Изомераза
В биохимии изомеразы представляют собой общий класс ферментов , которые преобразуют молекулу из одного изомера в другой. при которых связи разрываются Изомеразы способствуют внутримолекулярным перегруппировкам , и образуются. Общий вид такой реакции следующий:
![{\displaystyle {\ce {AB}}\quad {\xrightarrow[{\text{изомераза }}]{}}\quad {\ce {BA}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/3e7114e4d3961ea355ca13869f9ea72fc1b38e08)
Существует только один субстрат, дающий один продукт. Этот продукт имеет ту же молекулярную формулу , что и субстрат, но отличается связностью связей или пространственным расположением. Изомеразы катализируют реакции во многих биологических процессах, например, гликолизе и углеводном обмене .
изомеризация
[ редактировать ]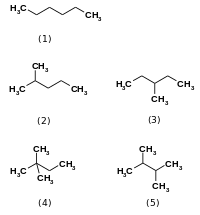

Изомеразы катализируют изменения внутри одной молекулы. [1] Они преобразуют один изомер в другой, а это означает, что конечный продукт имеет ту же молекулярную формулу, но другую физическую структуру. Сами изомеры существуют во многих разновидностях, но обычно их можно классифицировать как структурные изомеры или стереоизомеры . Структурные изомеры имеют разный порядок связей и/или разную связность связей друг от друга, как в случае гексана и четырех других его изомерных форм ( 2-метилпентан , 3-метилпентан , 2,2-диметилбутан и 2,3- диметилбутан ).
Стереоизомеры имеют одинаковый порядок отдельных связей и одинаковую связность, но трехмерное расположение связанных атомов различается. Например, 2-бутен существует в двух изомерных формах: цис -2-бутен и транс -2-бутен. [2] Подкатегории изомераз, содержащие рацемазы, эпимеразы и цис-транс-изомеры, являются примерами ферментов, катализирующих взаимное превращение стереоизомеров. Внутримолекулярные лиазы, оксидоредуктазы и трансферазы катализируют взаимные превращения структурных изомеров.
Распространенность каждого изомера в природе частично зависит от энергии изомеризации , разницы в энергии между изомерами. Близкие по энергии изомеры могут легко взаимопревращаться и часто встречаются в сопоставимых пропорциях. Энергия изомеризации, например, для превращения стабильного цис- изомера в менее стабильный транс -изомер больше, чем для обратной реакции, что объясняет, почему в отсутствие изомераз или внешнего источника энергии, такого как ультрафиолетовое излучение, данный цис -изомер имеет тенденцию присутствовать в больших количествах, чем транс -изомер. Изомеразы могут увеличивать скорость реакции за счет снижения энергии изомеризации. [3]
изомеразы Расчет кинетики на основе экспериментальных данных может быть более сложным, чем для других ферментов, поскольку использование экспериментов по ингибированию продуктов нецелесообразно. [4] То есть изомеризация не является необратимой реакцией , поскольку реакционный сосуд будет содержать один субстрат и один продукт, поэтому типичная упрощенная модель расчета кинетики реакции не работает. Существуют также практические трудности с определением определяющей скорость стадии при высоких концентрациях в одной изомеризации. Вместо этого изменение индикатора может преодолеть эти технические трудности, если существуют две формы несвязанного фермента. Этот метод использует изотопный обмен для косвенного измерения взаимного превращения свободного фермента между двумя его формами. Радиомеченный субстрат и продукт диффундируют в зависимости от времени. Когда система достигает равновесия, добавление немеченого субстрата нарушает или разбалансирует ее. Когда равновесие снова устанавливается, меченый радиоактивным изотопом субстрат и продукт отслеживаются для определения энергетической информации. [5]
Самое раннее использование этого метода прояснило кинетику и механизм действия фосфоглюкомутазы , отдав предпочтение модели непрямого переноса фосфата с одним промежуточным продуктом и прямого переноса глюкозы . [6] Затем этот метод был принят для изучения профиля пролинрацемазы и ее двух состояний: формы, которая изомеризует L- пролин , и другой формы, которая изомеризует D-пролин. При высоких концентрациях было показано, что состояние при этом взаимном превращении является лимитирующим и что эти формы фермента могут различаться только протонированием кислотной переходное и основной групп активного центра . [5]
Номенклатура
[ редактировать ]Обычно «названия изомераз образуются как « субстратная изомераза» (например, еноил-КоА-изомераза ) или как « субстратный тип изомеразы » (например, фосфоглюкомутаза )». [7]
Классификация
[ редактировать ]Каждая реакция, катализируемая ферментами, имеет уникальный классификационный номер. Реакции, катализируемые изомеразой, имеют свою категорию EC : EC 5. [8] Изомеразы подразделяются на шесть подклассов:
Рацемазы, эпимеразы
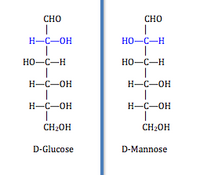
[ редактировать ]В эту категорию (КФ 5.1) входят ( рацемазы ) и эпимеразы ). Эти изомеразы инвертируют стереохимию целевого хирального углерода . Рацемазы действуют на молекулы с одним хиральным атомом углерода, инвертируя стереохимию, тогда как эпимеразы нацелены на молекулы с несколькими хиральными атомами углерода и действуют на один из них. Молекула только с одним хиральным углеродом имеет две энантиомерные формы, например, серин, имеющий изоформы D-серин и L-серин, отличающиеся только абсолютной конфигурацией хирального углерода. Молекула с несколькими хиральными атомами углерода имеет две формы каждого хирального углерода. Изомеризация по одному хиральному углероду нескольких дает эпимеры , отличающиеся друг от друга абсолютной конфигурацией только по одному хиральному углероду. [2] Например, D- глюкоза и D- манноза различаются по конфигурации всего по одному хиральному атому углерода. Этот класс далее подразделяется на группы, на которые действует фермент:
| Номер ЕС | Описание | Примеры |
|---|---|---|
| ЕС 5.1.1 | Воздействие на аминокислоты и производные | аланинрацемаза , метионинрацемаза |
| ЕС 5.1.2 | Воздействие на гидроксикислоты и их производные | лактатрацемаза , тартратэпимераза |
| ЕС 5.1.3 | Воздействие на углеводы и их производные | рибулозофосфат-3-эпимераза , УДФ-глюкозо-4-эпимераза |
| ЕС 5.1.99 | Воздействие на другие соединения | метилмалонил-КоА-эпимераза , гидантоинрацемаза |
Цис-транс-изомеразы
[ редактировать ]В эту категорию (EC 5.2) входят ферменты, катализирующие изомеризацию цис-транс-изомеров . Алкены и циклоалканы могут иметь цис-транс-стереоизомеры. Эти изомеры различаются не по абсолютной конфигурации, а скорее по положению групп заместителей относительно плоскости отсчета, например, поперек двойной связи или относительно кольцевой структуры. Цис -изомеры имеют группы заместителей на одной стороне, а транс- изомеры имеют группы на противоположных сторонах. [2]
Эта категория больше не разбивается. Все записи на данный момент включают:
Внутримолекулярные оксидоредуктазы
[ редактировать ]К этой категории (КФ 5.3) относятся внутримолекулярные оксидоредуктазы . Эти изомеразы катализируют перенос электронов из одной части молекулы в другую. Другими словами, они катализируют окисление одной части молекулы и одновременное восстановление другой части. [8] Подкатегории этого класса:
| Номер ЕС | Описание | Примеры |
|---|---|---|
| ЕС 5.3.1 | Взаимное преобразование альдоз и кетоз | Триозофосфатизомераза , Рибозо-5-фосфатизомераза |
| ЕС 5.3.2 | Взаимное преобразование кето- и енол-групп | Фенилпируваттаутомераза , оксалоацетаттаутомераза |
| ЕС 5.3.3 | Транспонирование двойных связей C=C | Стероидная дельта-изомераза , L-допахромизомераза |
| ЕС 5.3.4 | Транспонирование облигаций SS | Белковая дисульфид-изомераза |
| ЕС 5.3.99 | Другие внутримолекулярные оксидоредуктазы | Простагландин-D-синтаза , алленоксидциклаза |
Внутримолекулярные трансферазы
[ редактировать ]К этой категории (КФ 5.4) относятся внутримолекулярные трансферазы ( мутазы ). Эти изомеразы катализируют перенос функциональных групп из одной части молекулы в другую. [8] Фосфотрансферазы (КФ 5.4.2) до 1983 г. относились к категории трансфераз (КФ 2.7.5) с регенерацией доноров. [9] Этот подкласс можно разделить в зависимости от функциональной группы, которую переносит фермент:

| Номер ЕС | Описание | Примеры |
|---|---|---|
| ЕС 5.4.1 | Перенос ацильных групп | Лизолецитин ацилмутаза , метилмутаза прекоррина-8X |
| ЕС 5.4.2 | Фосфотрансферазы (фосфомутазы) | Фосфоглюкомутаза , Фосфопентомутаза |
| ЕС 5.4.3 | Перенос амино-групп | Бета-лизин 5,6-аминомутаза , тирозин 2,3-аминомутаза |
| ЕС 5.4.4 | Перенос гидроксигрупп | (гидроксиамино)бензолмутаза , изохоризматсинтаза |
| ЕС 5.4.99 | Перенос других групп | Метиласпартатмутаза , Хоризматмутаза |
Внутримолекулярные лиазы
[ редактировать ]К этой категории (EC 5.5) относятся внутримолекулярные лиазы . Эти ферменты катализируют «реакции, в которых группу можно считать удаленной из одной части молекулы, оставляя двойную связь, оставаясь при этом ковалентно связанной с молекулой». [8] Некоторые из этих катализируемых реакций включают разрыв кольцевой структуры.
Эта категория больше не разбивается. Все записи на данный момент включают:

Механизмы действия изомераз
[ редактировать ]Расширение и сжатие кольца за счет таутомеров
[ редактировать ]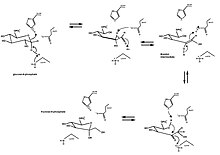
Классическим примером раскрытия и сжатия кольца является изомеризация глюкозы ( альдегида с шестичленным кольцом) во фруктозу ( кетон с пятичленным кольцом). Превращение D-глюкозо-6-фосфата в D-фруктозо-6-фосфат катализируется глюкозо-6-фосфатизомеразой , внутримолекулярной оксидоредуктазой . Общая реакция включает раскрытие кольца с образованием альдозы посредством кислотно-основного катализа и последующее образование промежуточного цис-эндиола. Затем образуется кетоза, и кольцо снова замыкается.
Глюкозо-6-фосфат сначала связывается с активным центром изомеразы. Изомераза раскрывает кольцо: ее His388 остаток протонирует кислород глюкозного кольца (и тем самым разрывает связь O5-C1) в сочетании с Lys518, кислород C1 депротонирующим гидроксильный . Кольцо раскрывается с образованием альдозы с прямой цепью и кислым протоном C2. Связь C3-C4 вращается, и Glu357 (с помощью His388) депронирует C2 с образованием двойной связи между C1 и C2. Образуется промежуточный цис -эндиол , и кислород C1 протонируется каталитическим остатком, что сопровождается депротонированием кислорода эндоола C2. с прямой цепью кетоза Образуется . Чтобы закрыть фруктозное кольцо, происходит обратное раскрытие кольца и кетоза протонируется. [10]
Эпимеризация
[ редактировать ]
Пример эпимеризации можно найти в цикле Кальвина, когда D-рибулозо-5-фосфат превращается в D-ксилулозо-5-фосфат под действием рибулозо-фосфат-3-эпимеразы . Субстрат и продукт различаются только стереохимией по третьему атому углерода в цепи. Основной механизм включает депротонирование третьего углерода с образованием реакционноспособного промежуточного енолята . Активный центр фермента содержит два остатка Asp . После того, как субстрат связывается с ферментом, первый Asp депротонирует третий углерод с одной стороны молекулы. Это оставляет плоскую sp 2 -гибридизированный промежуточный продукт. Второй Asp расположен на противоположной стороне активной стороны и протонирует молекулу, эффективно добавляя протон с обратной стороны. Эти совмещенные шаги инвертируют стереохимию третьего углерода. [11]
Внутримолекулярный перенос
[ редактировать ]
Хоризматмутаза представляет собой внутримолекулярную трансферазу и катализирует превращение хоризмата в префенат , используемый в качестве предшественника L-тирозина и L-фенилаланина в некоторых растениях и бактериях. Эта реакция представляет собой перегруппировку Кляйзена , которая может протекать с участием изомеразы или без нее, хотя скорость увеличивается на 10 6 сворачивание в присутствии хоризматмутазы. Реакция проходит через стула переходное состояние с субстратом в трансдиаксиальном положении. [12] Экспериментальные данные показывают, что изомераза избирательно связывает переходное состояние стула, хотя точный механизм катализа неизвестен. Считается, что это связывание стабилизирует переходное состояние посредством электростатических эффектов, что объясняет резкое увеличение скорости реакции в присутствии мутазы или при добавлении катиона, специально помещенного в активный центр. [13]
Внутримолекулярное оксидоредукция
[ редактировать ]
Изопентенилдифосфат-дельта-изомераза типа I (также известная как изомераза IPP) участвует в синтезе холестерина и, в частности, катализирует превращение изопентенилдифосфата (IPP) в диметилаллилдифосфат (DMAPP). В этой реакции изомеризации стабильная двойная связь углерод-углерод перестраивается с образованием высокоэлектрофильного аллильного изомера . Изомераза IPP катализирует эту реакцию путем стереоселективной антарафациальной транспозиции одного протона. Двойная связь протонируется при С4 с образованием третичного карбокатиона при С3. Соседний углерод C2 депротонируется с противоположной стороны, образуя двойную связь. [14] По сути, двойная связь сдвигается.
Роль изомеразы в заболеваниях человека
[ редактировать ]Изомераза играет роль в заболеваниях человека. Дефицит этого фермента может вызвать расстройства у человека.
Дефицит фосфогексозоизомеразы
[ редактировать ]Дефицит фосфогексозоизомеразы (PHI) также известен как дефицит фосфоглюкозоизомеразы или дефицит глюкозо-6-фосфатизомеразы и является наследственной недостаточностью фермента. PHI является второй наиболее часто встречающейся эртоэнзиопатией при гликолизе после дефицита пируваткиназы и связана с несфероцитарной гемолитической анемией различной степени тяжести. [15] [16] Это заболевание сосредоточено на белке глюкозо-6-фосфата. Этот белок можно найти в секрете некоторых раковых клеток. [17] PHI является результатом действия димерного фермента, который катализирует обратимое взаимное превращение фруктозо-6-фосфата и глюкозо-6-фосфата. [15]
PHI — очень редкое заболевание: на сегодняшний день в литературе описано всего 50 случаев. [15]
Диагноз ставят на основании клинической картины в сочетании с биохимическими исследованиями, выявляющими дефицит ГФИ эритроцитов (от 7 до 60% нормы) и выявлением мутации гена ГФИ с помощью молекулярного анализа. [15]
Дефицит фосфогексозоизомеразы может привести к состоянию, известному как гемолитический синдром . Как и у человека, исключительно у гомозиготных мутантов проявлялся гемолитический синдром, характеризующийся уменьшением числа эритроцитов, снижением гематокрита, снижением гемоглобина , увеличением числа ретикулоцитов и концентрацией билирубина в плазме, а также повышением печеночно- и селезёночно-соматических показателей. . [16]
Дефицит триозофосфат-изомеразы
[ редактировать ]Заболевание, называемое дефицитом триозофосфат-изомеразы (ТПИ), представляет собой тяжелое аутосомно-рецессивно-наследственное мультисистемное нарушение гликолитического метаболизма . [18] Оно характеризуется гемолитической анемией и нейродегенерацией и вызвано анаэробной метаболической дисфункцией. Эта дисфункция возникает в результате миссенс-мутации, которая влияет на кодируемый белок TPI. [19] Наиболее распространенной мутацией является замена гена Glu104Asp, которая приводит к наиболее тяжелому фенотипу и ответственна примерно за 80% клинического дефицита TPI. [18]
Дефицит TPI встречается очень редко: в литературе описано менее 50 случаев. [20] Будучи аутосомно-рецессивным наследственным заболеванием, дефицит TPI имеет 25% риск рецидива у гетерозиготных родителей. [18] [20] Это врожденное заболевание, которое чаще всего протекает при гемолитической анемии и проявляется желтухой. [18] Большинство пациентов с TPI по мутации Glu104Asp или гетерозиготными по нулевому аллелю TPI и Glu104Asp имеют ожидаемую продолжительность жизни от младенчества до раннего детства. У пациентов с TPI с другими мутациями обычно наблюдается более высокая продолжительность жизни. Есть только два случая, когда люди с ТПИ жили старше 6 лет. В этих случаях участвуют два брата из Венгрии, у одного из которых неврологические симптомы не развивались до 12 лет, и у старшего брата, у которого нет неврологических симптомов и который страдает анемией. только. [21]
У людей с TPI очевидные симптомы появляются после 6–24 месяцев. Эти симптомы включают: дистонию, тремор, дискинезию, симптомы пирамидного тракта, кардиомиопатию и поражение мотонейронов спинного мозга. [18] У пациентов также наблюдаются частые бактериальные инфекции дыхательной системы. [18]
TPI обнаруживается через дефицит ферментативной активности и накопление дигидроксиацетонфосфата (DHAP), который является токсичным субстратом, в эритроцитах. [18] [20] Это можно обнаружить посредством физического осмотра и ряда лабораторных исследований. При обнаружении обычно наблюдаются миопатические изменения в мышцах и хроническая аксональная нейропатия в нервах. [18] Диагноз ТПИ можно подтвердить с помощью молекулярной генетики. [18] Анализ ДНК ворсин хориона или анализ эритроцитов плода можно использовать для обнаружения TPI при антенатальной диагностике. [18]
Лечение ТПИ не является специфическим и варьируется в зависимости от случая. Из-за разнообразия симптомов, вызываемых TPI, для лечения одного человека может потребоваться группа специалистов. Эта команда специалистов будет состоять из педиатров, кардиологов, неврологов и других специалистов здравоохранения, которые смогут разработать комплексный план действий. [22]
Для лечения ТПИ также можно принять поддерживающие меры, такие как переливание эритроцитов в случаях тяжелой анемии. В некоторых случаях селезенкаудаление (спленэктомия) может улучшить течение анемии. Не существует лечения, предотвращающего прогрессированиеневрологические нарушения при любых других негематологических клинических проявлениях заболеваний. [23]
Промышленное применение
[ редактировать ]Безусловно, наиболее распространенное использование изомераз в промышленности - это производство сахара . Глюкозоизомераза (также известная как ксилозоизомераза ) катализирует превращение D- ксилозы и D- глюкозы в D- ксилулозу и D- фруктозу . Как и большинство сахароизомераз, глюкозоизомераза катализирует взаимное превращение альдоз и кетоз . [24]
Преобразование глюкозы во фруктозу является ключевым компонентом производства кукурузного сиропа с высоким содержанием фруктозы . Изомеризация более специфична, чем старые химические методы производства фруктозы, что приводит к более высокому выходу фруктозы и отсутствию побочных продуктов . [24] Фруктоза, полученная в результате этой реакции изомеризации, является более чистой и не имеет остаточного привкуса примесей . Кукурузный сироп с высоким содержанием фруктозы предпочитают многие производители кондитерских изделий и газированных напитков из-за высокой подслащивающей способности фруктозы (в два раза выше, чем у сахарозы). [25] ), его относительно низкая стоимость и неспособность кристаллизоваться. Фруктоза также используется в качестве подсластителя для диабетиков . [24] Основные проблемы использования глюкозоизомеразы связаны с ее инактивацией при более высоких температурах и требованием высокого pH (между 7,0 и 9,0) в реакционной среде. Умеренно высокие температуры, выше 70 °C, увеличивают выход фруктозы как минимум вдвое на стадии изомеризации. [26] Для фермента требуется двухвалентный катион, такой как Co. 2+ и мг 2+ в период пиковой активности это дополнительные затраты для производителей. Глюкозоизомераза также имеет гораздо более высокое сродство к ксилозе, чем к глюкозе, что требует тщательно контролируемой среды. [24]
Изомеризация ксилозы в ксилулозу имеет свои собственные коммерческие применения, поскольку интерес к биотопливу возрос. Эта реакция часто наблюдается в природе у бактерий , которые питаются разлагающимися растительными веществами. промышленное применение - производство этанола , достигаемое путем ферментации ксилулозы Его наиболее распространенное . Использование гемицеллюлозы в качестве исходного материала очень распространено. Гемицеллюлоза содержит ксилан , который сам состоит из ксилозы в β(1,4)-связях . [27] Использование глюкозоизомеразы очень эффективно превращает ксилозу в ксилулозу, на которую затем можно воздействовать путем ферментации дрожжей . В целом, обширные исследования в области генной инженерии были вложены в оптимизацию глюкозо-изомеразы и содействие ее извлечению из промышленных приложений для повторного использования.
Глюкозоизомераза способна катализировать изомеризацию ряда других сахаров, включая D- рибозу , D- аллозу и L- арабинозу . Наиболее эффективными являются субстраты, подобные глюкозе и ксилозе, имеющие экваториальные гидроксильные группы при третьем и четвертом атомах углерода. [28] Текущая модель механизма глюкозо-изомеразы представляет собой модель гидридного сдвига, основанную на рентгеновской кристаллографии и исследованиях изотопного обмена. [24]
Мембранассоциированные изомеразы
[ редактировать ]Некоторые изомеразы связываются с биологическими мембранами в виде периферических мембранных белков или закрепляются через одну трансмембранную спираль . [29] например, изомеразы с тиоредоксиновым доменом и некоторые пролилизомеразы .
Ссылки
[ редактировать ]- ^ Номенклатура ферментов, рекомендации Номенклатурного комитета Международного биохимического союза 1978 года по номенклатуре и классификации ферментов . Нью-Йорк: Академическая пресса. 1979. ISBN 9780323144605 .
- ^ Перейти обратно: а б с Макнот А.Д. (1997). Сборник химической терминологии (2-е изд.). Оксфорд: Научные публикации Блэквелла. ISBN 978-0-9678550-9-7 .
- ^ Уайтселл Дж.К., Фокс Массачусетс (2004). Органическая химия (3-е изд.). Садбери, Массачусетс: Джонс и Бартлетт. стр. 220–222. ISBN 978-0-7637-2197-8 .
- ^ Корниш-Боуден А (22 февраля 2013 г.). Основы кинетики ферментов (4-е изд.). Вайнхайм: Wiley-VCH. стр. 238–241. ISBN 978-3-527-66548-8 .
- ^ Перейти обратно: а б Фишер Л.М., Олбери В.Дж., Ноулз-младший (май 1986 г.). «Энергетика пролинрацемазы: эксперименты по возмущению индикаторов с использованием [14C] пролина, которые измеряют скорость взаимного превращения двух форм свободного фермента». Биохимия . 25 (9): 2538–42. дои : 10.1021/bi00357a038 . ПМИД 3521737 .
- ^ Бриттон Х.Г., Кларк Дж.Б. (ноябрь 1968 г.). «Механизм фосфоглюкомутазной реакции. Исследования фосфоглюкомутазы мышц кролика флюсовыми методами» . Биохимический журнал . 110 (2): 161–80. дои : 10.1042/bj1100161 . ПМК 1187194 . ПМИД 5726186 .
- ^ Брюс П.Я. (2010). Основная органическая химия (2-е изд.). Река Аппер-Седл, Нью-Джерси: Прентис-Холл. ISBN 978-0-321-59695-6 .
- ^ Перейти обратно: а б с д Уэбб ЕС (1992). Номенклатура ферментов 1992 г.: рекомендации Номенклатурного комитета Международного союза биохимии и молекулярной биологии по номенклатуре и классификации ферментов (6-е изд.). Сан-Диего: опубликовано для Международного союза биохимии и молекулярной биологии издательством Academic Press. ISBN 978-0-12-227164-9 .
- ^ Список ферментов класса 5 — изомеразы (PDF) . Номенклатурный комитет Международного союза биохимии и молекулярной биологии (NC-IUBMB). 2010.
- ^ Соломонс Дж.Т., Циммерли Э.М., Бернс С., Кришнамурти Н., Свон М.К., Крингс С., Мюрхед Х., Чиргвин Дж., Дэвис К. (сентябрь 2004 г.). «Кристаллическая структура мышиной фосфоглюкозоизомеразы с разрешением 1,6 А и ее комплекса с глюкозо-6-фосфатом раскрывает каталитический механизм раскрытия сахарного кольца». Журнал молекулярной биологии . 342 (3): 847–60. дои : 10.1016/j.jmb.2004.07.085 . ПМИД 15342241 .
- ^ Терада Т., Мукаэ Х., Охаси К., Хосоми С., Мидзогучи Т., Уэхара К. (апрель 1985 г.). «Характеристика фермента, который катализирует изомеризацию и эпимеризацию D-эритрозо-4-фосфата». Европейский журнал биохимии . 148 (2): 345–51. дои : 10.1111/j.1432-1033.1985.tb08845.x . ПМИД 3987693 .
- ^ Багг Т. (2012). «Глава 10: Изомеразы». Введение в химию ферментов и коферментов (3-е изд.). Уайли. ISBN 978-1-118-34896-3 .
- ^ Каст П., Грисостоми С., Чен И.А., Ли С., Кренгель У., Сюэ Ю., Хилверт Д. (ноябрь 2000 г.). «Стратегически расположенный катион имеет решающее значение для эффективного катализа хоризматмутазой» . Журнал биологической химии . 275 (47): 36832–8. дои : 10.1074/jbc.M006351200 . ПМИД 10960481 .
- ^ Чжэн В., Сунь Ф., Бартлам М., Ли Х, Ли Р., Рао З. (март 2007 г.). «Кристаллическая структура изопентенилдифосфатизомеразы человека при разрешении 1,7 А раскрывает ее каталитический механизм биосинтеза изопреноидов». Журнал молекулярной биологии . 366 (5): 1447–58. дои : 10.1016/j.jmb.2006.12.055 . ПМИД 17250851 .
- ^ Перейти обратно: а б с д Куглер В., Лакомек М. (март 2000 г.). «Дефицит глюкозо-6-фосфат-изомеразы». Лучшие практики и исследования Байьера. Клиническая гематология . 13 (1): 89–101. дои : 10.1053/beha.1999.0059 . ПМИД 10916680 .
- ^ Перейти обратно: а б Меркл С., Преч В. (1993). «Дефицит глюкозо-6-фосфат-изомеразы, связанный с несфероцитарной гемолитической анемией у мышей: животная модель заболевания человека» (PDF) . Кровь . 81 (1): 206–13. дои : 10.1182/blood.V81.1.206.206 . ПМИД 8417789 .
- ^ Кроне В., Шнайдер Г., Шульц Д., Арнольд Х., Блюм К.Г. (1 января 1970 г.). «Обнаружение дефицита фосфогексозоизомеразы в культурах фибробластов человека». Гумангенетика . 10 (3): 224–30. дои : 10.1007/BF00295784 . ПМИД 5475507 . S2CID 34123426 .
- ^ Перейти обратно: а б с д и ж г час я дж Орос ПФ. «Дефицит триозофосфат-изомеразы» . Сирота . Проверено 14 ноября 2013 г.
- ^ Селото А.М., Фрэнк А.К., Сейгл Дж.Л., Палладино М.Дж. (ноябрь 2006 г.). «Дрозофиловая модель наследственной гликолитической энзимопатии с дефицитом триозофосфат-изомеразы у человека» . Генетика . 174 (3): 1237–46. doi : 10.1534/genetics.106.063206 . ПМК 1667072 . ПМИД 16980388 .
- ^ Перейти обратно: а б с Олах Дж., Орос Ф., Кесерю Г.М., Ковари З., Ковач Дж., Холлан С., Овади Дж. (апрель 2002 г.). «Дефицит триозофосфат-изомеразы: нейродегенеративное заболевание неправильного сворачивания» (PDF) . Труды Биохимического общества . 30 (2): 30–8. дои : 10.1042/bst0300030 . ПМИД 12023819 . Архивировано из оригинала (PDF) 3 декабря 2013 г. Проверено 27 ноября 2013 г.
- ^ Холлан С., Фуджи Х., Хироно А., Хироно К., Карро Х., Мива С., Харшаньи В., Гьоди Э., Инсельт-Ковач М. (ноябрь 1993 г.). «Наследственный дефицит триозофосфат-изомеразы (TPI): два тяжелобольных брата, один с неврологическими симптомами, а другой без неврологических симптомов». Генетика человека . 92 (5): 486–90. дои : 10.1007/bf00216456 . ПМИД 8244340 . S2CID 3110178 .
- ^ «Дефицит триозофосфат-изомеразы» . НОРД . Проверено 14 декабря 2013 г.
- ^ «Дефицит триозофосфат-изомеразы -TPI» (PDF) . Проверено 26 ноября 2013 г.
- ^ Перейти обратно: а б с д и Бхосале С.Х., Рао М.Б., Дешпанде В.В. (июнь 1996 г.). «Молекулярные и промышленные аспекты глюкозо-изомеразы» . Микробиологические обзоры . 60 (2): 280–300. дои : 10.1128/мр.60.2.280-300.1996 . ПМК 239444 . ПМИД 8801434 .
- ^ Бейкер С. (1976). «Сиропы из чистой фруктозы». Технологическая биохимия . 11 :20–25.
- ^ Антрим Р.Л., Колилла В., Шнайдер Б.Дж. (1979). «Производство глюкозоизомеразы сиропов с высоким содержанием фруктозы». Прикладная биохимия и биоинженерия . 2 : 97–155.
- ^ Ван П.Ю., Шопсис С., Шнайдер Х. (май 1980 г.). «Ферментация пентозы дрожжами». Связь с биохимическими и биофизическими исследованиями . 94 (1): 248–54. дои : 10.1016/s0006-291x(80)80213-0 . ПМИД 6446306 .
- ^ Чен В.П. (август – сентябрь 1980 г.). «Глюкозоизомераза». Технологическая биохимия . 15 : 36–41.
- ^ Суперсемейства однопроходных трансмембранных лиаз в базе данных Membranome.