Рибонуклеотидредуктаза
| рибонуклеозиддифосфатредуктаза | |||
|---|---|---|---|
 Гетероктамер рибонуклеозиддифосфатредуктазы, E.Coli | |||
| Идентификаторы | |||
| Номер ЕС. | 1.17.4.1 | ||
| Номер CAS. | 9047-64-7sy | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
Рибонуклеотидредуктаза ( RNR ), также известная как рибонуклеозиддифосфатредуктаза ( rNDP ), представляет собой фермент , который катализирует образование дезоксирибонуклеотидов из рибонуклеотидов . [1] Он катализирует это образование, удаляя 2'-гидроксильную группу рибозного кольца нуклеозиддифосфатов. В результате этого восстановления образуются дезоксирибонуклеотиды. [2] Дезоксирибонуклеотиды, в свою очередь, используются в синтезе ДНК . Реакция, катализируемая РНР, строго консервативна у всех живых организмов. [3] Более того, RNR играет решающую роль в регулировании общей скорости синтеза ДНК, так что соотношение ДНК и клеточной массы поддерживается на постоянном уровне во время деления клеток и восстановления ДНК . [4] Несколько необычной особенностью фермента RNR является то, что он катализирует реакцию, протекающую по свободнорадикальному механизму действия. [5] [6] Субстратами для RNR являются ADP , GDP , CDP и UDP . dTDP (дезокситимидиндифосфат) синтезируется другим ферментом ( тимидилаткиназой ) из dTMP (дезокситимидинмонофосфат).
Структура [ править ]
Рибонуклеотидредуктазы делятся на три класса. Ферменты RNR класса I состоят из большой альфа-субъединицы и малых бета-субъединиц, которые объединяются с образованием активного гетеродимерного тетрамера . Восстанавливая NDP до 2'-dNDP, фермент катализирует синтез de novo дезоксирибонуклеотидов (dNTP), которые являются предшественниками синтеза ДНК и необходимы для пролиферации клеток . [7] РНР класса II продуцируют 5'-дезоксиаденозильный радикал путем гомолитического расщепления связи C-Co в аденозилкобаламине. Кроме того, RNR класса III содержат стабильный глициловый радикал. [8]
Люди являются носителями RNR класса I. Альфа-субъединица кодируется геном RRM1, тогда как существуют две изоформы бета-субъединицы, кодируемые генами RRM2 и RRM2B:
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Каждый альфа- мономер класса I состоит из трех доменов : [9]
- один преимущественно спиральный домен, содержащий 220 N-концевых остатков,
- вторая большая десятицепочечная α/β-структура, содержащая 480 остатков,
- и третья небольшая пятицепочечная α/β-структура, содержащая 70 остатков.
В Pfam второй домен интерпретируется как два отдельных домена:
- более короткий полностью альфа-N-концевой домен,
- и более длинный С-концевой домен.
|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


Бета-субъединица класса I обычно содержит диметаллический центр и стабильный тирозильный радикал . У людей бета-субъединица зависит от кофактора дижелеза. В E. coli тирозильный радикал расположен в положении 122 (Y122), обеспечивая стабильный радикал для субъединиц RNR2 класса I. [13] У A. aegypti этот тирозильный радикал расположен в положении 184 (Y184). [14] Тирозильный радикал глубоко похоронен внутри белка в гидрофобной среде, расположенной рядом с железным центром, который используется для стабилизации тирозильного радикала. В структуре двух ц-оксосвязанных желез преобладают лиганды, служащие центрами связывания железа: четыре карбоксилата [ аспартат (D146), глутамат (Е177, Е240 и Е274)] и два гистидина (Н180 и Н277). [14] Ассоциация происходит между C-концом RNR2 и C-концом RNR1. [9] Ферментативная активность зависит от ассоциации субъединиц RNR1 и RNR2. Активный центр состоит из активных дитиоловых групп из RNR1, а также дижелезного центра и тирозильного радикала из субъединицы RNR2.
Другие остатки RNR2, такие как аспартат (D273), триптофан (W48) и тирозин (Y356), дополнительно стабилизируют тирозильный радикал в активном центре, тем самым обеспечивая перенос электрона. [9] Эти остатки помогают в переносе радикального электрона от тирозина (Y122) RNR2 к цистеину (C439) RNR1. Перенос электрона начинается на тирозине RNR2 (Y122) и продолжается в RNR2 к триптофану (W48), который отделен от тирозина RNR1 (Y731) на 2,5 нанометра . Перенос электронов от RNR2 к RNR1 происходит через тирозин (Y356 к Y731) и продолжается через тирозин (Y730) к цистеину (C439) в активном центре. [15] Сайт-направленные мутации первичной структуры RNR указывают на то, что все указанные выше остатки участвуют в переносе свободного радикала на большие расстояния в активный центр. [9]
У комаров A. aegypti RNR1 сохраняет большинство важнейших аминокислотных остатков, включая аспартат (D64) и валин (V292 или V284), которые необходимы для аллостерической регуляции ; остатки пролина (P210 и P610), лейцина (L453 и L473) и метионина (M603), расположенные в гидрофобном активном центре; остатки цистеина (С225, С436 и С451), которые участвуют в удалении атома водорода и переносе радикального электрона в активный центр; остатки цистеина (C225 и C436), аспарагина (N434) и глутамата (E441), которые связывают рибонуклеотидный субстрат; остатки тирозина (Y723 и Y743), определяющие радикальный перенос; и остатки цистеина (C838 и C841), которые используются для регенерации дитиоловых групп в активном центре. [14]
Функция [ править ]
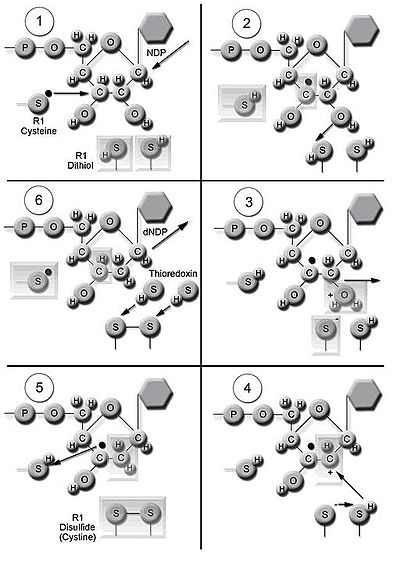
Фермент рибонуклеотидредуктаза (RNR) катализирует синтез dNDP de novo. [16] Катализ рибонуклеозид-5'-дифосфатов (NDP) включает восстановление 2'-углерода рибозо-5-фосфата с образованием 2'-дезоксирибонуклеозид-5'-дифосфатов (dNDP), восстановленных 2'-дезоксипроизводным. Это сокращение начинается с образования свободного радикала. После однократного восстановления RNR требует отдачи электронов от дитиоловых групп белка тиоредоксина . Регенерация тиоредоксина происходит, когда никотинамидадениндинуклеотидфосфат ( НАДФН ) обеспечивает два атома водорода, которые используются для восстановления дисульфидных групп тиоредоксина.
Три класса RNR имеют схожие механизмы восстановления NDP, но различаются доменом, генерирующим свободный радикал, конкретным металлом в структуре металлопротеина и донорами электронов. На всех занятиях используется химия свободных радикалов. [9] Редуктазы класса I используют железный центр с превращением двухвалентного железа в трехвалентное для генерации свободного тирозильного радикала. Восстановление субстратов НДП происходит в аэробных условиях. Редуктазы I класса делятся на IA и IB из-за различий в регуляции. Редуктазы класса IA распространены в эукариотах , эубактериях , бактериофагах и вирусах . Редуктазы класса IB обнаружены у эубактерий. Редуктазы класса IB также могут использовать радикал, образующийся при стабилизации биядерного марганцевого центра. Редуктазы класса II генерируют свободный радикал 5'-дезоксиаденозильный радикал из кобаламина (кофермент B12) и имеют более простую структуру, чем редуктазы класса I и класса III. Восстановление NDP или рибонуклеотид-5'-трифосфатов (NTP) происходит как в аэробных , так и в анаэробных условиях. Редуктазы II класса распространены в архебактериях , эубактериях и бактериофагах. Редуктазы класса III используют радикал глицина, образующийся с помощью S-аденозилметионина и железо-серного центра. Снижение NTP ограничивается анаэробными условиями. Редуктазы III класса распространены в архебактериях, эубактериях и бактериофагах. [9] [14] Организмы не ограничиваются наличием одного класса ферментов. Например, E. coli имеет RNR как класса I, так и класса III.
восстановления каталитического Механизм
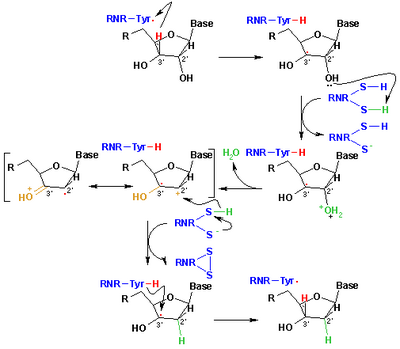
Принятый в настоящее время механизм восстановления рибонуклеотидов до дезоксирибонуклеотидов изображен на следующей схеме. На первом этапе происходит отщепление 3'-H субстрата 1 радикалом Cys439. Впоследствии реакция включает отщепление одной молекулы воды от углерода C-2' рибонуклеотида, катализируемое Cys225 и Glu441. На третьем этапе происходит перенос атома водорода от Cys225 на углерод C-2' 2'-кетильного радикала 3 после предыдущего переноса протона от Cys462 к Cys225. В конце этого этапа получаются радикальный анионный дисульфидный мостик и промежуточный кетон с закрытой оболочкой 4. Этот интермедиат был идентифицирован при превращении нескольких 2'-замещенных аналогов субстрата, а также с природным субстратом. [17] взаимодействие с мутантами ферментов. Следующим шагом является окисление анионного дисульфидного мостика с одновременным восстановлением субстрата с образованием 5. Спиновая плотность смещается от атомов серы к атому C-3' субстрата с одновременным переносом протона от Glu441 к углероду C. -3'. Последний этап является обратным первому и включает перенос водорода от Cys439 к C-3', регенерирующий исходный радикал и приводящий к конечному продукту 6.
Теоретические модели некоторых этапов этих механизмов с использованием полной модели белка R1 можно найти в исследованиях Cerqueira et al. . [18] [19]
Регламент [ править ]
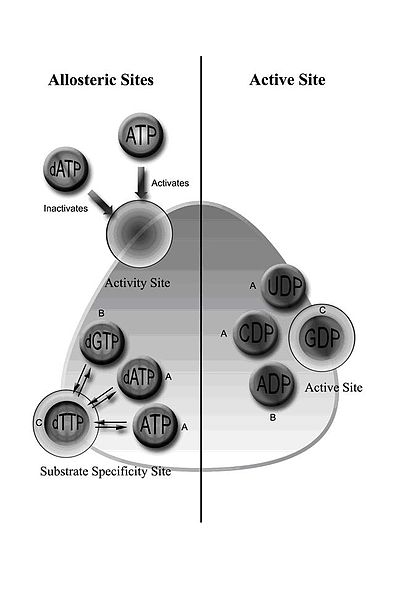
RNR класса I включает субъединицы RNR1 и RNR2, которые могут объединяться с образованием гетеродимерного тетрамера. [5] RNR1 содержит оба аллостерических сайта, опосредующих регуляцию специфичности и активности субстрата. [11] В зависимости от аллостерической конфигурации один из четырех рибонуклеотидов связывается с активным центром.
Регуляция RNR предназначена для поддержания сбалансированного количества dNTP. Связывание эффекторных молекул либо увеличивает, либо снижает активность RNR. Когда АТФ связывается с сайтом аллостерической активности, он активирует RNR. Напротив, когда dATP связывается с этим сайтом, он деактивирует RNR. [9] Помимо контроля активности, аллостерический механизм также регулирует специфичность субстрата и гарантирует, что фермент производит равное количество каждого dNTP для синтеза ДНК. [9] Во всех классах связывание АТФ или dATP с аллостерическим сайтом индуцирует восстановление цитидин-5'-дифосфата (CDP) и уридин-5'-дифосфата (UDP); 2'-дезоксигуанозин-5'-трифосфат (dGTP) индуцирует восстановление аденозин-5'-дифосфата (ADP); а 2'-дезокситимидин-5'-трифосфат (dTTP) индуцирует восстановление гуанозин-5'-дифосфата (GDP) (рис. 1).
Редуктазы класса IB не ингибируются dATP, поскольку им не хватает примерно 50 N-концевых аминокислот, необходимых для сайта аллостерической активности. [20] Кроме того, важно, чтобы активность рибонуклеотидредуктазы находилась под транскрипционным и посттранскрипционным контролем, поскольку синтез неповрежденной ДНК зависит от сбалансированного пула дезоксирибонуклеотидов. [21] Эукариотические клетки с редуктазами класса IA обладают механизмом негативного контроля, отключающим синтез dNTP по мере их накопления. Этот механизм защищает клетку от токсических и мутагенных эффектов, которые могут возникнуть в результате перепроизводства дНТФ, поскольку изменения в сбалансированных пулах дНТФ приводят к повреждению ДНК и гибели клеток. [22] [23] Хотя перепроизводство dNTP или несбалансированное их поступление может привести к неправильному включению нуклеотидов в ДНК, поступление dNTP может обеспечить восстановление ДНК. p53R2 представляет собой небольшую субъединицу рибонуклеотидредуктазы, которая может индуцировать такое восстановление. Изменения внутри этого гомолога R2, индуцированного p53, могут вызвать истощение митохондриальной ДНК и, следовательно, p53R2 служит основным фактором снабжения dNTP. [24]
RNR может использовать морфеиновую модель аллостерической регуляции . [25]
RNR1 Ингибиторы и RNR2
В целом ингибиторы RNR класса I можно разделить на три основные группы: ингибиторы трансляции, которые блокируют синтез фермента; ингибиторы димеризации, которые предотвращают объединение двух субъединиц RNR (R1 и R2); и каталитические ингибиторы, которые инактивируют субъединицу R1 и/или субъединицу R2. [18]
RNR класса I можно ингибировать пептидами , аналогичными C-концу RNR2. Эти пептиды могут конкурировать с RNR2 за связывание с RNR1, в результате чего RNR1 не образует ферментативно активного комплекса с RNR2. [26] [27] Хотя С-конец белков RNR2 у разных видов разный, RNR2 может взаимодействовать с RNR1 у разных видов. [28] Когда С-конец мышиного RNR2 был заменен С-концевыми аминокислотными остатками RNR2 E. coli (7 или 33), химерная субъединица RNR2 все еще связывается с субъединицами мышиного RNR1. Однако у них отсутствует ферментативная активность, вероятно, из-за элиминации остатков, участвующих в переносе свободнорадикального электрона от субъединицы RNR2 к субъединице RNR1. [27]
Маленькие пептиды могут специфически ингибировать связывание субъединиц RNR2 с RNR1, если они имеют значительное сходство с нормальным С-концом RNR2. [29] Это ингибирование связывания RNR2 с RNR1 было успешно протестировано на RNR вируса простого герпеса (HSV). Когда в конкурентных анализах использовался олигомер из 7 аминокислот (GAVVNDL), укороченный с C-конца субъединицы RNR2, это предотвращало образование нормального RNR2 ферментативно активного комплекса с RNR1. [30] Другие небольшие пептидные ингибиторы, подобные C-концу RNR2, также успешно использовались для ингибирования ферментативной активности RNR HSV и, следовательно, репликации HSV. [31] на мышах на моделях стромального кератита и неоваскуляризации роговицы ( , вызванное вирусом простого герпеса ) небольшой C-концевой аналог RNR2 BILD 1263 ингибирует RNR и эффективен в предотвращении этих заболеваний. окулярное заболевание Сообщалось, что [32] В некоторых случаях, хотя лечение небольшими аналогами С-конца не может остановить распространение заболевания, они все же могут помочь в выздоровлении. при устойчивом к ацикловиру ВПГ (PAAr5) небольшой пептидный ингибитор BILD 1633 в 5–10 раз более эффективен, чем BILD 1263, против кожной инфекции PAAr5. Сообщалось, что [33] Комбинированный подход к терапии (BILD 1633 и ацикловир) более эффективен для лечения местных поражений у мышей. Эти данные позволяют предположить, что небольшие пептидные ингибиторы, конкурирующие с RNR2 за связывание с RNR1, полезны для предотвращения распространения ВПГ.
Галлий ингибирует RNR2, заменяя Fe 3+ на активном сайте. Мальтолат галлия представляет собой биодоступную пероральную форму галлия, которая использует эту ингибирующую активность для лечения рака, инфекций и других заболеваний. [34]
Препараты гидроксимочевины [35] и мотексафин гадолиний препятствуют действию этого фермента. [36]
Ссылки [ править ]
- ^ Элледж С.Дж., Чжоу З., Аллен Дж.Б. (март 1992 г.). «Рибонуклеотидредуктаза: регуляция, регуляция, регуляция». Тенденции биохимических наук . 17 (3): 119–23. дои : 10.1016/0968-0004(92)90249-9 . ПМИД 1412696 .
- ^ Сниден Дж. Л., Леб Л. А. (сентябрь 2004 г.). «Мутации в субъединице R2 рибонуклеотидредуктазы, придающие устойчивость к гидроксимочевине» . Журнал биологической химии . 279 (39): 40723–8. дои : 10.1074/jbc.M402699200 . ПМИД 15262976 .
- ^ Торрентс Э., Элой П., Гиберт И., Родригес-Треллес Ф. (август 2002 г.). «Рибонуклеотидредуктазы: дивергентная эволюция древнего фермента» . Журнал молекулярной эволюции . 55 (2): 138–52. Бибкод : 2002JMolE..55..138T . дои : 10.1007/s00239-002-2311-7 . ПМИД 12107591 . S2CID 24603578 .
- ^ Херрик Дж., Склави Б. (январь 2007 г.). «Рибонуклеотидредуктаза и регуляция репликации ДНК: старая история и древнее наследие» . Молекулярная микробиология . 63 (1): 22–34. дои : 10.1111/j.1365-2958.2006.05493.x . ПМИД 17229208 . S2CID 9473163 .
- ^ Jump up to: Перейти обратно: а б Эклунд Х., Эрикссон М., Улин У., Нордлунд П., Логан Д. (август 1997 г.). «Рибонуклеотидредуктаза - структурные исследования радикального фермента». Биологическая химия . 378 (8): 821–5. дои : 10.1515/bchm.1997.378.8.815 . ПМИД 9377477 .
- ^ Стуббе Дж., Риггс-Джеласко П. (ноябрь 1998 г.). «Использование свободных радикалов: образование и функция тирозильного радикала в рибонуклеотидредуктазе». Тенденции биохимических наук . 23 (11): 438–43. дои : 10.1016/S0968-0004(98)01296-1 . ПМИД 9852763 .
- ^ Фэйрман Дж.В., Виджератна С.Р., Ахмад М.Ф., Сюй Х., Накано Р., Джа С., Прендергаст Дж., Велин Р.М., Флодин С., Роос А., Нордлунд П., Ли З., Уолц Т., Дилвис К.Г. (март 2011 г.). «Структурные основы аллостерической регуляции рибонуклеотидредуктазы человека путем олигомеризации, индуцированной нуклеотидами» . Структурная и молекулярная биология природы . 18 (3): 316–22. дои : 10.1038/nsmb.2007 . ПМК 3101628 . ПМИД 21336276 .
- ^ Ларссон К.М., Джордан А., Элиассон Р., Райхард П., Логан Д.Т., Нордлунд П. (ноябрь 2004 г.). «Структурный механизм регуляции специфичности аллостерического субстрата в рибонуклеотидредуктазе». Структурная и молекулярная биология природы . 11 (11): 1142–9. дои : 10.1038/nsmb838 . ПМИД 15475969 . S2CID 1025702 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час Джордан А., Райхард П. (1998). «Рибонуклеотидредуктазы» . Ежегодный обзор биохимии . 67 (1): 71–98. doi : 10.1146/annurev.biochem.67.1.71 . ПМИД 9759483 .
- ^ Jump up to: Перейти обратно: а б ПДБ : 1ПЭУ ; Уппстен М., Фернегорд М., Джордан А., Элиассон Р., Эклунд Х., Улин У. (июнь 2003 г.). «Структура большой субъединицы рибонуклеотидредуктазы класса Ib Salmonella typhimurium и ее комплексов с аллостерическими эффекторами». Журнал молекулярной биологии . 330 (1): 87–97. дои : 10.1016/S0022-2836(03)00538-2 . ПМИД 12818204 .
- ^ Jump up to: Перейти обратно: а б Улин У, Эклунд Х (август 1994 г.). «Структура белка рибонуклеотидредуктазы R1». Природа . 370 (6490): 533–9. Бибкод : 1994Natur.370..533U . дои : 10.1038/370533a0 . ПМИД 8052308 . S2CID 8940689 .
- ^ Нордлунд П., Эклунд Х. (июль 1993 г.). «Структура и функция белка рибонуклеотидредуктазы R2 Escherichia coli». Журнал молекулярной биологии . 232 (1): 123–64. дои : 10.1006/jmbi.1993.1374 . ПМИД 8331655 .
- ^ Хёгбом М., Андерссон М.Е., Нордлунд П. (март 2001 г.). «Кристаллические структуры окисленных биядерных центров марганца в Mn-замещенной рибонуклеотидредуктазе класса I из Escherichia coli: карбоксилатные сдвиги, имеющие значение для активации O2 и генерации радикалов». Журнал биологической неорганической химии . 6 (3): 315–23. дои : 10.1007/s007750000205 . ПМИД 11315567 . S2CID 20748553 .
- ^ Jump up to: Перейти обратно: а б с д Фам ДК, Блачута Б.Дж., Никол Х., Винзерлинг Дж.Дж. (сентябрь 2002 г.). «Субъединицы рибонуклеотидредуктазы комара желтой лихорадки Aedes aegypti: клонирование и экспрессия». Биохимия насекомых и молекулярная биология . 32 (9): 1037–44. дои : 10.1016/S0965-1748(02)00041-3 . ПМИД 12213240 .
- ^ Чанг MC, Йи К.С., Стуббе Дж., Носера Д.Г. (май 2004 г.). «Включение рибонуклеотидредуктазы путем образования световых радикалов аминокислот» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (18): 6882–7. Бибкод : 2004PNAS..101.6882C . дои : 10.1073/pnas.0401718101 . ПМК 406436 . ПМИД 15123822 .
- ^ Кокс М., Нельсон Д.Р. (2008). Ленингерские принципы биохимии . Сан-Франциско: WH Freeman. ISBN 978-0-7167-7108-1 .
- ^ Серкейра Н.М., Фернандес П.А., Эрикссон Л.А., Рамос М.Дж. (декабрь 2004 г.). «Активация рибонуклеотидов ферментом рибонуклеотидредуктазой: понимание роли фермента». Журнал вычислительной химии . 25 (16): 2031–7. дои : 10.1002/jcc.20127 . ПМИД 15481089 . S2CID 19665974 .
- ^ Jump up to: Перейти обратно: а б Серкейра Н.М., Перейра С., Фернандес П.А., Рамос М.Дж. (2005). «Обзор ингибиторов рибонуклеотидредуктазы: привлекательная мишень противоопухолевой терапии». Современная медицинская химия . 12 (11): 1283–94. дои : 10.2174/0929867054020981 . ПМИД 15974997 .
- ^ Серкейра Н.М., Фернандес П.А., Эрикссон Л.А., Рамос М.Дж. (март 2006 г.). «Дегидратация рибонуклеотидов, катализируемая рибонуклеотидредуктазой: роль фермента» . Биофизический журнал . 90 (6): 2109–19. Бибкод : 2006BpJ....90.2109C . дои : 10.1529/biophysj.104.054627 . ПМЦ 1386789 . ПМИД 16361339 .
- ^ Элиассон Р., Понтис Э., Джордан А., Райхард П. (октябрь 1996 г.). «Аллостерическая регуляция третьей рибонуклеотидредуктазы (фермента NrdEF) энтеробактерий» . Журнал биологической химии . 271 (43): 26582–7. дои : 10.1074/jbc.271.43.26582 . ПМИД 8900130 .
- ^ Теландер Л. (июнь 2007 г.). «Рибонуклеотидредуктаза и синтез митохондриальной ДНК». Природная генетика . 39 (6): 703–4. дои : 10.1038/ng0607-703 . ПМИД 17534360 . S2CID 22565931 .
- ^ Кунц Б.А. (1988). «Мутагенез и дисбаланс пула дезоксирибонуклеотидов». Мутационные исследования . 200 (1–2): 133–47. дои : 10.1016/0027-5107(88)90076-0 . ПМИД 3292903 .
- ^ Мейт М. (апрель 1989 г.). «Молекулярная основа мутаций, вызванных дисбалансом пула дезоксирибонуклеозидтрифосфата в клетках млекопитающих». Экспериментальные исследования клеток . 181 (2): 305–16. дои : 10.1016/0014-4827(89)90090-6 . ПМИД 2647496 .
- ^ Бурдон А., Минай Л., Серр В., Жайс Дж.П., Сарзи Э., Обер С., Кретьен Д., де Лонле П., Пакис-Флюклингер В., Аракава Х., Накамура Ю., Мюнних А., Рётиг А. (июнь 2007 г.). «Мутация RRM2B, кодирующего p53-контролируемую рибонуклеотидредуктазу (p53R2), вызывает серьезное истощение митохондриальной ДНК». Природная генетика . 39 (6): 776–80. дои : 10.1038/ng2040 . ПМИД 17486094 . S2CID 22103978 .
- ^ Селвуд Т., Яффе ЭК (март 2012 г.). «Динамическая диссоциация гомоолигомеров и контроль функции белка» . Архив биохимии и биофизики . 519 (2): 131–43. дои : 10.1016/j.abb.2011.11.020 . ПМЦ 3298769 . ПМИД 22182754 .
- ^ Климент I, Сьёберг Б.М., Хуан С.И. (май 1991 г.). «Карбокси-концевые пептиды как зонды для взаимодействия субъединиц рибонуклеотидредуктазы Escherichia coli: кинетический анализ исследований ингибирования». Биохимия . 30 (21): 5164–71. дои : 10.1021/bi00235a008 . ПМИД 2036382 .
- ^ Jump up to: Перейти обратно: а б Хаманн К.С., Лентендж С., Ли Л.С., Салем Дж.С., Ян Ф.Д., Куперман Б.С. (март 1998 г.). «Химерные ингибиторы малых субъединиц рибонуклеотидредуктазы млекопитающих: двойная функция С-конца R2?» . Белковая инженерия . 11 (3): 219–24. дои : 10.1093/белок/11.3.219 . ПМИД 9613846 .
- ^ Косентино Дж., Лавалле П., Рахит С., Планте Р., Годетт Ю., Лавец С., Уайтхед П.В., Дюсеппе Дж.С., Лепин-Френетт С., Дансеро Н. (январь 1991 г.). «Специфическое ингибирование рибонуклеотидредуктаз пептидами, соответствующими С-концу их второй субъединицы». Биохимия и клеточная биология . 69 (1): 79–83. дои : 10.1139/o91-011 . ПМИД 2043345 .
- ^ Куперман Б.С. (2003). «Олигопептидное ингибирование рибонуклеотидредуктаз класса I». Биополимеры . 71 (2): 117–31. дои : 10.1002/bip.10397 . ПМИД 12767114 . S2CID 25196379 .
- ^ Филатов Д., Ингемарсон Р., Греслунд А., Теландер Л. (август 1992 г.). «Роль карбоксильного конца рибонуклеотидредуктазы вируса простого герпеса малой субъединицы во взаимодействии субъединиц и формировании структуры железо-тирозильного центра» . Журнал биологической химии . 267 (22): 15816–22. дои : 10.1016/S0021-9258(19)49608-7 . ПМИД 1322407 .
- ^ Коэн Э.А., Годро П., Бразо П., Ланжелье Ю. (1986). «Специфическое ингибирование рибонуклеотидредуктазы герпесвируса нонапептидом, полученным из карбокси-конца субъединицы 2». Природа . 321 (6068): 441–3. Бибкод : 1986Natur.321..441C . дои : 10.1038/321441a0 . ПМИД 3012360 . S2CID 4238076 .
- ^ Брандт Ч.Р., Спенсер Б., Имеш П., Гарно М., Дезиэль Р. (май 1996 г.). «Оценка пептидомиметического ингибитора рибонуклеотидредуктазы на мышиной модели заболевания глаз вирусом простого герпеса 1 типа» . Антимикробные средства и химиотерапия . 40 (5): 1078–84. дои : 10.1128/aac.40.5.1078 . ПМК 163269 . ПМИД 8723444 .
- ^ Дуан Дж., Люцци М., Пэрис В., Ламберт М., Лавец С., Мосс Н., Харамилло Дж., Готье Дж., Дезиэль Р., Кордингли М.Г. (июль 1998 г.). «Противовирусная активность селективного ингибитора рибонуклеотидредуктазы в отношении устойчивого к ацикловиру вируса простого герпеса типа 1 in vivo» . Антимикробные средства и химиотерапия . 42 (7): 1629–35. дои : 10.1128/aac.42.7.1629 . ПМЦ 105657 . ПМИД 9660995 .
- ^ Бернштейн Л.Р. (декабрь 1998 г.). «Механизмы терапевтической активности галлия» (PDF) . Фармакологические обзоры . 50 (4): 665–82. ПМИД 9860806 .
- ^ «Сведения о ЕС 1.17.4.1 – рибонуклеозиддифосфатредуктаза» . Бренда . Проверено 25 июля 2015 г.
- ^ Хашеми С.И., Унгерстедт Дж.С., Захеди Авваль Ф., Холмгрен А. (апрель 2006 г.). «Мотексафин гадолиний, опухолеселективный препарат, воздействующий на тиоредоксинредуктазу и рибонуклеотидредуктазу» . Журнал биологической химии . 281 (16): 10691–7. дои : 10.1074/jbc.M511373200 . ПМИД 16481328 .
Внешние ссылки [ править ]
- Рибонуклеотиды + редуктазы Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
- База данных рибонуклеотидредуктазы (RNRdb). Архивировано 15 октября 2018 г. в Wayback Machine.