH в непрерывной буферной системе с высокой буферной емкостью . [13]
При 25 °C Трис-буфер имеет эффективный диапазон pH от 7,5 до 9,0. В приведенных здесь условиях (с учетом концентрации буферных компонентов, механизма буферизации, pH и температуры) эффективный pH смещается в диапазоне примерно от 10,0 до 10,5. Все нативные буферные системы имеют низкую проводимость и диапазон pH от 3,8 до 10,2. Таким образом, непрерывные нативные буферные системы используются для разделения белков в соответствии с их pI. [14]
Хотя значение pH (10,00) буфера для электрофореза не соответствует физиологическому значению pH внутри типа клеток или тканей , разделенные кольцевые белковые полосы непрерывно элюируются в физиологический буферный раствор (pH 8,00) и выделяются в различных фракциях. (ср. рис. Электрофореграмма ). [6] При условии, что необратимая денатурация не может быть продемонстрирована независимой процедурой, большинство белковых молекул стабильны в водном растворе при значениях pH от 3 до 10 и температуре ниже 50 °C. [15] Поскольку Джоулево тепло и температура, образующиеся во время электрофореза, могут превышать 50 °C, [16] и, таким образом, оказывать негативное влияние на стабильность и миграцию белков в геле, система разделения, состоящая из камеры электрофореза, коллектора фракций и других устройств, охлаждается в холодильнике до 4 °C, что значительно снижает риск возникновения теплоконвекционных потоков . [17] Перегреву геля препятствует внутренний контур охлаждения колонки геля как неотъемлемой части камеры электрофореза, а также создание постоянной мощности с помощью источника питания (см. рис. Оборудование ). [18]
Свойства геля и время полимеризации
[ редактировать ]Наилучшие условия полимеризации акриламидных гелей достигаются при температуре 25–30 °C. [19] полимеризация, по-видимому, прекращается через 20–30 мин реакции, хотя остаточные мономеры (10–30%). после этого времени обнаруживаются [20] Сополимеризация мономера акриламида (АА) и N,N'-метиленбисакриламида сшивающего агента (Бис-АА), инициируемая реакциями персульфата аммония (APS)/ тетраметилэтилендиамина (TEMED), наиболее эффективна при щелочном pH раствора акриламида. Таким образом, акриламидные цепи создаются и сшиваются одновременно. Благодаря свойствам буфера для электрофореза гель-полимеризация проводится при pH 10,00, обеспечивая эффективное использование TEMED и APS в качестве катализаторов реакции полимеризации и одновременно подавляя конкурентный гидролиз образующейся сетки акриламидного полимера . Полимерные сети представляют собой трехмерно связанные полимерные цепи. В противном случае белки можно было бы модифицировать путем реакции с неполимеризованными мономерами акриламида, образуя продукты ковалентного акриламида присоединения , что может привести к образованию нескольких полос. [21]
Кроме того, время полимеризации геля может напрямую влиять на время элюирования пиков разделенных металлопротеинов на электрофореграмме из-за сжатия и расширения гелей и их пор, если время инкубации реакционной смеси (раствора геля), используемой для приготовления гель не оптимизированы (см. рис. Электрофореграмма , см. раздел Воспроизводимость и восстановление). Чтобы обеспечить максимальную воспроизводимость размера пор геля и получить полностью полимеризованный и не ограничивающий гель с большими порами для проведения PAGE, полиакриламидный гель полимеризуют в течение 69 часов при комнатной температуре (RT) в гелевой колонке. находится на литейном стенде. [18] Экзотермическое тепло , выделяемое в процессе полимеризации, постоянно рассеивается , при этом температура может быстро подняться до более чем 75 °C в первые минуты, после чего она медленно падает. [22] Через 69 часов гель достигает комнатной температуры и находится в самом низкоэнергетическом состоянии , поскольку основные химические реакции и гелеобразование завершены. [18] Гелеобразование означает, что растворитель (вода) иммобилизуется внутри полимерной сетки посредством водородных связей , а также сил Ван-дер-Ваальса . В результате приготовленный гель является однородным (с точки зрения равномерного распределения поперечных связей по образцу геля). [23] ), по своей природе стабильный и не содержит мономеров и радикалов . Свежие полиакриламидные гели также гидрофильны , электрически нейтральны и не связывают белки. [24] По тем же причинам можно исключить эффекты просеивания, вызванные гравитационным сжатием геля. Таким образом, в среде без свойств молекулярного сита можно ожидать высокого разрешения. [25]
Перед началом электрофореза подготовленный гель, содержащий 4% Т (общее содержание полимера (Т)), 2,67% С (концентрация сшивающего агента (С)) предварительно прогоняется для его уравновешивания . [6] По существу, он не требует просеивания и оптимален для электрофореза белков с молекулярной массой, превышающей или равной 200 k u . Белки мигрируют в нем в большей или меньшей степени на основе своей свободной подвижности. [26] По этим причинам взаимодействие геля с биомолекулами пренебрежимо мало, и, таким образом, белки разделяются чисто и предсказуемо при времени полимеризации 69 часов (см. рис. Электрофореграмма ). Отделенные металлопротеины, включая биомолекулы размером от примерно <1 ku до более 30 ku (например, металлические шапероны , прионы металлов , белки-переносчики , амилоиды , металлоферменты , металлопептиды , металлотионеин , фитохелатины ), не диссоциируются на апопротеины и кофакторы металлов. [27]
Воспроизводимость и восстановление
[ редактировать ]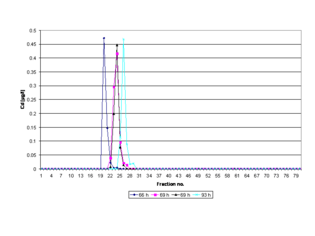
Биоактивные структуры (нативная или трехмерная конформация или форма) изолированных белковых молекул не претерпевают каких-либо существенных конформационных изменений . Таким образом, белки, содержащие кофактор активного металла, могут быть воспроизводимо изолированы в одних и тех же фракциях после проведения PAGE. [11] Смещение пика на соответствующей электрофореграмме указывает на то, что стандартизированное время полимеризации в геле (69 часов, RT) не применяется в эксперименте PAGE . Более низкое отклонение стандартизированного времени полимеризации (<69 часов) означает неполную полимеризацию и, следовательно, внутреннюю нестабильность из-за размягчения геля во время сшивки полимеров, когда материал достигает равновесия набухания. [28] тогда как превышение этого срока (> 69 часов) является показателем старения геля (см. рис. Электрофореграмма ). [29] Явление старения геля тесно связано с долговременным снижением вязкости водных растворов полиакриламида. [30] и повышенное набухание гидрогелей. [31]
В стандартных условиях металлопротеины с различным диапазоном молекулярных масс и изоэлектрических точек были извлечены в биологически активную форму с количественным выходом более 95%. [18] С помощью препаративного SDS-PAGE стандартные белки ( цитохром с , альдолаза , овальбумин и бычий сывороточный альбумин ) с молекулярной массой 14–66 ку могут быть восстановлены со средним выходом около 73,6%. [32] Препаративный изотахофорез (ИТП) применяют для выделения палладийсодержащих белков с молекулярной массой 362 ку (выход: 67%) и 158 ку (выход: 97%). [33]
Количественная оценка и идентификация
[ редактировать ]Физиологические концентрации ( в диапазоне частей на миллиард ) Fe, Cu, Zn, Ni , Mo , Pd, Co , Mn , Pt , Cr , Cd и других металлических кофакторов могут быть идентифицированы и абсолютно количественно определены в аликвоте фракции с помощью массы индуктивно связанной плазмы. спектрометрия (ИСП-МС) [34] или рентгеновская флуоресценция полного отражения (TXRF), [35] например. В случае ИСП-МС структурная информация связанных металлобиомолекул необратимо теряется из-за ионизации образца плазмой . [36] [37] Еще одним признанным высокочувствительным методом определения микроэлементов в биологических образцах является атомно-абсорбционная спектрометрия в графитовой печи (ГФ-ААС) (см. рис. Электроферограмма ). [38] Благодаря высокой чистоте и оптимизированной концентрации разделенных металлопротеинов , например, терапевтических рекомбинантных фармацевтических препаратов растительного происхождения, таких как медный шаперон для супероксиддисмутазы (CCS) из лекарственных растений , в нескольких конкретных фракциях ПААГ родственные структуры этих биоактивных аналитов могут быть выяснена количественно с помощью раствора ЯМР-спектроскопии в неденатурирующих условиях. [39]
Приложения
[ редактировать ]Неправильно свернутые металлические белки, например, CCS или Cu-Zn-супероксиддисмутаза (SOD1), присутствующие в мозге , крови или других клинических образцах, указывают на нейродегенеративные заболевания , такие как болезнь Альцгеймера (БА) или боковой амиотрофический склероз (БАС). [40] Активные молекулы CCS или SOD способствуют внутриклеточному гомеостатическому контролю основных видов ионов металлов (например, Cu 1+/2+ , Зн 2+ , Фе 2+/3+ , Мн 2+ , Является 3+ ) в организмах, и, таким образом, эти биомолекулы могут балансировать прооксидативные и антиоксидантные процессы в цитоплазме . [41] В противном случае свободные или слабосвязанные ионы переходных металлов участвуют в реакциях типа Фентона, в которых образуется вредный гидроксильный радикал , который, если его не сдерживать, будет разрушительным для белков. [42] Потеря активного CCS увеличивает выработку бета-амилоида в нейронах , что, в свою очередь, является основным патологическим признаком AD. [43] Таким образом, медный шаперон супероксиддисмутазы предлагается рассматривать как один из наиболее перспективных биомаркеров меди токсичности при этих заболеваниях. [44] CCS следует анализировать в первую очередь в крови, поскольку метаанализ данных сыворотки показал, что пациенты с АД имеют более высокие уровни Cu в сыворотке, чем здоровые люди из контрольной группы . [45]
Персоны
[ редактировать ]QPNC-PAGE, первоначально называвшийся «PNC-PAGE», был изобретен и разработан в конце 1990-х годов Берндом Кастенхольцем. [46] и значительное влияние оказала новаторская работа Дэвида Э. Гарфина . [47]
См. также
[ редактировать ]- Электрофорез в агарозном геле
- Электроэлюция
- Электрокинетика
- Электрофорез
- Гель-электрофорез
- Гель-электрофорез белков
- Масс-спектрометрия с индуктивно связанной плазмой
- Металлом
- Спектроскопия ядерно-магнитного резонанса белков
- Белковая масс-спектрометрия
- Очистка белка
- Количественная протеомика
- Эксклюзионная хроматография
Ссылки
[ редактировать ]- ^ Зелерт Х., Краузе Ф. (2008). «Препаратное выделение белковых комплексов и других биочастиц путем элюирования из полиакриламидных гелей». Электрофорез . 29 (12): 2617–36. дои : 10.1002/elps.200800061 . ПМИД 18494038 . S2CID 35874355 .
- ^ Кастенхольц Б (2007). «Новая надежда на диагностику и терапию болезни Альцгеймера». Буквы о белках и пептидах . 14 (4): 389–93. дои : 10.2174/092986607780363970 . ПМИД 17504097 .
- ^ Сварт С., Якубовски Н. (2016). «Обновленная информация о состоянии метрологии металлопротеинов» . Журнал аналитической атомной спектрометрии . 31 (9): 1756–65. дои : 10.1039/C6JA00181E .
- ^ Финни Лос-Анджелес, О'Халлоран ТВ (2003). «Видование переходных металлов в клетке: идеи химии рецепторов ионов металлов». Наука . 300 (5621): 931–6. Бибкод : 2003Sci...300..931F . дои : 10.1126/science.1085049 . ПМИД 12738850 . S2CID 14863354 .
- ^ Черкьяро Дж., Маньери Т.М., Бертучи Ф.Р. (2013). «Аналитические методы количественного определения меди, цинка и железа в клетках млекопитающих» . Металломика . 5 (10): 1336–45. дои : 10.1039/c3mt00136a . ПМИД 23925479 .
- ^ Jump up to: а б с д и Кастенхольц Б., Гарфин Д.Е. (2010). «Выделение кислых, основных и нейтральных металлопротеинов с помощью QPNC-PAGE» (PDF) . Предшественники природы : 1–4. дои : 10.1038/npre.2010.4617.1 .
- ^ Гарфин Д.Е. (1995). «Глава 2 – Электрофоретические методы» . Введение в биофизические методы исследования белков и нуклеиновых кислот : 53–109. дои : 10.1016/B978-012286230-4/50003-1 .
- ^ Гарфин Д.Е. (1990). «[35] Изоэлектрическая фокусировка». Изоэлектрическая фокусировка . Методы энзимологии. Том. 182. стр. 459–77. дои : 10.1016/0076-6879(90)82037-3 . ISBN 9780121820831 . ПМИД 2314254 .
- ^ Юн Х.Д., Ким Э.Дж., Роу Дж.Х., Ха Ю.К., Кан СО (1996). «Новая никельсодержащая супероксиддисмутаза из Streptomyces spp» . Биохимический журнал . 318 (Часть 3): 889–96. дои : 10.1042/bj3180889 . ПМЦ 1217701 . ПМИД 8836134 .
- ^ Сак Р., Петерсен А., Вебер Б., Фибиг Х., Кромвель О. (2004). «Аналитический и препаративный электрофорез в нативном полиакриламидном геле: исследование рекомбинантного и природного основного аллергена пыльцы трав Phl p 2». Электрофорез . 25 (1): 14–9. дои : 10.1002/elps.200305697 . ПМИД 14730563 . S2CID 20585733 .
- ^ Jump up to: а б Кастенхольц, Б (2004). «Препаративный непрерывный электрофорез в нативном полиакриламидном геле (PNC-PAGE): эффективный метод выделения кофакторов кадмия в биологических системах». Аналитические письма . 37 (4). Информа Великобритания Лимитед: 657–665. дои : 10.1081/al-120029742 . ISSN 0003-2719 . S2CID 97636537 .
- ^ Бэ С.Х., Харрис А.Г., Хейнс П.Г., Чен Х., Гарфин Д.Е., Хейзелл С.Л., Пайк Ю.К., Уолш Б.Дж., Кордвелл С.Дж. (2003). «Стратегии обогащения и идентификации основных белков в протеомных проектах» . Протеомика . 3 (5): 569–79. дои : 10.1002/pmic.200300392 . ПМИД 12748937 . S2CID 26482563 .
- ^ Кастенхольц Б (2006). «Сравнение электрохимического поведения высокомолекулярных белков кадмия в Arabidopsis thaliana и овощных растениях при использовании препаративного нативного непрерывного электрофореза в полиакриламидном геле (PNC-PAGE)». Электроанализ . 18 (1): 103–6. дои : 10.1002/elan.200403344 .
- ^ Маклеллан Т. (1982). «Буферы для электрофореза для полиакриламидных гелей при различных pH». Аналитическая биохимия . 126 (1): 94–9. дои : 10.1016/0003-2697(82)90113-0 . ПМИД 7181120 .
- ^ Гордон А.Х. (1969). Лабораторные методы в биохимии и молекулярной биологии: электрофорез белков в полиакриламидных и крахмальных гелях (Часть I, Глава 2 Акриламидный гель, 34–45) . Эльзевир. дои : 10.1016/S0075-7535(08)70324-3 . ISBN 9780444533418 .
- ^ Вулли П. (1987). «Термическая нестабильность гелей для электрофореза». Электрофорез . 8 (8): 339–45. дои : 10.1002/elps.1150080802 . S2CID 97116687 .
- ^ Тиселиус А (1937). «Новый аппарат для электрофоретического анализа коллоидных смесей». Труды Фарадеевского общества . 33 : 524–534. дои : 10.1039/TF9373300524 .
- ^ Jump up to: а б с д Кастенхольц Б (2006). «Важный вклад новой количественной препаративной процедуры непрерывного электрофореза в нативном полиакриламидном геле (QPNC-PAGE) для выяснения метаболизма металлических кофакторов при заболеваниях, связанных с неправильным сворачиванием белков - теория». Буквы о белках и пептидах . 13 (5): 503–8. дои : 10.2174/092986606776819637 . ПМИД 16800806 .
- ^ Гельфи С., Ригетти П.Г. (1981). «Кинетика полимеризации полиакриламидных гелей II. Влияние температуры». Электрофорез . 2 (4): 220–28. дои : 10.1002/elps.1150020405 . S2CID 93109120 .
- ^ Чен Б., Крамбах А. (1979). «Оценка эффективности полимеризации при формировании полиакриламидного геля с использованием непрерывного оптического сканирования в процессе полимеризации». Журнал биохимических и биофизических методов . 1 (2): 105–16. дои : 10.1016/0165-022X(79)90017-4 . ПМИД 551105 .
- ^ Бонавентура С., Бонавентура Дж., Стивенс Р., Миллингтон Д. (1994). «Акриламид в полиакриламидных гелях может модифицировать белки во время электрофореза». Аналитическая биохимия . 222 (1): 44–8. дои : 10.1006/abio.1994.1451 . ПМИД 7856869 .
- ^ Уитли М.А., Филлипс Ч.Р. (1983). «Температурные эффекты при полимеризации полиакриламидных гелей, используемых для иммобилизации бактериальных клеток» . Биотехнология и биоинженерия . 25 (2): 623–6. дои : 10.1002/бит.260250228 . ПМИД 18548679 .
- ^ Кизилай МОЙ, Окей О (2003). «Влияние гидролиза на пространственную неоднородность в поли(акриламидных) гелях различной плотности сшивки». Полимер . 44 (18): 5239–50. дои : 10.1016/S0032-3861(03)00494-4 .
- ^ Гарфин Д.Е. (2009). «25-е ежегодное собрание Американского общества электрофореза» . Экспертное обозрение по протеомике . 6 (3): 239–41. дои : 10.1586/апр.09.18 . ПМИД 19489696 . S2CID 24490398 .
- ^ Хертен С (1963). « Молекулярно-ситовый электрофорез в сшитых полиакриламидных гелях». Журнал хроматографии А. 11 : 66–70. дои : 10.1016/S0021-9673(01)80870-0 . ПМИД 13954823 .
- ^ Гарфин Д.Е. (2009) [1990]. «Глава 29 одномерный гель-электрофорез» . Руководство по очистке белков, 2-е издание . Методы энзимологии. Том. 463. стр. 497–513. дои : 10.1016/S0076-6879(09)63029-9 . ISBN 978-0-12-374536-1 . ПМИД 19892189 .
- ^ Фитри Н, Кастенхольц Б, Бухари Б, Амран МБ, Варганегара FM (2008). «Видование молибдена в сыром соке флоэмы клещевины». Аналитические письма . 41 (10): 1773–84. дои : 10.1080/00032710802162442 . S2CID 95715133 .
- ^ Дамлянович В., Лагерхольм Б.К., Якобсон К. (2005). «Объемная и микроструктурная конъюгация белков внеклеточного матрикса с охарактеризованными полиакриламидными субстратами для анализов клеточной механотрансдукции» . БиоТехники . 39 (6): 847–51. дои : 10.2144/000112026 . ПМИД 16382902 .
- ^ Стейскал Дж., Гордон М., Торкингтон Дж.А. (1980). «Коллапс полиакриламидных гелей». Полимерный вестник . 3 (11): 621–5. дои : 10.1007/BF01135333 . S2CID 98565268 .
- ^ Кулицке В.М., Кневске Р., Кляйн Дж. (1982). «Получение, характеристика, свойства раствора и реологическое поведение полиакриламида». Прогресс в науке о полимерах . 8 (4): 373–468. дои : 10.1016/0079-6700(82)90004-1 .
- ^ Юн Дж., Цай С., Суо З., Хейворд Р.К. (2010). «Кинетика пороупругого набухания тонких слоев гидрогеля: сравнение теории и эксперимента». Мягкая материя . 6 (23): 6004–12. Бибкод : 2010SMat....6.6004Y . дои : 10.1039/C0SM00434K . S2CID 2867196 .
- ^ Охаши Т., Моритани С., Андох Х., Сато С., Омори С., Лотспейх Ф., Икеда М. (1991). «Препаративная высокопроизводительная электроэлюция белков после разделения электрофорезом в полиакриламидном геле с додецилсульфатом натрия и ее применение для анализа аминокислотных последовательностей и получения антител» . Журнал хроматографии . 585 (1): 153–9. дои : 10.1016/0021-9673(91)85069-р . ПМИД 1666109 .
- ^ Вебер Г., Мессершмидт Дж., фон Болен А., Кастенхольц Б., Гюнтер К. (2004). «Улучшенное разделение палладия в биологических матрицах с помощью сочетания гель-проникающей хроматографии и изотахофореза». Электрофорез . 25 (12): 1758–64. дои : 10.1002/elps.200305833 . ПМИД 15213973 . S2CID 22292130 .
- ^ Рашовский П (2011). «Использование колоночного гель-электрофореза для онлайн-подключения к ИСП-МС для металлопротеомики» . Архив диссертаций Масариковского университета, Брно.
- ^ Пессанья С., Карвалью М.Л., Беккер М., фон Болен А. (2010). «Количественное определение тяжелых металлов на разных стадиях производства вина методами рентгеновской флуоресценции полного отражения и энергодисперсионной рентгеновской флуоресценции: сравнение на двух виноградниках». Spectrochimica Acta Часть B: Атомная спектроскопия . 65 (6): 504–7. Бибкод : 2010AcSpB..65..504P . дои : 10.1016/j.sab.2010.04.003 .
- ^ Якубовский Н., Лобински Р., Моенс Л. (2004). «Металлобиомолекулы. Основа жизни, задача атомной спектроскопии» . Журнал аналитической атомной спектрометрии . 19 (1): 1–4. дои : 10.1039/B313299B .
- ^ Мунику С., Шпунар Дж., Лобински Р. (2009). «Металломика: понятие и методология». Обзоры химического общества . 38 (4): 1119–38. дои : 10.1039/B713633C . ПМИД 19421584 .
- ^ Линь Т.В., Хуан С.Д. (2001). «Прямое и одновременное определение меди, хрома, алюминия и марганца в моче с помощью многоэлементного атомно-абсорбционного спектрометра с графитовой печью». Аналитическая химия . 73 (17): 4319–25. дои : 10.1021/ac010319h . ПМИД 11569826 .
- ^ Кастенхольц Б., Гарфин Д.Е. (2009). «Лекарственные растения: природный источник шаперонов для лечения неврологических расстройств». Буквы о белках и пептидах . 16 (2): 116–20. дои : 10.2174/092986609787316234 . ПМИД 19200033 .
- ^ Шюман К., Классен Х.Г., Дитер Х.Х., Кениг Дж., Мультауп Г., Рюкгауэр М., Саммер К.Х., Бернхардт Дж., Бисальски Х.К. (2002). «Семинар по консенсусу в Хоэнхайме: медь» . Европейский журнал клинического питания . 56 (6): 469–83. дои : 10.1038/sj.ejcn.1601315 . ПМИД 12032645 .
- ^ Кастенхольц Б. , Гарфин Д.Е. , Хорст Дж., Нагель К.А. (2009). «Металлические шапероны растений: новый взгляд на терапию деменции». Амилоид . 16 (2): 81–83. дои : 10.1080/13506120902879392 . ПМИД 20536399 . S2CID 37490474 .
- ^ Робинсон, Нью-Джерси, Виндж ДР (2010). «Медные металлочапероны» . Ежегодный обзор биохимии . 79 : 537–62. doi : 10.1146/annurev-biochem-030409-143539 . ПМЦ 3986808 . ПМИД 20205585 .
- ^ Грей Э.Х., Де Вос К.Дж., Дингуолл С., Перкинтон М.С., Миллер CC (2010). «Дефицит медного шаперона супероксиддисмутазы увеличивает выработку бета-амилоида» . Журнал болезни Альцгеймера . 21 (4): 1101–5. дои : 10.3233/JAD-2010-100717 . ПМК 3023902 . ПМИД 20693630 .
- ^ Пал А (2014). «Токсичность меди, вызванная гепатоцеребральными и нейродегенеративными заболеваниями: острая необходимость в прогностических биомаркерах». Нейротоксикология . 40 : 97–101. Бибкод : 2014NeuTx..40...97P . дои : 10.1016/j.neuro.2013.12.001 . ПМИД 24342654 .
- ^ Букосси С., Вентриглия М., Панетта В., Салустри С., Паскуалетти П., Мариани С., Сиотто М., Россини П.М., Сквитти Р. (2011). «Медь при болезни Альцгеймера: метаанализ исследований сыворотки, плазмы и спинномозговой жидкости» . Журнал болезни Альцгеймера . 24 (1): 175–85. дои : 10.3233/JAD-2010-101473 . ПМИД 21187586 . S2CID 33194620 .
- ^ «Метод выделения кофакторов металлов из биологических органических систем с использованием препаративного непрерывного гель-электрофореза в нативном полиакриламиде (PNC-PAGE)» . Гугл Патенты . Проверено 15 июля 2024 г.
- ^ Финщенко Ю., Саламанзаде А., Давалос Р. (2014). «AES 2013: Ежегодное собрание общества электрофореза AES». Американская лаборатория . 46 (6).
Дальнейшее чтение
[ редактировать ]- Михов, Б (2022). Основы электрофореза: основная теория и практика . Де Грюйтер, ISBN 9783110761627. doi : 10.1515/9783110761641 . ISBN 9783110761641 .
- Финщенко Ю., Саламанзаде А., Давалос Р. (2014). «AES 2013: Ежегодное собрание Общества электрофореза AES» . Американская лаборатория . Проверено 19 апреля 2024 г.
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) CS1 maint: числовые имена: список авторов ( ссылка )
Внешние ссылки
[ редактировать ]- Развитие электрокинетической науки Обществом электрофореза AES
- Протоколы очистки белка Еврейского университета в Иерусалиме