Супероксиддисмутаза ( СОД , EC 1.15.1.1 ) представляет собой фермент , который попеременно катализирует дисмутацию (или разделение) супероксида ( O − 2 ) анион- радикал на нормальный молекулярный кислород (O 2 ) и перекись водорода ( H 22О 2 ). Супероксид вырабатывается как побочный продукт метаболизма кислорода и, если его не регулировать, вызывает многие виды повреждений клеток. [2] Перекись водорода также вредна и разлагается другими ферментами, такими как каталаза . Таким образом, СОД является важной антиоксидантной защитой почти во всех живых клетках, подвергающихся воздействию кислорода. Единственным исключением являются Lactobacillus plantarum и родственные лактобактерии , которые используют внутриклеточный марганец для предотвращения повреждения от реактивного O. − 2 . [3] [4]
Ирвин Фридович и Джо МакКорд из Университета Дьюка обнаружили ферментативную активность супероксиддисмутазы в 1968 году. [5] Ранее СОД были известны как группа металлопротеинов с неизвестной функцией; например, CuZnSOD был известен как эритрокупреин (или гемокупреин, или цитокупреин) или как ветеринарный противовоспалительный препарат «Орготеин». [6] Точно так же Брюэр (1967) идентифицировал белок, который позже стал известен как супероксиддисмутаза, как индофенолоксидазу, путем анализа белков крахмальных гелей с использованием метода феназина-тетразолия. [7]
Существует три основных семейства супероксиддисмутазы, в зависимости от структуры белка и металлического кофактора : тип Cu/Zn (который связывает как медь, так и цинк ), типы Fe и Mn (которые связывают либо железо, либо марганец ) и тип Ni. (который связывает никель ).
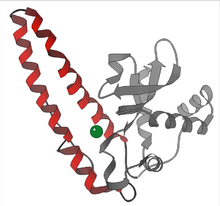
Активный сайт СОД человеческого марганца, марганец показан фиолетовым цветом. [9]
Димеры Mn-SOD и Fe-SOD
Медь и цинк – чаще всего используются эукариотами , включая человека. Цитозоли (Cu-Zn - практически всех эукариотических клеток содержат фермент СОД с медью и цинком СОД). Например, коммерчески доступный Cu-Zn-SOD обычно очищают из бычьих эритроцитов. Бычий фермент Cu-Zn представляет собой гомодимер с молекулярной массой 32500. Это был первый СОД, чья кристаллическая структура атомных деталей была решена в 1975 году. [10] Это 8-нитевой бета-цилиндр « греческого ключа », активный центр которого находится между стволом и двумя поверхностными петлями. Две субъединицы плотно соединены друг с другом, в основном за счет гидрофобных и некоторых электростатических взаимодействий. Лигандами меди и цинка являются шесть гистидиновых и одна аспартатная боковые цепи; один гистидин связан между двумя металлами. [11]
Железо. Многие бактерии содержат форму фермента с железом (Fe-SOD); некоторые бактерии содержат Fe-СОД, другие Mn-СОД, а некоторые (например, E. coli ) содержат и то, и другое. Fe-СОД также можно найти в хлоропластах растений. Трехмерные структуры гомологичных супероксиддисмутаз Mn и Fe имеют одинаковое расположение альфа-спиралей, а их активные центры содержат одинаковый тип и расположение боковых цепей аминокислот. Обычно это димеры, но иногда и тетрамеры.
Марганец. Почти все митохондрии и многие бактерии содержат форму с марганцем (Mn-SOD). Например, Mn-SOD обнаружен в митохондриях человека. Лигандами ионов марганца являются 3 гистидина боковые цепи , аспартатная боковая цепь и молекула воды или гидроксилиганд , в зависимости от степени окисления Mn (соответственно II и III). [12]
Никель – прокариотический . Он имеет гексамерную (6-копийную) структуру, построенную из правозакрученных пучков из 4 спиралей, каждый из которых содержит N-концевые крючки, которые хелатируют ион Ni. Ni-крючок содержит мотив His-Cys-XX-Pro-Cys-Gly-X-Tyr; он обеспечивает большинство взаимодействий, критически важных для связывания металлов и катализа, и, следовательно, является вероятным диагностическим средством NiSOD. [13] [14]
У высших растений изоферменты СОД локализованы в разных клеточных компартментах. Mn-СОД присутствует в митохондриях и пероксисомах . Fe-СОД обнаружен главным образом в хлоропластах, но также обнаружен в пероксисомах, а CuZn-СОД локализован в цитозоле , хлоропластах, пероксисомах и апопласте . [16] [17]
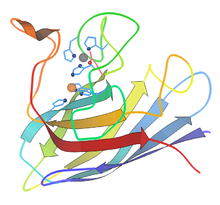
Кристаллическая структура человеческого фермента SOD1 ( N-конец радужного цвета = синий, C-конец = красный) в комплексе с медью (оранжевая сфера) и цинком (серая сфера) [18]
Кристаллографическая структура тетрамерного человеческого фермента SOD3 (мультяшная диаграмма) в комплексе с катионами меди и цинка (оранжевая и серая сферы соответственно) [19]
В высших растениях ферменты супероксиддисмутазы (СОД) действуют как антиоксиданты и защищают клеточные компоненты от окисления активными формами кислорода (АФК). [20] АФК могут образовываться в результате засухи, травм, гербицидов и пестицидов, озона, метаболической активности растений, дефицита питательных веществ, фотоингибирования, температуры над и под землей, токсичных металлов, а также ультрафиолетовых или гамма-лучей. [21] [22] Если быть конкретнее, молекулярный O 2 восстанавливается до O − 2 (АФК, называемая супероксидом), когда она поглощает возбужденный электрон, высвободившийся из соединений цепи переноса электронов. Известно, что супероксид денатурирует ферменты, окисляет липиды и фрагментирует ДНК. [21] СОД катализируют образование O 2 и H 22О 2 из супероксида ( O − 2 ), что приводит к уменьшению вредных реагентов.
При адаптации к повышенному уровню окислительного стресса концентрации СОД обычно увеличиваются с увеличением степени стресса. Компартментализация различных форм СОД по всему растению позволяет им очень эффективно противодействовать стрессу. В растениях существуют три хорошо известных и изученных класса металлических коферментов СОД. Во-первых, Fe-СОД состоят из двух разновидностей: гомодимера (содержащего 1–2 г Fe) и тетрамера (содержащего 2–4 г Fe). Считается, что они являются древнейшими металлоферментами СОД и встречаются как у прокариот, так и у эукариот. Fe СОД наиболее часто локализуются внутри хлоропластов растений, где они являются аборигенными. Во-вторых, СОД Mn состоят из разновидностей гомодимера и гомотетрамера, каждый из которых содержит один атом Mn (III) на субъединицу. Они обнаруживаются преимущественно в митохондриях и пероксисомах. В-третьих, электрические свойства СОД Cu-Zn сильно отличаются от свойств двух других классов. Они концентрируются в хлоропластах , цитозоле и в некоторых случаях во внеклеточном пространстве. Обратите внимание, что СОД Cu-Zn обеспечивают меньшую защиту, чем СОД Fe, когда они локализованы в хлоропластах. [20] [21] [22]
Лейкоциты человека используют такие ферменты, как НАДФН-оксидаза, для выработки супероксида и других активных форм кислорода для уничтожения бактерий. Поэтому во время инфекции некоторые бактерии (например, Burkholderia pseudomallei ) вырабатывают супероксиддисмутазу, чтобы защитить себя от гибели. [23]
СОД превосходит повреждающие реакции супероксида, тем самым защищая клетку от токсичности супероксида.Реакция супероксида с нерадикалами запрещена по спину . В биологических системах это означает, что его основные реакции происходят с самим собой (дисмутация) или с другим биологическим радикалом, таким как оксид азота (NO), или с металлом переходного ряда. Супероксидный анион-радикал ( O − 2 ) самопроизвольно дисмутирует на O 2 и перекись водорода ( H 22О 2 ) довольно быстро (~10 5 М −1 с −1 при pH 7). [ нужна ссылка ] СОД необходим, поскольку супероксид реагирует с чувствительными и важными клеточными мишенями. Например, он реагирует с радикалом NO и образует токсичный пероксинитрит .
Поскольку некаталитическая реакция дисмутации супероксида требует взаимодействия двух молекул супероксида друг с другом, скорость дисмутации имеет второй порядок по отношению к начальной концентрации супероксида. Таким образом, период полураспада супероксида, хотя и очень короткий при высоких концентрациях (например, 0,05 секунды при 0,1 мМ), на самом деле довольно длительный при низких концентрациях (например, 14 часов при 0,1 нМ). Напротив, реакция супероксида с СОД имеет первый порядок по концентрации супероксида. Более того, супероксиддисмутаза имеет самое большое k кат / K M (приблизительное значение каталитической эффективности) среди всех известных ферментов (~7 x 10 9 М −1 с −1 ), [24] эта реакция ограничена только частотой столкновений между ней и супероксидом. То есть скорость реакции «диффузионно-ограничена».
Высокая эффективность супероксиддисмутазы кажется необходимой: даже при субнаномолярных концентрациях, достигаемых за счет высоких концентраций СОД внутри клеток, супероксид инактивирует фермент аконитазу цикла лимонной кислоты , может отравлять энергетический обмен и высвобождать потенциально токсичное железо. Аконитаза является одной из нескольких железосеросодержащих (де)гидратаз в метаболических путях, которые, как было показано, инактивируются супероксидом. [25]
SOD1 — чрезвычайно стабильный белок. В голо-форме (связанной как с медью, так и с цинком) температура плавления составляет > 90 °C. В апо-форме (не связанной с медью или цинком) температура плавления составляет ~ 60 ° C. [26] Методом дифференциальной сканирующей калориметрии (ДСК) голо-СОД1 разворачивается по двухсостоятельному механизму: от димера до двух развернутых мономеров. [26] В экспериментах по химической денатурации голо-СОД1 разворачивается по трехуровневому механизму с наблюдением свернутого мономерного промежуточного продукта. [27]
Супероксид – одна из основных активных форм кислорода в клетке. Как следствие, СОД выполняет ключевую антиоксидантную роль. Физиологическое значение СОД иллюстрируется тяжелыми патологиями, наблюдаемыми у мышей, генетически модифицированных с отсутствием этих ферментов. Мыши, у которых отсутствует SOD2, умирают через несколько дней после рождения на фоне массивного окислительного стресса . [28] У мышей, лишенных SOD1, развивается широкий спектр патологий, включая гепатоцеллюлярную карциному, [29] ускорение возрастной потери мышечной массы, [30] более раннее возникновение катаракты и сокращение продолжительности жизни. Мыши, у которых отсутствует SOD3, не имеют каких-либо очевидных дефектов и имеют нормальную продолжительность жизни, хотя они более чувствительны к гипероксическому повреждению. [31] Мыши с нокаутом любого фермента СОД более чувствительны к летальному воздействию соединений, генерирующих супероксид, таких как паракват и дикват ( гербициды ).
У дрозофил, лишенных SOD1, продолжительность жизни резко сокращается, тогда как мухи, лишенные SOD2, умирают до рождения. Истощение СОД1 и СОД2 в нервной системе и мышцах дрозофилы связано с сокращением продолжительности жизни. [32] Накопление нейрональных и мышечных АФК , по-видимому, способствует возрастным нарушениям. Когда индуцируется сверхэкспрессия митохондриального SOD2, продолжительность жизни взрослой дрозофилы увеличивается. [33]
У черных садовых муравьев ( Lasius niger ) продолжительность жизни маток на порядок больше, чем у рабочих, несмотря на отсутствие систематических различий в нуклеотидных последовательностях между ними. [34] Было обнаружено, что ген SOD3 наиболее дифференциально сверхэкспрессируется в мозге королевы и рабочих муравьев. Это открытие повышает вероятность важной роли антиоксидантной функции в модуляции продолжительности жизни. [34]
Нокдаун СОД у червя C. elegans не вызывает серьезных физиологических нарушений. Однако продолжительность жизни C. elegans можно продлить с помощью миметиков супероксида/ каталазы, что позволяет предположить, что окислительный стресс является основным фактором, определяющим скорость старения . [35]
Нокаутные или нулевые мутации в SOD1 очень вредны для аэробного роста почкующихся дрожжей Saccharomyces cerevisiae и приводят к резкому сокращению продолжительности жизни после диауксии. типа У S. cerevisiae дикого степень повреждения ДНК увеличивалась с возрастом в 3 раза, но более чем в 5 раз у мутантов с удаленными генами SOD1 или SOD2 . [36] Уровни активных форм кислорода увеличиваются с возрастом в этих мутантных штаммах и демонстрируют закономерность, аналогичную закономерности увеличения повреждений ДНК с возрастом. Таким образом, оказывается, что супероксиддисмутаза играет существенную роль в сохранении целостности генома во время старения S. cerevisiae .Нокаут SOD2 или нулевые мутации вызывают задержку роста дыхательных источников углерода в дополнение к уменьшению продолжительности жизни после диауксии.
Было создано несколько прокариотических нулевых мутантов SOD, включая E. coli . Потеря периплазматического CuZnSOD приводит к потере вирулентности и может стать привлекательной мишенью для новых антибиотиков.
Мутации в SOD1 могут вызывать семейный БАС (некоторые данные также показывают, что SOD1 дикого типа в условиях клеточного стресса участвует в значительной части спорадических случаев БАС, которые составляют 90% пациентов с БАС). [45] по механизму, который в настоящее время не понятен, но не за счет потери ферментативной активности или снижения конформационной стабильности белка SOD1. Сверхэкспрессия SOD1 связана с нервными расстройствами, наблюдаемыми при синдроме Дауна . [46] У пациентов с талассемией СОД будет увеличиваться как форма компенсационного механизма. Однако в хронической стадии СОД оказывается недостаточной и имеет тенденцию к снижению из-за разрушения белков в результате массивной реакции оксидант-антиоксидант. [47]
У мышей внеклеточная супероксиддисмутаза (SOD3, ecSOD) способствует развитию гипертонии . [48] [49] Инактивация СОД2 у мышей приводит к перинатальной летальности. [28]
Дополнительная супероксиддимутаза была предложена в качестве лечения для предотвращения бронхолегочной дисплазии у недоношенных детей , однако эффективность этого лечения не ясна. [50]
СОД использовался в экспериментальном лечении хронического воспаления при воспалительных заболеваниях кишечника. [51] [52] СОД может уменьшить цис-платиной , вызванную нефротоксичность (исследования на грызунах). [53] Как «Орготеин» или «онтозеин», фармакологически активный очищенный СОД из бычьей печени, он также эффективен при лечении воспалительных заболеваний мочевыводящих путей у человека. [54] Какое-то время СОД из бычьей печени даже имел одобрение регулирующих органов в нескольких европейских странах для такого использования. Это было прервано опасениями по поводу прионных заболеваний . [ нужна ссылка ]
Агент -миметик СОД , TEMPOL , в настоящее время проходит клинические испытания для радиозащиты и предотвращения радиационно-индуцированного дерматита . [55] TEMPOL и аналогичные нитроксиды-миметики СОД проявляют множественное действие при заболеваниях, связанных с окислительным стрессом. [56]
Синтез таких ферментов, как супероксиддисмутаза, L-аскорбатоксидаза и ДНК-полимераза Дельта 1 , инициируется у растений при активации генов , связанных со стрессовыми состояниями растений. [57] Наиболее распространенными стрессовыми условиями могут быть травмы, засуха или засоление почвы . Ограничить этот процесс, инициируемый условиями сильного засоления почвы, можно путем введения в растения экзогенного глутамина . Снижение уровня экспрессии генов, ответственных за синтез супероксиддисмутазы, увеличивается с увеличением концентрации глутамина. [57]
СОД может уменьшить повреждение кожи свободными радикалами, например, уменьшить фиброз после облучения при раке молочной железы. Однако исследования такого рода следует рассматривать как предварительные, поскольку в исследовании не было адекватного контроля, включая отсутствие рандомизации, двойного слепого метода или плацебо. [58] Известно, что супероксиддисмутаза обращает вспять , возможно, за счет дедифференцировки миофибробластов обратно фиброз в фибробласты . [59] [ нужны дальнейшие объяснения ]
СОД коммерчески получают из морского фитопланктона , бычьей печени, хрена , дыни и некоторых бактерий. С терапевтической целью СОД обычно вводят местно. Нет никаких доказательств того, что употребление незащищенных СОД или продуктов, богатых СОД, может иметь какие-либо физиологические эффекты, поскольку весь проглоченный СОД расщепляется на аминокислоты перед всасыванием . Однако прием внутрь СОД, связанного с белками пшеницы, может улучшить его терапевтическую активность, по крайней мере теоретически. [60]
^ Jump up to: а б PDB : 1ВАР ; Боргшталь Г.Е., Пардж Х.Э., Хики М.Дж., Джонсон М.Дж., Буассино М., Холлевелл Р.А. и др. (апрель 1996 г.). «Полиморфный вариант Ile58Thr митохондриальной супероксиддисмутазы марганца человека снижает активность за счет дестабилизации тетрамерного интерфейса». Биохимия . 35 (14): 4287–4297. дои : 10.1021/bi951892w . ПМИД 8605177 . S2CID 7450190 .
^ Квинт П., Ройцель Р., Микульски Р., Маккенна Р., Сильверман Д.Н. (февраль 2006 г.). «Кристаллическая структура нитрированной супероксиддисмутазы марганца человека: механизм инактивации». Свободно-радикальная биология и медицина . 40 (3): 453–458. doi : 10.1016/j.freeradbiomed.2005.08.045 . ПМИД 16443160 .
^ Румфельдт Дж.А., Статопулос П.Б., Чакрабаррти А., Лепок Дж.Р., Мейеринг Э.М. (январь 2006 г.). «Механизм и термодинамика гуанидинхлорид-индуцированной денатурации мутантных супероксиддисмутаз Cu,Zn, связанных с БАС». Журнал молекулярной биологии . 355 (1): 106–123. дои : 10.1016/j.jmb.2005.10.042 . ПМИД 16307756 .
^ Jump up to: а б Ли Ю, Хуанг Т.Т., Карлсон Э.Дж., Мелов С., Урселл П.С., Олсон Дж.Л. и др. (декабрь 1995 г.). «Дилятационная кардиомиопатия и неонатальная смертность у мутантных мышей, у которых отсутствует супероксиддисмутаза марганца». Природная генетика . 11 (4): 376–381. дои : 10.1038/ng1295-376 . ПМИД 7493016 . S2CID 10900822 .
^ Мюллер Ф.Л., Сонг В., Лю Ю., Чаудхури А., Пик-Даль С., Стронг Р. и др. (июнь 2006 г.). «Отсутствие супероксиддисмутазы CuZn приводит к повышенному окислительному стрессу и ускорению возрастной атрофии скелетных мышц». Свободно-радикальная биология и медицина . 40 (11): 1993–2004. doi : 10.1016/j.freeradbiomed.2006.01.036 . ПМИД 16716900 .
^ Ока С., Хираи Дж., Ясукава Т., Накахара Ю., Иноуэ Ю.Х. (август 2015 г.). «Корреляция накопления активных форм кислорода за счет истощения супероксиддисмутазы с возрастными нарушениями в нервной системе и мышцах взрослых дрозофил». Биогеронтология . 16 (4): 485–501. дои : 10.1007/s10522-015-9570-3 . ПМИД 25801590 . S2CID 18050827 .
^ Муид К.А., Каракая Х.К., Коч А. (февраль 2014 г.). «Отсутствие активности супероксиддисмутазы вызывает фрагментацию ядерной ДНК в процессе старения». Связь с биохимическими и биофизическими исследованиями . 444 (2): 260–263. дои : 10.1016/j.bbrc.2014.01.056 . hdl : 11147/5542 . ПМИД 24462872 .
^ Альбертелла М., Джентьала Р.Р., Параскевас Т., Эрет Д., Брускеттини М., Солл Р. и др. (Кокрейновская группа по неонатологии) (октябрь 2023 г.). «Супероксиддисмутаза при бронхолегочной дисплазии у недоношенных детей». Кокрановская база данных систематических обзоров . 2023 (10): CD013232. дои : 10.1002/14651858.CD013232.pub2 . PMC 10561150. PMID 37811631 .
^ Сеги Дж., Жиронелла М., Санс М., Гранелл С., Гил Ф., Гимено М. и др. (сентябрь 2004 г.). «Супероксиддисмутаза облегчает колит, вызванный TNBS, за счет снижения окислительного стресса, экспрессии молекул адгезии и рекрутирования лейкоцитов в воспаленный кишечник». Журнал биологии лейкоцитов . 76 (3): 537–544. дои : 10.1189/jlb.0304196 . ПМИД 15197232 . S2CID 15028921 .
^ МакГиннесс Дж.Э., Проктор П.Х., Демопулос Х.Б., Хокансон Дж.А., Киркпатрик Д.С. (1978). «Уменьшение нефротоксичности цис-платины с помощью орготеина (супероксиддисмутазы)». Физиологическая химия и физика . 10 (3): 267–277. ПМИД 733940 .
^ Марбергер Х., Хубер В., Барч Г., Шульте Т., Свобода П. (1974). «Орготеин: новый противовоспалительный металлопротеиновый препарат, оценка клинической эффективности и безопасности при воспалительных состояниях мочевыводящих путей». Международная урология и нефрология . 6 (2): 61–74. дои : 10.1007/bf02081999 . ПМИД 4615073 . S2CID 23880216 .
^ Номер клинического исследования NCT01324141 для «Местного применения MTS-01 при дерматите во время лучевой и химиотерапии при анальном раке» на сайте ClinicalTrials.gov.
^ Возенин-Бротонс MC, Сиван В., Голт Н., Ренард С., Джеффротин С., Деланиан С. и др. (январь 2001 г.). «Антифибротическое действие Cu/Zn СОД опосредовано репрессией TGF-бета1 и фенотипической реверсией миофибробластов». Свободно-радикальная биология и медицина . 30 (1): 30–42. дои : 10.1016/S0891-5849(00)00431-7 . ПМИД 11134893 .
Arc.Ask3.Ru Номер скриншота №: 1ea5bcb1f8096721d34a01a0e02928d8__1721395440 URL1:https://arc.ask3.ru/arc/aa/1e/d8/1ea5bcb1f8096721d34a01a0e02928d8.html Заголовок, (Title) документа по адресу, URL1: Superoxide dismutase - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)